THESE. En vue de l'obtention du. Délivré par l'université Toulouse III - Paul Sabatier Discipline ou spécialité : Cancérologie
|
|
|
- Monique Sénéchal
- il y a 8 ans
- Total affichages :
Transcription
1 THESE En vue de l'obtention du DOCTORAT DE L UNIVERSITÉ DE TOULOUSE Délivré par l'université Toulouse III - Paul Sabatier Discipline ou spécialité : Cancérologie Présentée et soutenue par Zeïneb KARAA Le 12 décembre 2008 Titre : MicroARN et Hypoxie : Etude du contrôle de l expression de HIF-1α et du VEGF Pr. Catherine MULLER JURY Président du jury Dr. Fatima MECHTA-GRIGOURIOU Dr. Nelson DUSETTI Pr. Hervé PRATS Rapporteur Rapporteur Directeur de thèse Ecole doctorale... : Ecole (préciser Doctorale la qualité Biologie de chacun - Santé des - Biotechnologies membres) de Toulouse Unité de recherche : U858- I2MR- Équipe 15 Directeur de Thèse : Hervé PRATS Rapporteurs : Dr. Fatima MECHTA-GRIGOURIOU et Dr. Nelson DUSETTI
 de Toulouse Unité de recherche : U858- I2MR- Équipe 15 Directeur de Thèse :")
2 2
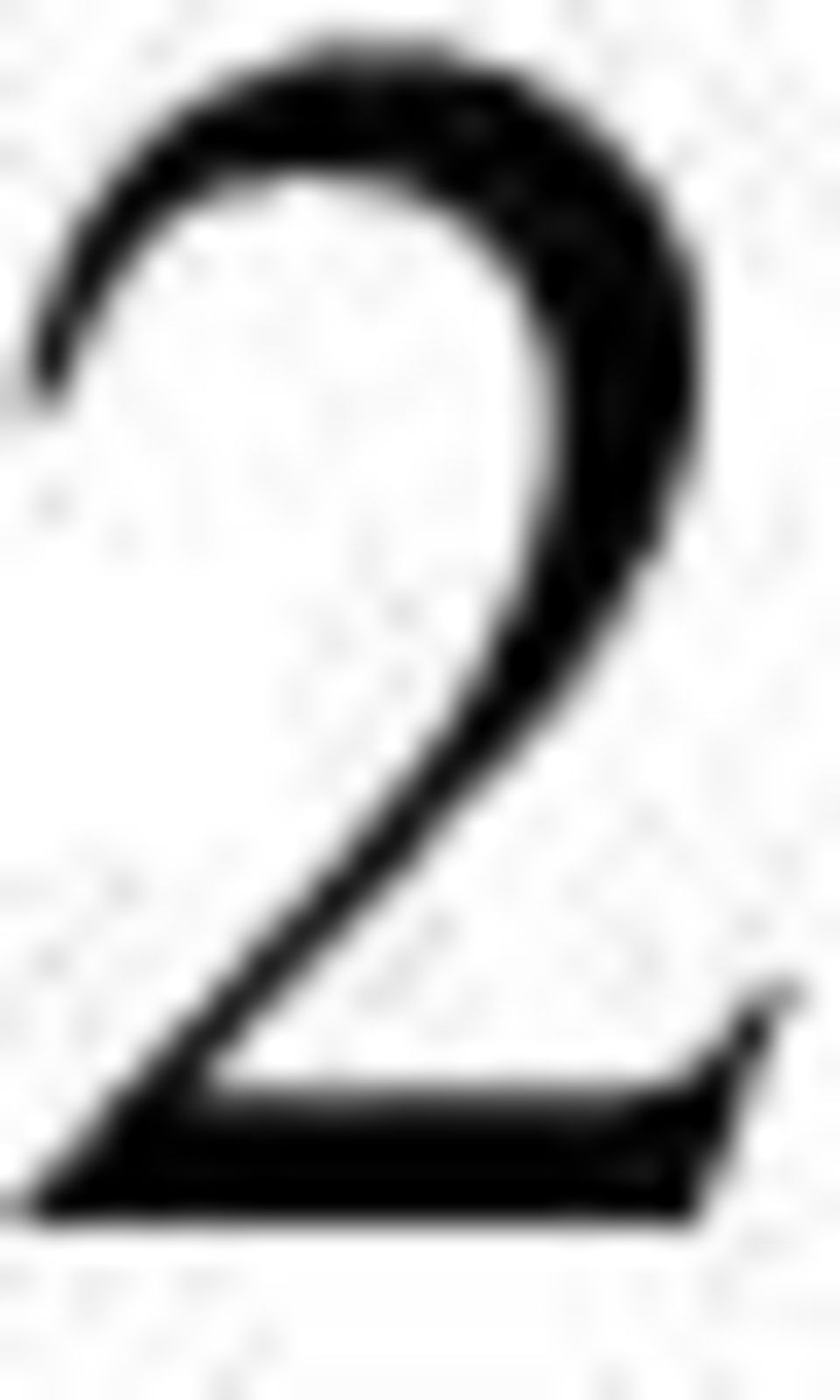
3 3
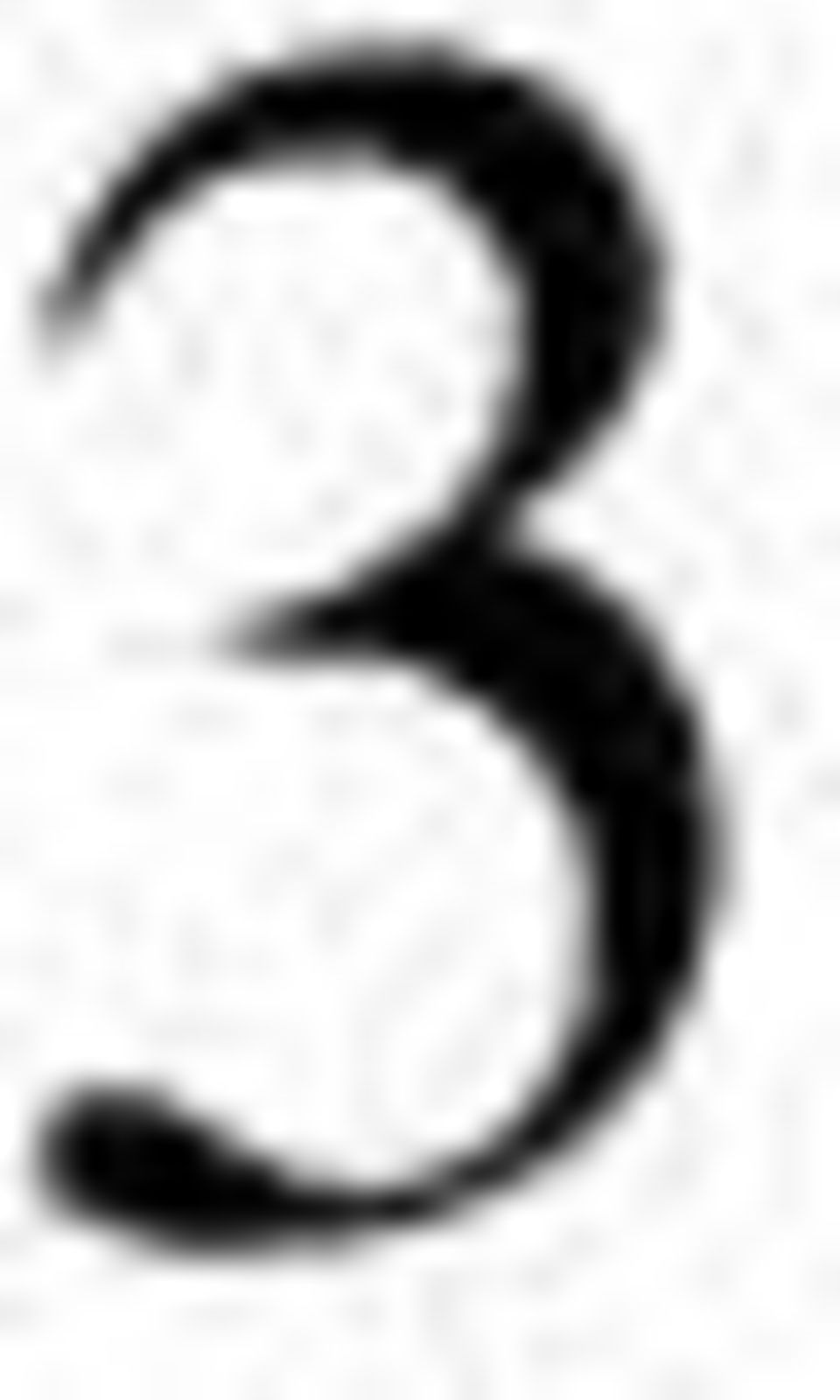
4 2
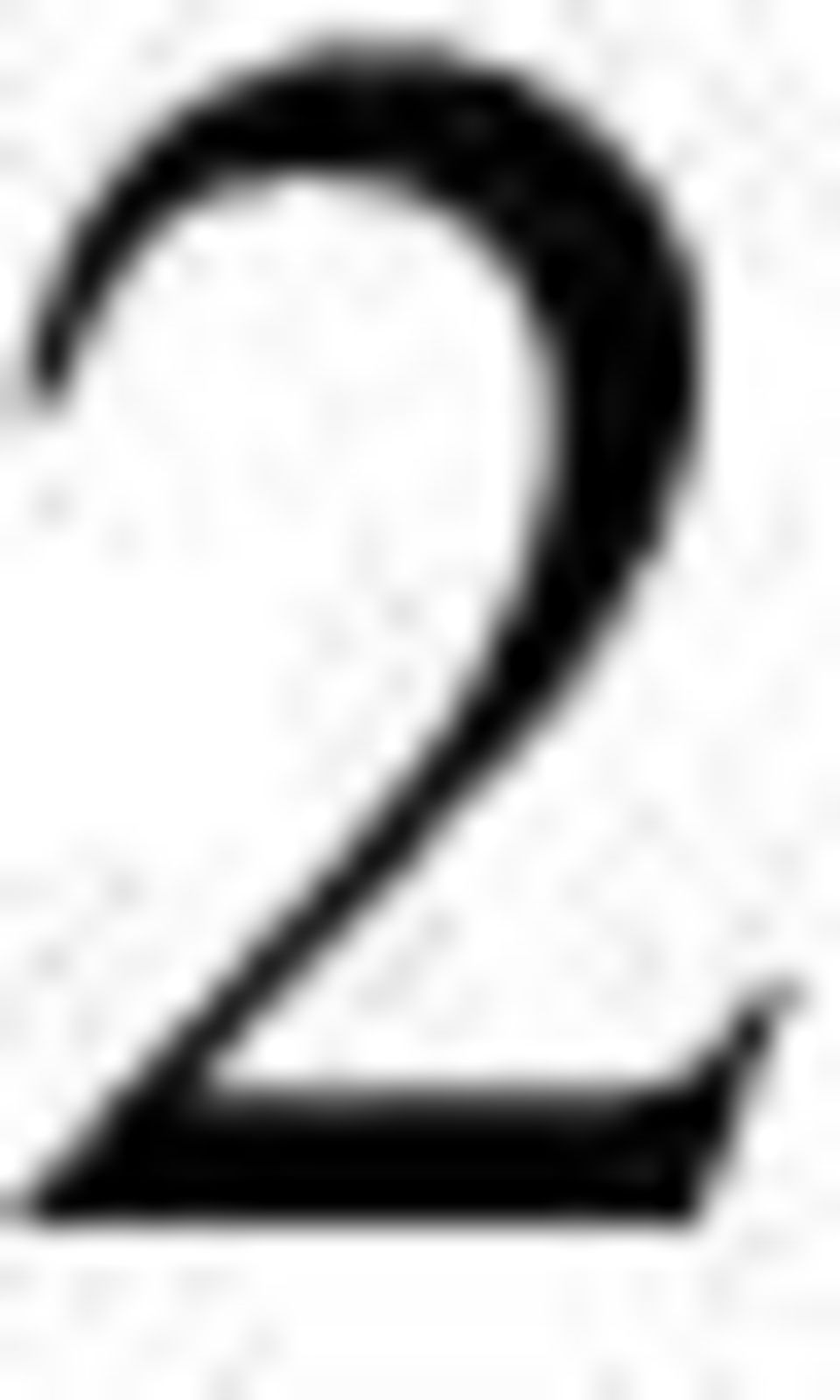
5 RÉSUMÉ Une diminution de la pression en oxygène (l hypoxie) constitue pour la cellule un changement environnemental qui induit des modifications importantes de l expression génique. Le facteur de transcription HIF-1, composé de deux sous-unités HIF-1α et HIF-1β, représente l acteur majeur dans la réponse à l hypoxie. Il est indispensable à la transduction de nombreux gènes en hypoxie, dont celle du VEGF. Le VEGF est un facteur angiogénique majeur et son inhibition est le principe de nouveaux traitements anticancéreux. L expression des gènes codant pour HIF-1α et VEGF est extrêmement régulée et, au cours de ma thèse, j ai essentiellement exploré le rôle des microarn dans ces régulations. Les microarn sont des petits ARN non codants qui jouent un rôle très important dans la régulation de l expression génique, principalement en bloquant la traduction des ARN messagers cibles. En conditions de stress, l initiation de la traduction du VEGF peut avoir lieu au niveau de deux sites d entrée interne des ribosomes, appelés IRES (IRES-A, et IRES-B) conduisant, dans le cas du choix de l IRES-B à la synthèse de la forme la plus diffusible du VEGF, et donc la plus active lors de l angiogenèse. Nous avons pu démontrer que mir-16 inhibe la traduction du VEGF en se fixant à un seul site présent dans la région 3 UTR du VEGF. De plus, cette étude démontre que mir-16 est capable d inhiber sélectivement la traduction initiée à l IRES- B du VEGF. Ainsi, et pour la première fois, nous montrons qu un microarn peut contrôler différents types d initiation de la traduction et ce, au sein d un même transcrit. Notre étude démontre ainsi que mir-16 régule négativement et préférentiellement l expression des formes les plus diffusibles de VEGF. Par ailleurs, nous avons étudié le rôle de mir-16 sur HIF-1α, et nous avons pu montrer que, dans ce cas, mir-16 régule positivement l accumulation de HIF-1α en s associant à la protéine HIF-1α et en contrôlant la stabilité de la protéine. Cet effet positif d un microarn sur une protéine passe donc par une voie particulièrement originale puisque, pour la première fois, nous montrons qu un microarn peut agir sur une autre cible qu un ARN messager. Enfin, dans un modèle de xénogreffe, nous avons pu montrer que l inhibition de mir-16 inhibe la croissance tumorale en bloquant l accumulation et donc l activité de HIF-1α dans les tumeurs. J ai donc pu montrer que mir-16 est un microarn fondamental dans la régulation de l angiogenèse puisqu il opère dans une boucle de rétrocontrôle où il régule positivement l expression de HIF-1α et négativement celle du VEGF. L ensemble de mon travail montre un nouvel aspect de la finesse et de la puissance du contrôle de l expression des gènes impliqués dans l angiogenèse tumorale. 1

6 2

7 SOMMAIRE RESUME...1 SOMMAIRE...3 INTRODUCTION...11 I ERE PARTIE : LES MECANISMES DE REGULATION DE L EXPRESSION GENIQUE..13 I- LES DIFFERENTES ETAPES DE LA REGULATION DE L EXPRESSION GENIQUE15 A- La transcription B- La maturation des ARN messagers L ajout de la coiffe L épissage Mécanisme général L épissage alternatif La maturation en C- Le contrôle de la stabilité des ARN messagers D- La traduction Mécanismes d initiation de la traduction Initiation à la coiffe Initiation par entrée interne des ribosomes (IRES) Régulation de la traduction Au niveau des facteurs d initiation de la traduction Par choix de codons alternatifs d initiation de la traduction Par la présence de petits cadres ouverts de lecture E- Régulation de l expression de l expression génique par les micro-arn Biogenèse des microarn Principes de l interaction miarn-arnm Mécanismes post-transcritionnels régulés par les microarn Régulation de l initiation de la traduction
... 26 2-Régulation de la traduction.")
8 3-1-1 Régulation de l initiation de la traduction initiée à la coiffe, par les miarn Régulation de la traduction indépendamment de la coiffe, par les miarn Régulation de la stabilité des messagers, la déadénylation P-bodies et les granules de stress Deux exemples de régulations positives de l expression génique par les microarn II EME PARTIE : L HYPOXIE...43 II- L HYPOXIE...45 A- HIF Généralités HIF HIF-1α HIF-1α, description Le KO de HIF-1α HIF-1β HIF-1β, description Le KO de HIF-1β HIF-2 et HIF HIF-2α et HIF-3α HIF-2β, HIF-3β et HIF4β B- La régulation de la stabilité de HIF-1α Régulation de la stabilité de HIF-1α, dépendante de l oxygène Description, mécanisme Les PHD Rôle des PHD dans le contrôle de HIF-1α VHL Description de VHL Contrôle de HIF-1α par VHL Le KO de VHL MDM Généralités, Description Contrôle de HIF-1α par MDM HDAC4, HDAC Régulation de la stabilité de HIF-1α en normoxie HSP Description de HSP

9 2-1-2 Contrôle de HIF-1α par HSP RACK Description de RACK Contrôle de HIF-1α par RACK C- La régulation de l activité transcriptionnelle de HIF Mécanisme, généralités CBP/p CBP/p300, généralités Rôle du couple CBP/p300 dans la régulation de l activité transcriptionnelle de HIF FIH HDAC D- Conséquences de l hypoxie Contrôle de la transcription en hypoxie HIF-1, régulateur central de l'expression de gènes en hypoxie HIF-1α versus HIF-2α Régulation de la traduction en hypoxie Régulation du mécanisme général de la traduction Micro-ARN et hypoxie Hypoxie et cancer III EME PARTIE : LE VEGF-A...69 III- LE VEGF-A...71 A- La famille des VEGF et leurs récepteurs La famille des VEGF Le VEGF-A Le VEGF-B Le VEGF-C Le VEGF-D Le VEGF-E Le PlGF Les récepteurs au VEGF Le VEGFR Le VEGFR Le VEGFR B- Rôles du VEGF et de ses isoformes dans l angiogenèse Rôles du VEGF dans l angiogenèse

10 1-1 Dans l angiogenèse physiologique Dans le développement embryonnaire Chez l adulte Dans l angiogenèse pathologique Rôles des isoformes du VEGF dans l angiogenèse Dans l angiogenèse physiologique Dans l angiogenèse pathologique C- La régulation de l expression génique du VEGF La transcription du VEGF Les facteurs de transcription du VEGF SP AP Conditions de régulation de la transcription du VEGF Les facteurs de croissance Les hormones Les oncogènes et les suppresseurs de tumeur L initiation alternative de la transcription du VEGF Maturation et stabilité des ARNm VEGF L épissage La maturation en Contrôle de la stabilité des ARNm VEGF La régulation de la traduction du VEGF Les IRES du VEGF Les codons alternatifs d initiation de la traduction du VEGF Le petit cadre ouvert de lecture du VEGF Les éléments en cis de la région 3 UTR RESULTATS...91 I- CONTROLE DE L EXPRESSION DU VEGF PAR MIR Rationnel Objectifs II- CONTROLE DE L EXPRESSION DE HIF-1α PAR MIR Rationnel Objectifs

11 Résultats III CONTROLE DE L EXPRESSION DE HIF-1α PAR MIR-17-5P Rationnel Objectifs Résultats Conclusions CONCLUSIONS, DISCUSSIONS ET PERSPECTIVES BIBLIOGRAPHIE Rationnel RESULTATS 7

12 8
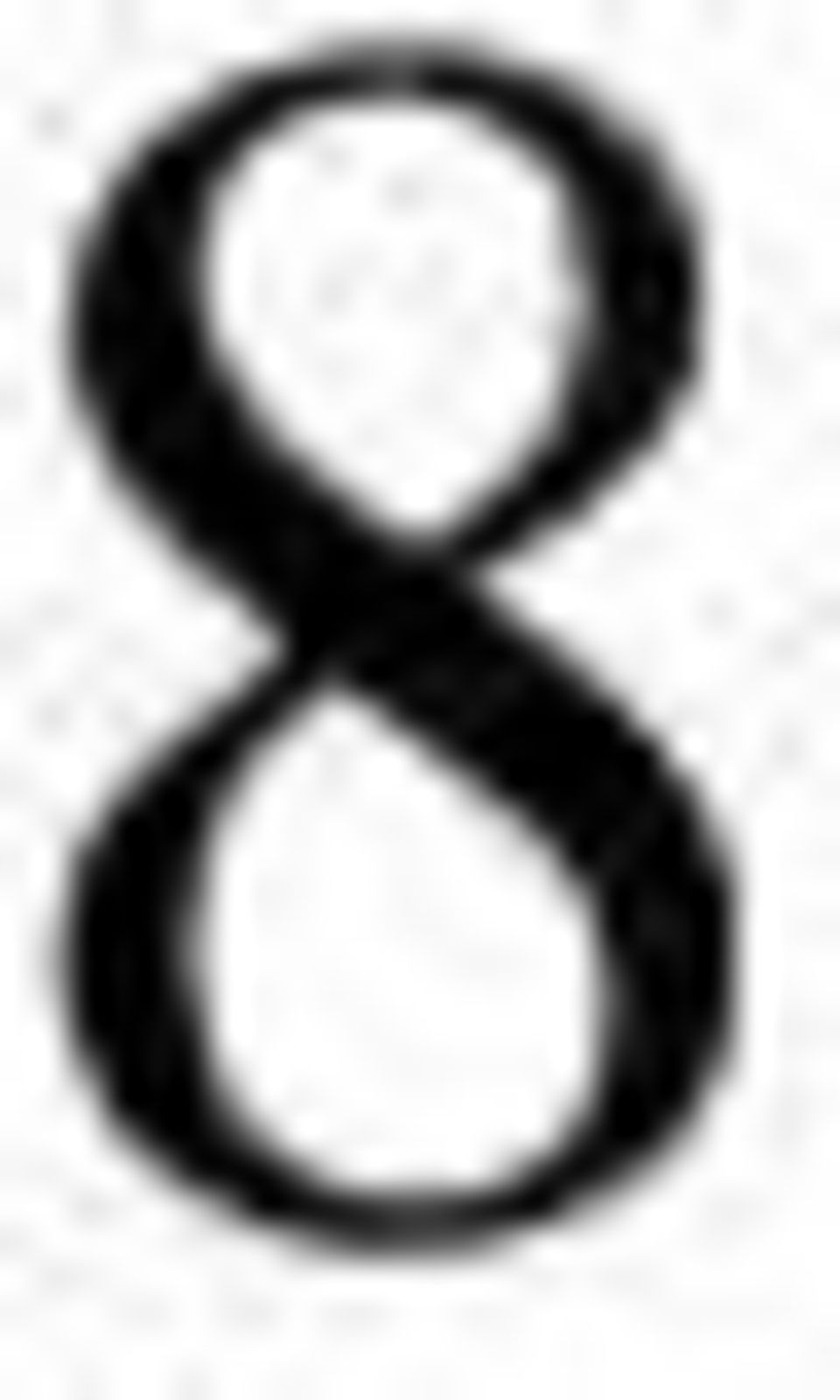
13 LISTE DES ABRÉVIATIONS 5 SS / 3 SS : 5 and 3 splice site (Site donneur et site accepteur d épissage) AA: acide aminé ADN : acide déoxyribonucléique AGO: argonaute ARE: AU rich element ARN Pol : ARN polymérase ARN: acide ribonucléique ARNm: ARN messager ARNr: ARN ribosomique ARNt: ARN de transfert ATP : adénosine triphosphate CTAD: C- terminal trans activation domain CTD : domaine carboxy-terminal eef: eukaryotic elongation factor EJC: exon-exon junction complex ELISA : Enzyme-linked immunosorbent assay (dosage d'immunosorption liée à enzyme) EMCV : Encephalomyocarditis virus ESE : exonic splicing enhancers ESS: exonic splice silencer GTP : guanosine triphosphate HeLa : cellules proviennant d'un prélèvement de métastase effectué sur une patiente atteinte d'un cancer du col de l'utérus et décédée en 1951, Henrietta Lacks. HEK 293: human Embryonic Kidney 293 cells (cellules embryonnaires humaines de rein) HIF-1: hypoxia inductible factor 1 HRE: hypoxia responsive element ISS : intronic splice silencer IRES : internal ribosome entry site. ITAF : IRES-trans-acting. factors kb : kilo base kda: kilo Dalton KI: knockin LNA : locked nucleic acid Met-ARNti : ARN de transfert- méthionine initiateur miarn : microarn mi-risc : mirna-induced silencing complexes NLS: nuclear localization signal nt: nucléotides NTAD: N- terminal trans activation domain ODD: O 2 dependant degradation PAS: Per-ARNt-Sim pb: paire de base PBS : Phosphate Buffered Saline PHD: proline-hydroxylase PIC : preinitiation complex pb: paire de bases PBS : Phosphate Buffered Saline PHD: proline-hydroxylase pri-pré-microarn : microarn précurseur primaire pré-microarn : microarn précurseur pré-microarn : microarn précurseur RPA: Rnase protection assay RISC : RNA-induced silencing complex 9

14 RPA: Rnase protection assay sirna: small interfering RNA snrnp: small nuclear ribonucleoprotein S: svedberg (unité de mesure du taux de sédimentation) SR: serine arginine rich RPA: Rnase protection assay SG : stress granules uaug : upstream AUG uorf: upstream ORF UTR : untranslated (non traduit) VEGF : vascular endothelial growth factor VHL: Von Hippel Lindau sirna: small interfering RNA snrnp: small nuclear ribonucleoprotein S: svedberg (unité de mesure du taux de sédimentation) SR: serine arginine rich SG : stress granules uaug : upstream AUG uorf: upstream ORF VEGF : vascular endothelial growth factor VHL: Von Hippel Lindau 10
 SR: serine arginine rich SG : stress granules uaug : upstream AUG uorf: upstream ORF VEGF : vascular endothelial growth factor VHL: Von Hippel")
15 INTRODUCTION 11

16 12
17 I ÈRE PARTIE : LES MÉCANISMES DE RÉGULATION DE L EXPRESSION GÉNIQUE 13

18 14

19 I- Les différentes étapes de la régulation de l expression génique A- La transcription La transcription est l étape de la régulation de l expression génique qui consiste à synthétiser de l ARN à partir d une matrice ADN. La régulation de la transcription est cruciale et déterminante sur le niveau d expression d un gène donné, et par conséquent, sur la croissance et la différenciation cellulaires. Chez les organismes eucaryotes, cette réaction est assurée par trois ARN polymérases : Pol I, Pol II et Pol III qui assurent respectivement la synthèse des ARNs ribosomiques, messagers et de transfert. Ces ARN polymérases sont assistées d autres facteurs en trans regroupés sous le terme général de «facteurs de transcription». Ces derniers, en réponse à un signal donné, sont impliqués dans la reconnaissance des éléments en cis contenus dans les séquences promotrices, et vont donner le signal pour activer la transcription d un gène cible. La transcription des ARN messagers (ARNm) commence par la liaison de facteurs de transcription spécifiques près du site d initiation de la transcription par l ARN polymérase II. Leur liaison initie le recrutement de l ensemble des éléments composant la machinerie d initiation de la transcription au niveau d un site qui sera alors défini comme étant le cœur du promoteur (core promoter). La définition de ce site sert au positionnement de l ARN polymérase II pour former le complexe de préinitiation (preinitiation complex, PIC). Le «cœur du promoteur» contient plusieurs éléments de séquence dont la boîte TATA (TBP-binding site), BRE (TFIIB-recognition element), Inr (initiator element) et DPE (downstream promoter element). La majorité des promoteurs contient au moins un de ces éléments en cis, aucun n est absolument requis. Enfin, une fois que l ARN polymérase II a synthétisé une trentaine de bases d ARN, elle se détache du cœur du promoteur et de la machinerie d initiation de la transcription et entre en phase d élongation (Hahn, 2004). 15

20 B- La maturation des ARN messagers Dans le noyau des cellules eucaryotes, la grande majorité des ARN pré-messagers (ARN pré-m) transcrits par l ARN polymérase II subissent une série de modifications cotranscriptionnelles : ajout de la coiffe à l'extrémité 5' non traduite (UTR), épissage des introns et maturation en 3. Le messager mature résultant est ensuite exporté vers le cytoplasme où il sera traduit. 1- L ajout de la coiffe La coiffe est constituée d un résidu guanosine méthylé en position N7, relié au premier nucléotide retrouvé en 5 de l ARNm via une liaison 5-5 triphosphate (Figure 1). L ajout de la coiffe est une modification co-transcriptionnelle qui se produit en trois étapes. Le phosphate-γ initialement présent à l extrémité 5 de l ARN pré-m est hydrolysé par l ARN triphosphatase (RTase). Ensuite, l ARN guanilytransférase (GTase) transforme une molécule de GTP en GMP pour la transférer à l extrémité diphosphorylée de l ARN pré-m. Il y a alors formation d un pont 5-5 triphosphate. Enfin, la méthylation du résidu guanosine en position N7 par l ARN (guanine-7) méthyltransférase (MTase) achève la mise en place de la coiffe. Cette étape est un processus directement couplé à la transcription puisque la GTase et la MTase sont toutes deux directement recrutées au niveau de l extrémité carboxyterminale l ARN polymérase II (lorsqu elle est phosphorylée). 16
.")
21 La coiffe exerce un rôle critique dans le métabolisme des ARNm à plusieurs niveaux. Sa formation est tout d abord indispensable à la poursuite des étapes ultérieures de maturation des ARN pré-m par l intermédiaire de la liaison du complexe CBP20/CBP80 (cap bing protein 20 et 80). Le recrutement de ce complexe va participer à la mise en place de la machinerie d épissage du premier intron et va influencer l efficacité de la maturation en 3 (Proudfoot et al., 2002). La coiffe est indispensable pour la stabilité des messagers puisqu elle fait office de barrière aux exonucléases qui agissent en 5 3. Enfin, la coiffe joue aussi un rôle essentiel à la traduction des transcrits matures une fois exportés dans le cytoplasme (CF Chaptre D1-1) (Proudfoot et al., 2002). 2- L épissage 2-1- Mécanisme général L épissage des introns constitue, chez les eucaryotes supérieurs, une étape clé dans le contrôle de l expression génique en déterminant la nature de la phase codante du transcrit mature (Black, 2003). L épissage d un ARN pré-m est dirigé par des éléments de séquences en cis indispensables au recrutement de la machinerie d épissage en trans (Figure 2). Le dinucléotide GU marque la jonction exon/intron en 5 et se situe au sein d une séquence de 6 à 9 nts qui constitue le site 5 d épissage (5 SS splice site). Le site 3 d épissage (3 SS splice site) comporte 3 éléments de séquence: le site de branchement (A) suivi d une séquence riche en pyrimidines (Py) et du dinucléotide terminal AG à l extrémité 3 de l intron (3 SS). L épissage des introns est un processus qui se produit en deux étapes de trans-estérification. Ainsi, la liaison 3-5 phosphodiester du résidu guanosine présent au niveau du 5 SS subit une attaque nucléophile par le résidu adénosine hautement conservé trouvé au site de branchement. Il y a alors formation d une interaction covalente entre ces deux nucléotides permettant à l intron d adopter la forme d un lasso. L extrémité 3 libre de l exon, en amont de l intron à épisser, réalise la deuxième trans-estérification par l attaque nucléophile de la liaison 3-5 phosphodiester du résidu guanosine 3 SS. Il y a alors détachement de l intron déjà sous forme de lasso, et liaison entre les deux exons adjacents. Les deux étapes de trans-estérification sont guidées par la fixation sur ces éléments en cis d un grand complexe catalytique macromoléculaire, le "spliceosome". Parmi les facteurs impliqués, on dénombre des snrnp de type U (U1, U2, U4, U5 et U6) et près d une centaine de protéines. La formation du spliceosome débute par la formation du complexe E (E: Early) où il y a appariement de U1 au 5 SS, de SF1 au résidu (A), de U2AF-65 à l élément Py 17
22 et de U2AF-35 au 3 SS. Par la suite, le pré-spliceosome est formé par la délocalisation de SF1 et la liaison de U2 au point d embranchement. Le complexe A apparaît grâce à la réorganisation du préspliceosome qui implique la formation d interactions entre les facteurs U1, U2 et U2AF. Cela contribue au rapprochement des exons favorisant leur éventuelle liaison. L arrivée du tri-snrnp U4/U5/U6 au sein des séquences consensus d épissage entraîne la formation du quatrième complexe, le complexe B. Sa présence permet de libérer le point d embranchement de même que le 3 SS. Finalement, le complexe catalytique, ou complexe C, est formé par l association de U6 et U2 au 5 SS, déclenchant le départ de U1 et U4. Les étapes de trans-estérification peuvent alors avoir lieu (Black, 2003). Des complexes multiprotéiques, appelés EJC (exon-exon junction complex), sont déposés après épissage au niveau des liaisons exon-exon. Ces complexes, jouent un rôle très important dans la maturation, l export nucléaire, la localisation cytoplasmique, la stabilité ainsi que la traductibilité des messagers (Le Hir and Seraphin, 2008). et de U2AF-35 au 3 SS. Par la suite, le pré-spliceosome est formé par la délocalisation de SF1 et la liaison de U2 au point d embranchement. Le complexe A apparaît grâce à la réorganisation du pré-spliceosome qui implique la formation d interactions entre les facteurs U1, U2 et U2AF. Cela contribue au rapprochement des exons favorisant leur éventuelle liaison. L arrivée du tri-snrnp U4/U5/U6 au sein des séquences consensus d épissage entraîne la formation du quatrième complexe, le complexe B. Sa présence permet de libérer le point d embranchement de même que le 3 SS. Finalement, le complexe catalytique, ou complexe C, est formé par l association de U6 et U2 au 5 SS, déclenchant le départ de U1 et U4. Les étapes de trans-estérification peuvent alors avoir lieu (Black, 2003). Des complexes multiprotéiques, appelés EJC (exon-exon junction complex), sont déposés après épissage au niveau des liaisons exon-exon. Ces complexes, jouent un rôle très important dans la maturation, l export nucléaire, la localisation cytoplasmique, la stabilité ainsi que la traductibilité des messagers (Le Hir and Seraphin, 2008). Des complexes multiprotéiques, appelés EJC (exon-exon junction complex), sont déposés après épissage au niveau des liaisons exon-exon. Ces complexes, jouent un rôle très important dans la maturation, l export nucléaire, la localisation cytoplasmique, la stabilité ainsi que la traductibilité des messagers (Le Hir and Seraphin, 2008). 18
23 2-2- L épissage alternatif L épissage alternatif est le processus selon lequel plusieurs ARNm peuvent être générés à partir d un même ARN pré-m, par l intermédiaire d un choix possible entre plusieurs sites 5 et/ou 3 d épissage. Ainsi, parmi les différents mécanismes associés à la transcription, que sont : l utilisation alternative de sites d initiation de la transcription, la maturation alternative en 3, l édition des ARN pré-m et l épissage alternatif, ce dernier est considéré comme la source la plus 19
24 importante de diversité des protéines chez les vertébrés (Maniatis and Reed, 2002). La prévalence de l épissage alternatif semble bien plus importante que ne l avaient indiquée les premières estimations. Des analyses plus récentes des ARNm dérivant du chromosome 22 humain indiquent qu environ 60% des gènes présentent deux ou plusieurs ARNm (Modrek and Lee, 2002). A titre d exemple, chez la Drosophile, l épissage alternatif du transcrit primaire Dscam (Down syndrom cell-adhesion molecule) peut générer, en théorie, ARNm différents (Schmucker et al., 2000). Les protéines qui régulent l épissage alternatif se lient à des séquences exoniques ou introniques activatrices (ESE ou ISE) ou inhibitrices (ESS ou ISS). La plupart des ESEs sont reconnues par des protéines SR (pour riches en dipeptides sérine/arginine (Graveley, 2000). Les protéines SR sont des facteurs ubiquitaires et multifonctionnels de l épissage constitutif et alternatif, essentiels aux différentes étapes de l assemblage du spliceosome. Elles facilitent la reconnaissance des extrémités 5 et 3 de l exon par la machinerie d'épissage, conduisant à la définition de l exon [concept de l "exon definition", (Berget, 1995)]. D'autres éléments régulateurs de la machinerie d'épissage appartiennent à la famille des hnrnp. Ces protéines ont un domaine de liaison à l ARN, de type RRM, comme les protéines SR, ou de type KH (pour "hnrnp K-Homology"), (Maniatis and Tasic, 2002). Ce domaine est suivi d une région plus variable qui peut être impliquée dans des interactions avec d autres protéines. Les protéines hnrnp ont une fonction générale d empaquetage des transcrits primaires et jouent des rôles plus spécifiques dans la maturation de certains ARNm (Maniatis and Tasic, 2002). Enfin, certains régulateurs de l épissage n appartiennent ni à la famille des protéines SR, ni à celle des hnrnp. 3- La maturation en 3 À l exception des transcrits codants pour les histones, tous les messagers eucaryotes comportent une queue polyadénosine à leur extrémité 3'. La maturation en 3 (clivage et polyadénylation) constitue une étape clé dans le contrôle de l expression génique en affectant la stabilité, l export et la traductibilité des transcrits. Cette modification co-transcriptionnelle comporte deux étapes successives essentielles, que sont le clivage du ARN pré-m et la polyadénylation du produit 5 de clivage (Figure 3). L hexanucléotide AAUAAA et l élément riche en GU, respectivement localisés en amont et en aval du site de clivage CA, représentent les éléments de séquence indispensables au recrutement des différentes protéines qui constituent le complexe de maturation en 3 (Zhao et al., 1999). Sur ces éléments en cis se fixent des facteurs en trans pour former le complexe de clivage. 20
25 L hexanucléotide conservé est reconnu par la sous-unité CPSF 160 du facteur spécifique de clivage et de polyadénylation (cleavage-polyadenylation specificity factor CPSF) tandis que l élément riche en GU est reconnu par la sous-unité CstF 64 du facteur de stimulation du clivage (CstF). Le complexe de clivage est également composé des facteurs de clivage (cleavage factor I et II CF I et CF II) ainsi que de la poly(a)polymérase (poly(a) polymerase PAP). La polyadénylation est assurée par la PAP qui ajoute des résidus adénosines en 3 du produit 5 de clivage. Cette étape requiert la présence du complexe CPSF et de la protéine de liaison nucléaire à la queue polyadénosine (poly(a)-binding protein II, PABP II). Le transcrit mature comporte, au final, une queue poly(a) constituée d environ 250 résidus adénosines. La maturation alternative en 3 consiste au choix entre deux ou plusieurs sites de clivage et de polyadénylation au sein d un transcrit primaire et il en résulte une différence de la région 3 non traduite (3 UTR) ou de la phase codante des messagers matures issus d un même ARN pré-m (Edwalds-Gilbert et al., 1997). Plusieurs études suggèrent que ce mécanisme de maturation en 3' est étroitement couplé aux autres processus de maturation des transcrits. Ces études supportent le modèle de définition des exons développé par Berget (Berget, 1995). Ce modèle suggère que les composants de la machinerie d épissage interagissent avec ceux impliqués dans la maturation en 3 définissant ainsi l exon 3' terminal d un pré-messager. 21
26 C- Le contrôle de la stabilité des ARN messagers Le contrôle de la stabilité (demi-vie) des messagers est un point essentiel dans la régulation de l expression génique en réponse à des stimuli intra- ou extra- cellulaires. Les extrémités 5 et 3 de l ARNm interviennent déjà dans sa protection contre la dégradation. En effet, la liaison du complexe formé par les protéines CBP20 et CBP80 à la coiffe, ainsi que la liaison de la protéine cytoplasmique PABPI (poly(a)-binding protein I) à la queue poly(a), représentent un encombrement stérique important qui constitue un obstacle aux 5 3 et 3 5 exonucléases. Par ailleurs, des éléments de séquence riches en dinucléotides AU (AU-rich elements AREs) capables de contrôler la demi-vie des ARNm ont été caractérisés dans les régions 3 UTR des ARNm. Parmi ces éléments, certains sont composés de séquences consensus pentamériques AUUUA, d autres de séquences nonamériques UUAUUUA(A/U)(A/U), ou plus simplement de motifs U-riches. Ces éléments en cis peuvent constituer le site de liaison d un ou plusieurs facteurs en trans appelés ARE-BPs (AU-rich elements binding proteins). La protéine AUF1 (AU-rich element RNA binding protein 1, aussi connue sous le nom de hnrnpd, pour heterogeneous nuclear ribonucleoprotein D), est une protéine nucléaire capable le stimuler ou d inhiber la dégradation des ARNm. Parmi les protéines appartenant à la super-famille des protéines elav (elav-related proteins), la liaison de la protéine HuR à des ARE induit la stabilisation des ARNm liés. D autres ARE-BPs régulent la stabilité des messagers. En fonction des besoins cellulaires, la demi-vie des ARNm va être prolongée ou écourtée. Dans ce dernier cas, la dégradation des ARNm va avoir lieu dans l exosome ou dans les P-bodies. (Barreau et al., 2005). D- La traduction La traduction des ARNm est une étape de l expression génique qui se déroule dans le cytoplasme des cellules eucaryotes (ou dans l appareil de Golgi, pour les protéines sécrétées). Comme les étapes précédentes, elle est soumise à de nombreuses régulations, dont seulement certaines seront évoquées. Seule la régulation de la traduction par les microarn sera approfondie. Avant d être traduits dans le cytoplasme, les ARNm matures doivent franchir les pores nucléaires, qui sont des structures multiprotéiques ancrées dans l enveloppe nucléaire. Ce processus, appelé export des ARNm, met en œuvre de nombreuses protéines, notamment celles de l EJC et du complexe CBP20/CBP80 (Proudfoot et al., 2002). 22
27 Comme pour la transcription, la traduction se déroule en trois étapes que sont l initiation, l élongation et la terminaison. L efficacité de la traduction pour un ARNm donné est déterminée par l étape d initiation de sa traduction. Cette étape joue donc un rôle déterminant dans la régulation de l expression génique. Elle fait intervenir de nombreux partenaires protéiques impliqués dans une série complexe d étapes successives qui ne seront que brièvement décrites dans le chapitre suivant. 1-Mécanismes d initiation de la traduction 1-1 Initiation à la coiffe L initiation de la traduction à la coiffe peut être schématiquement résumée à l enchaînement des étapes suivantes (Figure 4) (Holcik and Pestova, 2007): 1- Dissociation des sous-unités ribosomiques grâce à la liaison des facteurs (eif1a+eif1+eif3) à la sous-unité 40S (40S) et à l association d eif6 à la 60S (eif, pour eukaryotic translation initiation factor). 2- Formation du complexe 43S de pré-initiation, regroupant : eif2+gtp+met-arnti+40s (Met-ARNti : ARN de transfert- méthionine initiateur) 3- Recrutement du complexe 43S à la coiffe de l ARNm grâce au complexe eif4f 4- Balayage de la région 5 UTR de l ARNm 5- Reconnaissance du codon initiateur et relargage des facteurs précédemment associés à la 40S 6- Arrivée de la 60S, le ribosome 80S formé peut donc débuter la traduction Comme pour la transcription, la traduction se déroule en trois étapes que sont l initiation, l élongation et la terminaison. L efficacité de la traduction pour un ARNm donné est déterminée par l étape d initiation de sa traduction. Cette étape joue donc un rôle déterminant dans la régulation de l expression génique. Elle fait intervenir de nombreux partenaires protéiques impliqués dans une série complexe d étapes successives qui ne seront que brièvement décrites dans le chapitre suivant. Comme pour la transcription, la traduction se déroule en trois étapes que sont l initiation, l élongation et la terminaison. L efficacité de la traduction pour un ARNm donné est déterminée par l étape d initiation de sa traduction. Cette étape joue donc un rôle déterminant dans la régulation de l expression génique. Elle fait intervenir de nombreux partenaires protéiques impliqués dans une série complexe d étapes successives qui ne seront que brièvement décrites dans le chapitre suivant. 23
28 S (40S) 24
29 2- Le facteur eif2 est un trimère αβγ appartenant à la famille des protéines G. La sous-unité γ lie le GTP et le Met-ARNti. La sous-unité β contient la fonction catalytique GTPase. Enfin, la sousunité α est l unité régulatrice de ce trimère. Sa phosphorylation au niveau de son résidu sérine en position 51, décrite en conditions de stress, inhibe l activité catalytique de la sous-unité β, empêchant ainsi l initiation de la traduction. Cette phosphorylation est produite par différentes kinases : PERK (pancreatic endoplasmic reticulum eif2 kinase/eif2ak3), ou encore par PKR (double-stranded RNA-activated protein kinase/ EIF2AK2), ou encore par GCN2 (general control of nitrogen metabolism kinase 2/ EIF2AK4), ou enfin par EIF2AK1/HRI (heme regulated initiation factor 2 alpha kinase). 1- Le recrutement du complexe 43S à la coiffe se fait grâce au complexe eif4f et aux facteurs, eif4b et eif4h (Figure 5). L association de ces différents acteurs constitue le complexe 48S. Le complexe eif4f comporte trois facteurs : (1) le facteur eif4e qui lie la coiffe ; (2) le facteur eif4a (ARN hélicase ATP dépendante), qui déplie les structures secondaires de l ARN pour le balayage de la région 5 UTR ; (3) le facteur eif4g est une protéine d échafaudage qui relie d une part eif4e au complexe 43S et, d une autre part, la PABP au complexe d initiation de la traduction. Cette dernière interaction permet la circularisation de l ARNm en cours de traduction, mettant les régions 5 et 3 UTR à proximité les unes des autres. Enfin, les facteurs eif4b et eif4h stimulent l activité hélicase d eif4a. 25
30 4- Le balayage de la région 5 UTR par le complexe 48S dans le sens 5 3 met en œuvre l activité hélicase eif4a, jusqu à la rencontre du codon initiateur situé dans un «bon contexte nucléotidique». 5- La reconnaissance du codon initiateur se fait sous le contrôle des facteurs eif1 et eif1a (Pestova et al., 1998). Le facteur eif1 joue un rôle déterminant dans le choix du codon initiateur, alors que le facteur eif1a augmente l affinité d eif1 pour le complexe 43S-ARN et participe au changement de conformation du complexe ribosomique au niveau du codon initiateur. Lors de ce changement de conformation, eif1a avec eif5 participent à faire passer le complexe ribosomique d une conformation «ouverte» à une conformation dite «fermée», fixée devant le codon initiateur choisi par eif1 (Maag et al., 2006). 6- L assemblage du ribosome 80S se fait au niveau du site d initiation de la traduction. Au cours de cette étape le facteur eif5 stimule l hydrolyse du GTP par eif2, ce qui conduit dans un premier temps à la dissociation du complexe 43S pour, dans un second temps, permettre à la sousunité 60S de rejoindre la 40S et ainsi former le ribosome 80S. Cette association va positionner le Met-ARNti au niveau du site P du ribosome, la synthèse peptidique va alors pouvoir débuter Initiation par entrée interne des ribosomes (IRES) L initiation de la traduction par entrée interne des ribosomes, appelée IRES (pour internal ribosome entry site), représente un mécanisme alternatif à l initiation de la traduction initiée à la coiffe. Ce mécanisme fait intervenir des séquences particulières, contenues dans la région 5 UTR de certains ARNm ; celles-ci se présentent sous forme de structures secondaires en tige-boucle avec, éventuellement, des structures tertiaires produites par l enchevêtrement des premières. Au cours de l initiation de la traduction au niveau de ces structures, la sous-unité 40S ne passe pas par la coiffe, pas plus qu elle n effectue de balayage de la région 5 UTR de l ARNm, et vient se positionner directement au niveau du codon initiateur, situé à l extrémité 3 des IRES. Par conséquent, l initiation de la traduction par entrée interne des ribosomes est indépendante du facteur eif4e. Les IRES ont été initialement découvertes chez les virus (picornavirus, encephalomyocarditis virus), qui détournent la machinerie de la traduction de la cellule qu ils infectent en inhibant l initiation de la traduction initiée à la coiffe, pour mieux profiter ensuite des ribosomes rendus disponibles. Le facteur eif4g a été démontré comme étant la cible des protéases Lb et 2A de certains picornavirus libérant deux fragments : un petit N-terminal qui lie eife4 et un grand fragment de digestion liant eif4a avec eif3, capable de se fixer à la sous-unité 40S. Cette digestion 26
31 suffit pour inhiber la traduction initiée à la coiffe au profit de celle initiée aux IRES (Borman et al., 1997). Cependant, il n existe pas de mécanisme général commun pour définir l initiation de la traduction aux IRES, comme c est le cas pour l initiation de la traduction à la coiffe. Depuis leur découverte chez les virus, plus de 60 IRES virales et 90 IRES cellulaires ont été caractérisées à ce jour ( Le nombre croissant d IRES caractérisées a permis, au fur et à mesure, de comprendre les grandes lignes directrices qui régissent le(s) mécanisme(s) d initiation de la traduction aux IRES. Le recrutement du ribosome fait intervenir certains facteurs d initiation (à l exception d eif4e) ainsi que d autres facteurs en trans appelés ITAFs (IRES trans-acting factors) [Figure 6]. Certains de ces ITAFs sont indispensables au fonctionnement de toutes les IRES, alors que d autres sont plus spécifiques à certaines IRES particulières. Parmi les ITAFs «générales», la PTB (polypyrimidine-tractbinding protein) semble requise au recrutement du ribosome (Mitchell et al., 2005). D autres ITAFs comme hnrnpa1 (heterogeneous nuclear ribonucleoprotein A1) (Bonnal et al., 2005), La et unr (Mitchell et al., 2003) ont été caractérisées. Ces protéines semblent agir comme des «chaperonnes pour ARN» permettant à l IRES d adopter une configuration «active». Le champ d investigation sur le rôle des ITAFs et l identification de celles qui demeurent inconnues, demeure vaste. Cependant, il est important de noter que les IRES ont souvent été retrouvées mises à contribution dans des conditions de stress cellulaires, pendant lesquels la machinerie de traduction dépendante de la coiffe est mise en berne. Ceci a été illustré, entre autres, au cours de l apoptose dans une étude qui démontre que seuls 3 à 5% des messagers cellulaires sont traduits (associés aux 27
32 polysomes) et que parmi ces transcrits traduits, nombreux sont ceux qui possèdent des IRES fonctionnelles. De manière intéressante, les auteurs de cette étude ont observé une augmentation de l expression de la PTB dans les mêmes conditions d apoptose (Bushell et al., 2006). Des observations similaires ont été réalisées dans d autres conditions de stress, telle que l hypoxie (Cf II D-2-1), ou encore au cours de la mitose. C est pourquoi certains suggèrent que l initiation de la traduction aux IRES n est pas un simple mécanisme restreint à une poignée de transcrits. Ce serait un mécanisme alternatif d initiation de la traduction qui permettrait en conditions de stress, une fois la traduction initiée à la coiffe inhibée, de sélectionner les ARNm à traduire selon qu ils contiennent une, plusieurs ou aucune IRES (Spriggs et al., 2008). La recherche sur les IRES souffre surtout du petit nombre d IRES cellulaires caractérisées (environ 90 IRES décrites depuis 1991). Ceci tient au fait que ces IRES ne présentent aucune homologie de structure ou de séquence. L identification de chaque IRES se fait de manière individuelle, à l aide de systèmes basés sur l utilisation de constructions plasmidiques bicistroniques (Figure 7). Ces constructions possèdent deux cadres ouverts de lectures (cistrons) qui codent des gènes rapporteurs. Les deux cistrons sont séparés par la séquence contenant une IRES putative. L expression du premier cistron permet de mesurer l initiation de la traduction à la coiffe, alors que celle du second permet de mesurer l efficacité de la traduction initiée à l IRES. 28
33 Mais ce système souffre de nombreux travers, dont la présence potentielle de promoteurs cryptiques ou de sites d épissage alternatifs dans la séquence à tester. L utilisation de ce système doit donc être couplée à des expériences de northern blot, de protection d ARN, etc pour s assurer de l absence d éventuels transcrits «remaniés» qui viendraient fausser les mesures. L ensemble de ces difficultés ne vient certainement pas aider à étendre le champ des IRES, encore nombreuses à découvrir. 2-Régulation de la traduction Il ne s agira pas dans ce chapitre de dresser la liste complète de l ensemble de toutes les régulations décrites affectant l initiation de la traduction, mais seulement les plus importantes d entre elles. La régulation de la traduction par les microarn sera abordée ultérieurement. Étant donné tant le vaste champ d action de ces molécules sur la régulation de l expression génique que leur importance dans le cadre de cette thèse, un chapitre entier leur a été consacré Au niveau des facteurs d initiation de la traduction Comme il a été précédemment mentionné, l initiation de la traduction peut être régulée par la phosphorylation de la sérine 51 de la sous-unité α du facteur eif2. De plus l initiation de la traduction est régulée par (1) la quantité du facteur eif4e, (2) la régulation de son activité par la phosphorylation de son résidu sérine 209, et (3) par la séquestration de ce facteur par les membres de la famille des 4E-BPs (eif4e binding proteins). 1- Le niveau d expression du facteur eif4e est fondamental puisqu il est nettement inférieur (10 à 50 fois) à celui de tous les autres membres du contingent de l initiation de la traduction (Duncan et al., 1987). Toute modification de son niveau d expression est cruciale pour la régulation de l initiation de la traduction initiée à la coiffe. 2- La régulation de l activité du facteur eif4e par la phosphorylation de son résidu sérine 209 se fait en réponse à des stimuli apportés par des facteurs de croissance, des hormones, etc Cette phosphorylation est assurée par la protéine Mnk1 (MAP-kinase-interacting-kinase-1) qui s associe à l extrémité carboxy-terminale du facteur eif4g (Pyronnet et al., 1999). La phosphorylation d eif4e par la protéine Mnk1 augmente son affinité pour la coiffe, c est pourquoi l hypo-phosphorylation d eif4e au cours de la mitose, s accompagne d une inhibition de l initiation de la traduction initiée à la coiffe (Pyronnet et al., 2001). 3- Enfin, la disponibilité du facteur eif4e est très importante dans la régulation de l initiation de la traduction initiée à la coiffe. 29
34 En effet, au cours du processus d initiation, eif4e reconnaît la coiffe et interagit avec la protéine d échafaudage eif4g, qui elle-même interagit avec le complexe 43S, permettant à ce dernier de s associer indirectement à la coiffe. En empêchant la liaison eif4e-eif4g, l initiation de la traduction à la coiffe est rendue impossible. Ceci est assuré par les protéines de liaison au facteur eif4e membres de la famille des 4E-BPs qui entrent en compétition avec eif4g (encore appelé p220) au niveau de son site de fixation sur eif4e (Haghighat et al., 1995). Cette interaction entre eif4e et les 4E-BPs est régulée par la phosphorylation des 4E-BPs. En conditions de stress, la déphosphorylation des 4E-BPs permet leur liaison à eif4e, inhibant ainsi l initiation de la traduction initiée à la coiffe. Inversement, la stimulation de cette dernière est associée à une phosphorylation des 4E-BPs, essentiellement assurée par la protéine mtor (mammalian Target of Rapamycin, encore connue sous le nom de FRAP, FKBP and rapamycin-associated protein) Par choix de codons alternatifs d initiation de la traduction Le choix du codon initiateur de la traduction se fait sous le contrôle de la machinerie traductionnelle en trans (eif1 et eif1a), dans un contexte nucléotidique qui fut décrit par Marilyn Kozak (Kozak, 1986). Dans ce contexte (5 -GCCRCCAUGG-3 ) la base A du codon AUG initiateur correspond à la position +1 de la phase codante, et elle semble définie par une base purique en position -3 et une guanine en position +4. Toutefois, l initiation de la traduction peut s effectuer au niveau d autres codons, dits non canoniques, non AUG : CUG, ACG ou CUG. L efficacité de l initiation de la traduction initiée à ce type de codon est très variable ; elle semble être augmentée par la richesse en dinucléotide GC et par la présence de structures en épingle à cheveux (ou hairpin) présentes en aval de ce type de codon qui ralentiraient le balayage du ribosome (Kozak, 1990). L utilisation de ce type de codons alternatifs introduit à côté des autres mécanismes alternatifs (épissage, maturation en 3, etc ) une nouvelle source de diversité, permettant de synthétiser différentes protéines à partir du même transcrit. L utilisation de codons alternatifs en phase avec le codon AUG permet de produire une protéine rallongée ou tronquée selon qu il soit situé en amont ou en aval de ce dernier, respectivement Par la présence de petits cadres ouverts de lecture La présence de petits cadres de lecture situés dans la région 5 UTR en amont du codon AUG principal peut réguler l initiation de la traduction au niveau de ce dernier. Ces petits cadres de lecture, appelés uaug (upstream AUG), ont été particulièrement retrouvés dans les régions 30
35 5 UTR de transcrits codant des facteurs de croissance, de différenciation cellulaire, de transcription En fonction du contexte nucléotidique dans lequel est contenu le uaug, deux cas de figures peuvent se produire : - si le uaug est dans un contexte peu favorable, le complexe de préinitiation va préférentiellement poursuivre le balayage de la région 5 UTR à la recherche d un codon initiateur plus favorable. C est le processus de «fuite», ou de «balayage relâché». - si le uaug est dans un contexte favorable, une initiation de la traduction va avoir lieu en ce point, produisant une uorf (upstream open reading frames). Une fois la traduction de cette uorf finie, seuls certains ribosomes restés associés au messager poursuivent le balayage pour réinitier la traduction au niveau du codon initiateur du cadre de lecture principal. C est le processus de terminaison-réinitiation. Dans les deux cas, la présence de uaug permet la traduction de uorf qui conduit à une atténuation, voire une plus importante diminution de l efficacité de la traduction du cadre de lecture principal (Sachs and Geballe, 2006). E- Régulation de l expression de l expression génique par les micro-arn Les microarn (miarn) sont des ARN non codants de 21 à 23 nucléotides de long, qui ont récemment fait leur entrée dans la catégorie des régulateurs de l expression génique. Au cours de ces cinq dernières années, les miarn se sont imposés comme des éléments majeurs du contrôle de l expression génique au niveau post-transcriptionnel. Initialement découverts chez Caenorhabditis elegans, en 1993, les miarn étaient, jusqu à présent, considérés comme des régulateurs exclusifs des organismes multicellulaires. Ceci a amené certains chercheurs à penser que les miarn apportent un niveau de complexité supplémentaire dans la régulation de l expression génique distinguant les organismes multicellulaires des simples organismes unicellulaires. Cependant la découverte récente de l existence des miarn dans la Chlamydomonas reinhardtii, une algue unicellulaire, indique que les miarn sont probablement issus d un processus que l évolution aurait choisi de conserver (Molnar et al., 2007). Actuellement, chez l homme, on dénombre 695 mirna dans les banques de données ( Les études fonctionnelles réalisées sur les miarn indiquent que ces petits ARN participent dans la régulation de quasiment tous les processus cellulaires, puisque toute altération dans leur niveau d expression est associée à des pathologies (cancers, maladies 31
36 cardiovasculaires ) (Kloosterman and Plasterk, 2006). Ces résultats ne sont pas surprenants lorsqu ils sont mis en perspective avec ceux obtenus par les bioinformaticiens, dont les prédictions évaluent à près de 30% l ensemble des gènes codant pour des protéines potentiellement régulées par des microarn. 1- Biogenèse des microarn La genèse des microarn se fait en quatre étapes successives : (1) transcription du primicroarn, (2) clivage par Drosha pour libérer un précurseur pré-microarn, (3) export de ce dernier dans le cytoplasme et (4) clivage du pré-microarn par Dicer (Figure 8). La majorité des gènes codant les microarn sont transcrits par l ARN polymérase II, qui produit une molécule d ARN pro-précurseur en forme de tige-boucle (avec quelques mésappariements dans la tige), d une centaine des nucléotides, appelé pri-pré-microarn (Lee et al., 2004). Cette molécule est ensuite prise en charge par un complexe multiprotéique appelé Microprocessor composé des protéines Drosha et DGCR8 (chez les vertébrés, ou Pasha chez les invertébrés), (Seitz and Zamore, 2006). Drosha est une ribonucléase de type III agissant de concert avec DGCR8 (protéine de liaison à l ARN double brin) pour libérer un précurseur d environ 70 nucléotides ; ce précurseur forme toujours une structure en tige-boucle avec une extrémité 3 sortante (avec 2 nucléotides qui dépassent, 2-nt 3 overhang). Cette dernière caractéristique représente une «empreinte» de la digestion catalysée par les ribonucléases de type III. La majorité des miarn seraient contenus dans les introns de ARNm, et une étude montre que l excision de ces pri-pré-microarn par Drosha interviendrait sur l ARN en cours de transcription avant même l épissage des introns «hôtes» (Kim and Kim, 2007). Ce précurseur serait ensuite activement exporté dans le cytoplasme par l Exportine-5. Celle-ci reconnaît le pré-microarn grâce à son extrémité 3 sortante, et l exporte vers le cytoplasme via un mécanisme Ran-GTP-dépendant (Lund et al., 2004). Dans le cytoplasme, le pré-microarn va subir sa dernière étape de maturation, catalysée par Dicer, une autre ribonucléase de type III, assistée de la protéine TRBP (pour TAR RNA binding protein). Le microarn résultant est un duplexe de 21 nucléotides en moyenne avec deux extrémités 3 sortantes (2-nt 3 overhang) similaire aux petits ARNs opérant dans le mécanisme d ARN interférence (small interfering RNAs, sirna). Le miarn mature serait issu du brin de ce duplexe, qui aurait l extrémité 5 la moins stable d un point de vue thermodynamique (Khvorova et al., 2003). Ce miarn simple brin sera ensuite intégré dans le complexe ribonucléoprotéique appelé RISC (pour RNA-induced silencing complex) ou encore appelé mirisc (pour mirna-induced 32
37 silencing complex) afin de distinguer le mécanisme d ARN interférence faisant intervenir les miarn de celui qui fait intervenir les siarn. 33
38 Le complexe mirisc est un complexe ribonucléoprotéique qui contient le miarn mature, les protéines Dicer et TRBP auxquelles sont venues se joindre les protéines Argonautes. Les membres de la famille des protéines Argonautes (AGO) sont les composants les plus caractérisés des complexes mirisc. Chez les mammifères, la répression de la synthèse protéique amenée par les miarn via le complexe mirisc dépend de la présence des protéines AGO1-4 (Pillai et al., 2004). Par contre, lorsqu un miarn induit le clivage de l ARNm dont il lie la région 3 UTR, seule Ago2 peut assurer cette fonction dans le mirisc, car c est le seul membre de la famille des protéines Argonautes à posséder une fonction ribonucléase apportée par son domaine PIWI (RNaseH-like P- element induced wimpy testis) (Peters and Meister, 2007). 2- Principes de l interaction miarn-arnm Les microarn interagissent avec la région 3 UTR des ARN messagers par complémentarité de base entre les deux éléments. Chez les plantes, la complémentarité entre le miarn et l ARNm est presque parfaite. Ceci engendre la dégradation de l ARNm cible suivant un processus semblable au mécanisme d ARN interférence, avec un clivage endonucléocytique qui se produit au milieu du duplex miarn-arnm (Jones-Rhoades et al., 2006). Chez les métazoaires, les bases de l appariement entre miarn et ARNm sont tout autres, puisque la complémentarité entre le miarn et l ARNm est imparfaite. Cependant, cet appariement suit des règles précises, dont certaines sont indispensables à la répression exercée par le miarn sur l ARNm. La synthèse des différents exemples de duplexes miarn-arnm effectifs décrits dans la littérature (Grimson et al., 2007), associée aux efforts de la bioinformatique pour dégager des règles pour construire des algorithmes utilisés pour la prédiction de sites de microarn, a permis de définir les règles suivantes: 1- L hybridation entre le miarn et l ARNm doit être parfaite entre les nucléotides 2 et 8 du miarn. C est ce qui constitue le noyau de nucléation du duplexe miarn-arnm, appelé seed pour graine, car c est à ce niveau que l appariement s initie, qu il germe. Tout mésappariement, ou appariement de type G:U, à ce niveau, affecte considérablement l efficacité de la répression (Doench and Sharp, 2004). De plus, un résidu A, en position 1 du miarn, et /ou un résidu A/U en position 9 du miarn, c est-à-dire de part et d autre du seed augmenterait l efficacité de la répression médiée par la liaison du microarn à l ARNm (Lewis et al., 2005). Toutefois, la 34
Contrôle de l'expression génétique : Les régulations post-transcriptionnelles
 Contrôle de l'expression génétique : Les régulations post-transcriptionnelles http://perso.univ-rennes1.fr/serge.hardy/ utilisateur : biochimie mot de passe : 2007 L'ARNm, simple intermédiaire entre le
Contrôle de l'expression génétique : Les régulations post-transcriptionnelles http://perso.univ-rennes1.fr/serge.hardy/ utilisateur : biochimie mot de passe : 2007 L'ARNm, simple intermédiaire entre le
Contrôle de l'expression génétique :
 Contrôle de l'expression génétique : Les régulations post-transcriptionnelles L'ARNm, simple intermédiaire entre le génome et les protéines? gène protéine L'ARNm, simple intermédiaire entre le génome et
Contrôle de l'expression génétique : Les régulations post-transcriptionnelles L'ARNm, simple intermédiaire entre le génome et les protéines? gène protéine L'ARNm, simple intermédiaire entre le génome et
Tableau 1. Liste (non exhaustive) des protéines se localisant dans les P-Bodies
 des protéines se localisant dans les P-Bodies") NOM FONCTION EFFET DE L ABSENCE OU DE REFERENCES LA SUREXPRESSION SUR LES P- BODIES XRN1 exonucléase 5 3 Absence : augmente la taille et le Bashkirov et al. 1997 Shet et Parker 2003 Cougot nombre des P-bodies
NOM FONCTION EFFET DE L ABSENCE OU DE REFERENCES LA SUREXPRESSION SUR LES P- BODIES XRN1 exonucléase 5 3 Absence : augmente la taille et le Bashkirov et al. 1997 Shet et Parker 2003 Cougot nombre des P-bodies
CHAPITRE 3 LA SYNTHESE DES PROTEINES
 CHAITRE 3 LA SYNTHESE DES ROTEINES On sait qu un gène détient dans sa séquence nucléotidique, l information permettant la synthèse d un polypeptide. Ce dernier caractérisé par sa séquence d acides aminés
CHAITRE 3 LA SYNTHESE DES ROTEINES On sait qu un gène détient dans sa séquence nucléotidique, l information permettant la synthèse d un polypeptide. Ce dernier caractérisé par sa séquence d acides aminés
VI- Expression du génome
 VI- Expression du génome VI-1.- EXPRESSION DU GÉNOME- PRINCIPES GÉNÉRAUX DOGME CENTRAL Les gènes et l information génétique sont conservés sous forme d acides nucléiques La perpétuation à l identique de
VI- Expression du génome VI-1.- EXPRESSION DU GÉNOME- PRINCIPES GÉNÉRAUX DOGME CENTRAL Les gènes et l information génétique sont conservés sous forme d acides nucléiques La perpétuation à l identique de
Module 5 La maturation de l ARN et le contrôle post-transcriptionnel chez les eucaryotes
 Module 5 La maturation de l ARN et le contrôle post-transcriptionnel chez les eucaryotes Où trouver l'information complémentaire? MCB -11, GVII-5, 22, 23. La maturation des ARNm chez les eucaryotes Les
Module 5 La maturation de l ARN et le contrôle post-transcriptionnel chez les eucaryotes Où trouver l'information complémentaire? MCB -11, GVII-5, 22, 23. La maturation des ARNm chez les eucaryotes Les
Chapitre 7 : Structure de la cellule Le noyau cellulaire
 UE2 : Structure générale de la cellule Chapitre 7 : Structure de la cellule Le noyau cellulaire Professeur Michel SEVE Année universitaire 2010/2011 Université Joseph Fourier de Grenoble - Tous droits
UE2 : Structure générale de la cellule Chapitre 7 : Structure de la cellule Le noyau cellulaire Professeur Michel SEVE Année universitaire 2010/2011 Université Joseph Fourier de Grenoble - Tous droits
ULBI 101 Biologie Cellulaire L1. Le Système Membranaire Interne
 ULBI 101 Biologie Cellulaire L1 Le Système Membranaire Interne De la nécessité d un SMI Le volume augmente comme le cube de la dimension linéaire, alors que la surface n'est augmentée que du carré Une
ULBI 101 Biologie Cellulaire L1 Le Système Membranaire Interne De la nécessité d un SMI Le volume augmente comme le cube de la dimension linéaire, alors que la surface n'est augmentée que du carré Une
TD de Biochimie 4 : Coloration.
 TD de Biochimie 4 : Coloration. Synthèse de l expérience 2 Les questions posées durant l expérience 2 Exposé sur les méthodes de coloration des molécules : Générique Spécifique Autres Questions Pourquoi
TD de Biochimie 4 : Coloration. Synthèse de l expérience 2 Les questions posées durant l expérience 2 Exposé sur les méthodes de coloration des molécules : Générique Spécifique Autres Questions Pourquoi
Dr E. CHEVRET UE2.1 2013-2014. Aperçu général sur l architecture et les fonctions cellulaires
 Aperçu général sur l architecture et les fonctions cellulaires I. Introduction II. Les microscopes 1. Le microscope optique 2. Le microscope à fluorescence 3. Le microscope confocal 4. Le microscope électronique
Aperçu général sur l architecture et les fonctions cellulaires I. Introduction II. Les microscopes 1. Le microscope optique 2. Le microscope à fluorescence 3. Le microscope confocal 4. Le microscope électronique
Les OGM. 5 décembre 2008. Nicole Mounier
 Les OGM 5 décembre 2008 Nicole Mounier Université Claude Bernard Lyon 1 CGMC, bâtiment Gregor Mendel 43, boulevard du 11 Novembre 1918 69622 Villeurbanne Cedex OGM Organismes Génétiquement Modifiés Transfert
Les OGM 5 décembre 2008 Nicole Mounier Université Claude Bernard Lyon 1 CGMC, bâtiment Gregor Mendel 43, boulevard du 11 Novembre 1918 69622 Villeurbanne Cedex OGM Organismes Génétiquement Modifiés Transfert
IMMUNOLOGIE. La spécificité des immunoglobulines et des récepteurs T. Informations scientifiques
 IMMUNOLOGIE La spécificité des immunoglobulines et des récepteurs T Informations scientifiques L infection par le VIH entraîne des réactions immunitaires de l organisme qui se traduisent par la production
IMMUNOLOGIE La spécificité des immunoglobulines et des récepteurs T Informations scientifiques L infection par le VIH entraîne des réactions immunitaires de l organisme qui se traduisent par la production
Le rôle de l endocytose dans les processus pathologiques
 UE7 Cours n 9 C. LAMAZE 24.11.11 Elise GODEAU (partie1) Guillaume MERGENTHALER (partie2) Le rôle de l endocytose dans les processus pathologiques SOMMAIRE : I. L endocytose à récepteurs : la voie des clathrines
UE7 Cours n 9 C. LAMAZE 24.11.11 Elise GODEAU (partie1) Guillaume MERGENTHALER (partie2) Le rôle de l endocytose dans les processus pathologiques SOMMAIRE : I. L endocytose à récepteurs : la voie des clathrines
Biomarqueurs en Cancérologie
 Biomarqueurs en Cancérologie Définition, détermination, usage Biomarqueurs et Cancer: définition Anomalie(s) quantitative(s) ou qualitative(s) Indicative(s) ou caractéristique(s) d un cancer ou de certaines
Biomarqueurs en Cancérologie Définition, détermination, usage Biomarqueurs et Cancer: définition Anomalie(s) quantitative(s) ou qualitative(s) Indicative(s) ou caractéristique(s) d un cancer ou de certaines
Université d Evry-Val d Essonne Ecole Doctorale des Génomes Aux Organismes. Thèse
 Université d Evry-Val d Essonne Ecole Doctorale des Génomes Aux Organismes Thèse Présentée pour obtenir le grade de Docteur en sciences de l université d Evry-Val d Essonne Spécialité Bioinformatique par
Université d Evry-Val d Essonne Ecole Doctorale des Génomes Aux Organismes Thèse Présentée pour obtenir le grade de Docteur en sciences de l université d Evry-Val d Essonne Spécialité Bioinformatique par
Cellules procaryotes Service histologie Pr.k.mebarek
 Cellules procaryotes Service histologie Pr.k.mebarek I) Les cellules procaryotes II) Les cellules eucaryotes o 1) Caractéristiques générales des cellules eucaryotes o 2) Organisation des cellules eucaryotes
Cellules procaryotes Service histologie Pr.k.mebarek I) Les cellules procaryotes II) Les cellules eucaryotes o 1) Caractéristiques générales des cellules eucaryotes o 2) Organisation des cellules eucaryotes
Séquence 2. L expression du patrimoine génétique. Sommaire
 Séquence 2 L expression du patrimoine génétique Sommaire 1. La synthèse des protéines 2. Phénotypes, génotypes et environnement Synthèse de la séquence 2 Exercices de la séquence 2 Glossaire des séquences
Séquence 2 L expression du patrimoine génétique Sommaire 1. La synthèse des protéines 2. Phénotypes, génotypes et environnement Synthèse de la séquence 2 Exercices de la séquence 2 Glossaire des séquences
Hépatite chronique B Moyens thérapeutiques
 Hépatite chronique B Moyens thérapeutiques Dr Olfa BAHRI Laboratoire de Virologie Clinique Institut Pasteur de Tunis INTRODUCTION Plus de 300. 10 6 porteurs chroniques de VHB dans le monde Hépatite chronique
Hépatite chronique B Moyens thérapeutiques Dr Olfa BAHRI Laboratoire de Virologie Clinique Institut Pasteur de Tunis INTRODUCTION Plus de 300. 10 6 porteurs chroniques de VHB dans le monde Hépatite chronique
COUSIN Fabien KERGOURLAY Gilles. 19 octobre 2007. de l hôte par les. Master 2 MFA Responsable : UE Incidence des paramètres environnementaux
 COUSIN Fabien KERGOURLAY Gilles 19 octobre 2007 Inhibition des défenses de l hôte par les bactéries pathogènes Master 2 MFA Responsable : UE Incidence des paramètres environnementaux Gwennola ERMEL I Détection
COUSIN Fabien KERGOURLAY Gilles 19 octobre 2007 Inhibition des défenses de l hôte par les bactéries pathogènes Master 2 MFA Responsable : UE Incidence des paramètres environnementaux Gwennola ERMEL I Détection
www.gbo.com/bioscience 1 Culture Cellulaire Microplaques 2 HTS- 3 Immunologie/ HLA 4 Microbiologie/ Bactériologie Containers 5 Tubes/ 6 Pipetage
 2 HTS 3 Immunologie / Immunologie Informations Techniques 3 I 2 ELISA 96 Puits 3 I 4 ELISA 96 Puits en Barrettes 3 I 6 en Barrettes de 8 Puits 3 I 7 en Barrettes de 12 Puits 3 I 8 en Barrettes de 16 Puits
2 HTS 3 Immunologie / Immunologie Informations Techniques 3 I 2 ELISA 96 Puits 3 I 4 ELISA 96 Puits en Barrettes 3 I 6 en Barrettes de 8 Puits 3 I 7 en Barrettes de 12 Puits 3 I 8 en Barrettes de 16 Puits
chez les eucaryotes, source de diversification et de modulation de l'expression des gènes
 L'initiation de la traduction chez les eucaryotes, source de diversification et de modulation de l'expression des gènes Olivier Jean-Jean, Michel Cassan, Jean-Pierre Rousset Société Française de Génétique
L'initiation de la traduction chez les eucaryotes, source de diversification et de modulation de l'expression des gènes Olivier Jean-Jean, Michel Cassan, Jean-Pierre Rousset Société Française de Génétique
I. La levure Saccharomyces cerevisiae: mode de vie
 LES LEVURES UE «levures» -5 avril: généralités (MN Simon) -6 avril: analyse génétique (MN Simon) -6 avril: Cycle cellulaire I: la réplication (E. bailly) -7 avril: Cycle cellulaire II: la mitose (E. Bailly)
LES LEVURES UE «levures» -5 avril: généralités (MN Simon) -6 avril: analyse génétique (MN Simon) -6 avril: Cycle cellulaire I: la réplication (E. bailly) -7 avril: Cycle cellulaire II: la mitose (E. Bailly)
Les cytokines et leurs récepteurs. Laurence Guglielmi laurence.guglielmi@univ-montp1.frli
 Les cytokines et leurs récepteurs Laurence Guglielmi laurence.guglielmi@univ-montp1.frli l i@ i 1 Les cytokines et leurs récepteurs 2 mécanismes principaux d interactions cellulaires : - contact membranaire
Les cytokines et leurs récepteurs Laurence Guglielmi laurence.guglielmi@univ-montp1.frli l i@ i 1 Les cytokines et leurs récepteurs 2 mécanismes principaux d interactions cellulaires : - contact membranaire
Les renseignements suivants sont destinés uniquement aux personnes qui ont reçu un diagnostic de cancer
 Information importante pour les personnes atteintes d un cancer du poumon non à petites cellules de stade avancé Les renseignements suivants sont destinés uniquement aux personnes qui ont reçu un diagnostic
Information importante pour les personnes atteintes d un cancer du poumon non à petites cellules de stade avancé Les renseignements suivants sont destinés uniquement aux personnes qui ont reçu un diagnostic
Travaux dirigés de Microbiologie Master I Sciences des Génomes et des Organismes Janvier 2015
 Andrew Tolonen atolonen@genoscope.cns.fr Travaux dirigés de Microbiologie Master I Sciences des Génomes et des Organismes Janvier 2015 A- Généralités I- La vie sur terre telle que nous la connaissons ne
Andrew Tolonen atolonen@genoscope.cns.fr Travaux dirigés de Microbiologie Master I Sciences des Génomes et des Organismes Janvier 2015 A- Généralités I- La vie sur terre telle que nous la connaissons ne
MYRIAD. l ADN isolé n est à présent plus brevetable!
 MYRIAD La Cour Suprême des Etats-Unis revient sur plus de 30 ans de pratique : l ADN isolé n est à présent plus brevetable! Mauvaise passe pour les inventions en biotechnologies sur le territoire américain.
MYRIAD La Cour Suprême des Etats-Unis revient sur plus de 30 ans de pratique : l ADN isolé n est à présent plus brevetable! Mauvaise passe pour les inventions en biotechnologies sur le territoire américain.
Chapitre II La régulation de la glycémie
 Chapitre II La régulation de la glycémie Glycémie : concentration de glucose dans le sang valeur proche de 1g/L Hypoglycémie : perte de connaissance, troubles de la vue, voire coma. Hyperglycémie chronique
Chapitre II La régulation de la glycémie Glycémie : concentration de glucose dans le sang valeur proche de 1g/L Hypoglycémie : perte de connaissance, troubles de la vue, voire coma. Hyperglycémie chronique
THESE. présentée DEVANT L UNIVERSITE DE RENNES I. Pour obtenir. Le grade de : DOCTEUR DE L UNIVERSITE DE RENNES I. Mention : Biologie PAR
 Avertissement Au vu de la législation sur les droits d'auteur, ce travail de thèse demeure la propriété de son auteur, et toute reproduction de cette oeuvre doit faire l'objet d'une autorisation de l'auteur.
Avertissement Au vu de la législation sur les droits d'auteur, ce travail de thèse demeure la propriété de son auteur, et toute reproduction de cette oeuvre doit faire l'objet d'une autorisation de l'auteur.
voies de signalisation
 Olivier Lascols Faculté de Médecine ierre et Marie Curie Généralités s sur les voies de signalisation 1 I. Caractérisation risation d une d voie de transduction d un d signal Exemple du Signal Mitogénique
Olivier Lascols Faculté de Médecine ierre et Marie Curie Généralités s sur les voies de signalisation 1 I. Caractérisation risation d une d voie de transduction d un d signal Exemple du Signal Mitogénique
Université de Montréal. Développement d outils pour l analyse de données de ChIP-seq et l identification des facteurs de transcription
 Université de Montréal Développement d outils pour l analyse de données de ChIP-seq et l identification des facteurs de transcription par Eloi Mercier Département de bioinformatique Faculté de médecine
Université de Montréal Développement d outils pour l analyse de données de ChIP-seq et l identification des facteurs de transcription par Eloi Mercier Département de bioinformatique Faculté de médecine
Bases moléculaires des mutations Marc Jeanpierre
 Bases moléculaires des mutations Marc Jeanpierre Chaque enfant qui naît hérite de 10 à 30 nouvelles mutations ponctuelles. L essentiel des ces mutations sont heureusement des variations neutres de séquence
Bases moléculaires des mutations Marc Jeanpierre Chaque enfant qui naît hérite de 10 à 30 nouvelles mutations ponctuelles. L essentiel des ces mutations sont heureusement des variations neutres de séquence
Annales de Biologie Cellulaire QCM (niveau SVT 1 er année)
") Annales de Biologie Cellulaire QCM (niveau SVT 1 er année) Equipe pédagogique Université Bordeaux-1 Didier Morin, Michel Moenner, Sophie North, Gérard Tramu et IJsbrand Kramer Contact : i.kramer@iecb.u-bordeaux.fr
Annales de Biologie Cellulaire QCM (niveau SVT 1 er année) Equipe pédagogique Université Bordeaux-1 Didier Morin, Michel Moenner, Sophie North, Gérard Tramu et IJsbrand Kramer Contact : i.kramer@iecb.u-bordeaux.fr
Univers Vivant Révision. Notions STE
 Univers Vivant Révision Notions STE Chap. 13) L Écologie 1) a) Qu est-ce que l empreinte écologique? L empreinte écologique correspond à la surface terrestre et aquatique totale nécessaire à un individu,
Univers Vivant Révision Notions STE Chap. 13) L Écologie 1) a) Qu est-ce que l empreinte écologique? L empreinte écologique correspond à la surface terrestre et aquatique totale nécessaire à un individu,
Mention : STAPS. Sport, Prévention, Santé, Bien-être. Objectifs de la spécialité
 Mention : STAPS Sport, Prévention, Santé, Bien-être Objectifs de la spécialité L'objectif de la spécialité «Sport, Prévention, Santé, Bien être» est de doter les étudiants de compétences scientifiques,
Mention : STAPS Sport, Prévention, Santé, Bien-être Objectifs de la spécialité L'objectif de la spécialité «Sport, Prévention, Santé, Bien être» est de doter les étudiants de compétences scientifiques,
CHAPITRE 3 LES TASQ COMME PLATEFORME POUR LA CATALYSE PSEUDO-ENZYMATIQUE
 L. Stefan Chapitre 3 CHAPITRE 3 LES TASQ COMME PLATEFORME POUR LA CATALYSE PSEUDO-ENZYMATIQUE Nous avons démontré dans le chapitre précédent que les TASQ hydrosolubles développés au cours de cette thèse
L. Stefan Chapitre 3 CHAPITRE 3 LES TASQ COMME PLATEFORME POUR LA CATALYSE PSEUDO-ENZYMATIQUE Nous avons démontré dans le chapitre précédent que les TASQ hydrosolubles développés au cours de cette thèse
Guide de rédaction d un protocole de recherche clinique à. l intention des chercheurs évoluant en recherche fondamentale
 V E R S I O N A V R I L 2 0 1 2 C E N T R E D E R E C H E R C H E C L I N I Q U E É T I E N N E - L E B E L D U C H U S Guide de rédaction d un protocole de recherche clinique à l intention des chercheurs
V E R S I O N A V R I L 2 0 1 2 C E N T R E D E R E C H E R C H E C L I N I Q U E É T I E N N E - L E B E L D U C H U S Guide de rédaction d un protocole de recherche clinique à l intention des chercheurs
CATALOGUE DES PRESTATIONS DE LA
 1/23 La plate-forme Biopuces et Séquençage de Strasbourg est équipée des technologies Affymetrix et Agilent pour l étude du transcriptome et du génome sur puces à ADN. SOMMAIRE ANALYSE TRANSCRIPTIONNELLE...
1/23 La plate-forme Biopuces et Séquençage de Strasbourg est équipée des technologies Affymetrix et Agilent pour l étude du transcriptome et du génome sur puces à ADN. SOMMAIRE ANALYSE TRANSCRIPTIONNELLE...
5.5.5 Exemple d un essai immunologique
 5.5.5 Exemple d un essai immunologique Test de grossesse Test en forme de bâtonnet destiné à mettre en évidence l'hormone spécifique de la grossesse, la gonadotrophine chorionique humaine (hcg), une glycoprotéine.
5.5.5 Exemple d un essai immunologique Test de grossesse Test en forme de bâtonnet destiné à mettre en évidence l'hormone spécifique de la grossesse, la gonadotrophine chorionique humaine (hcg), une glycoprotéine.
Annales du Contrôle National de Qualité des Analyses de Biologie Médicale
 Annales du Contrôle National de Qualité des Analyses de Biologie Médicale ARN du virus de l hépatite C : ARN-VHC ARN-VHC 03VHC1 Novembre 2003 Edité : mars 2006 Annales ARN-VHC 03VHC1 1 / 8 ARN-VHC 03VHC1
Annales du Contrôle National de Qualité des Analyses de Biologie Médicale ARN du virus de l hépatite C : ARN-VHC ARN-VHC 03VHC1 Novembre 2003 Edité : mars 2006 Annales ARN-VHC 03VHC1 1 / 8 ARN-VHC 03VHC1
Domaine : Sciences, Technologies et Santé Mention : Nutrition, Sciences des aliments, Agroalimentaire
 Contexte Domaine : Sciences, Technologies et Santé Mention : Nutrition, Sciences des aliments, Agroalimentaire Fédération des spécialités de Master des 5 pôles universitaires partenaires de la région Nord-Pas-de-Calais
Contexte Domaine : Sciences, Technologies et Santé Mention : Nutrition, Sciences des aliments, Agroalimentaire Fédération des spécialités de Master des 5 pôles universitaires partenaires de la région Nord-Pas-de-Calais
Présentation Générale
 Mars 2009 Présentation Générale 1- Le Master Recherche en Sciences de la Vie et de la Santé à Nice Objectifs, environnement scientifique, organisation Enseignements, les spécialités, les cours et les stages
Mars 2009 Présentation Générale 1- Le Master Recherche en Sciences de la Vie et de la Santé à Nice Objectifs, environnement scientifique, organisation Enseignements, les spécialités, les cours et les stages
Cancer du rein métastatique Nouvelles approches thérapeutiques. Sophie Abadie-Lacourtoisie 20 mai 2006
 Cancer du rein métastatique Nouvelles approches thérapeutiques Sophie Abadie-Lacourtoisie 20 mai 2006 Cancer du rein à cellules claires métastatique (CRM) Plus de 30 % des CR deviendront métastatiques
Cancer du rein métastatique Nouvelles approches thérapeutiques Sophie Abadie-Lacourtoisie 20 mai 2006 Cancer du rein à cellules claires métastatique (CRM) Plus de 30 % des CR deviendront métastatiques
Médicaments du futur : Tendances et enjeux. Nicolas PY, Debiopharm International forumofac.14 26/09/2014
 Médicaments du futur : Tendances et enjeux Nicolas PY, Debiopharm International forumofac.14 26/09/2014 Quelques mots sur Debiopharm Groupe fondé en 1979 Siège à Lausanne 350 collaborateurs Financièrement
Médicaments du futur : Tendances et enjeux Nicolas PY, Debiopharm International forumofac.14 26/09/2014 Quelques mots sur Debiopharm Groupe fondé en 1979 Siège à Lausanne 350 collaborateurs Financièrement
Formavie 2010. 2 Différentes versions du format PDB...3. 3 Les champs dans les fichiers PDB...4. 4 Le champ «ATOM»...5. 6 Limites du format PDB...
 Formavie 2010 Les fichiers PDB Les fichiers PDB contiennent les informations qui vont permettre à des logiciels de visualisation moléculaire (ex : RasTop ou Jmol) d afficher les molécules. Un fichier au
Formavie 2010 Les fichiers PDB Les fichiers PDB contiennent les informations qui vont permettre à des logiciels de visualisation moléculaire (ex : RasTop ou Jmol) d afficher les molécules. Un fichier au
Mise en place d une solution automatique de stockage et de visualisation de données de capture des interactions chromatiniennes à l échelle génomique
 Rapport de stage de deuxième année de DUT Génie Biologique option Bioinformatique Mise en place d une solution automatique de stockage et de visualisation de données de capture des interactions chromatiniennes
Rapport de stage de deuxième année de DUT Génie Biologique option Bioinformatique Mise en place d une solution automatique de stockage et de visualisation de données de capture des interactions chromatiniennes
Caisse Nationale de l'assurance Maladie des Travailleurs Salariés Sécurité Sociale
 Caisse Nationale de l'assurance Maladie des Travailleurs Salariés Sécurité Sociale Circulaire CNAMTS MMES et MM les Directeurs Date : - des Caisses Régionales d'assurance Maladie 27/03/96 - des Caisses
Caisse Nationale de l'assurance Maladie des Travailleurs Salariés Sécurité Sociale Circulaire CNAMTS MMES et MM les Directeurs Date : - des Caisses Régionales d'assurance Maladie 27/03/96 - des Caisses
THERANOSTIC. Pr. C. BOHUON Conférence CORATA Namur 08 juin 2011
 THERANOSTIC Pr. C. BOHUON Conférence CORATA Namur 08 juin 2011 THERANOSTIC Therapy + Diagnostic Nouveau mot reliant le choix d un médicament à la détection d une anomalie génétique (mutation ). Ex.: Cancers
THERANOSTIC Pr. C. BOHUON Conférence CORATA Namur 08 juin 2011 THERANOSTIC Therapy + Diagnostic Nouveau mot reliant le choix d un médicament à la détection d une anomalie génétique (mutation ). Ex.: Cancers
LA MITOSE CUEEP - USTL DÉPARTEMENT SCIENCES BAHIJA DELATTRE
 Biologie LA MITOSE CUEEP - USTL DÉPARTEMENT SCIENCES BAHIJA DELATTRE Février 2006 I. L'INTRODUCTION Chaque cellule d'un organisme supérieur provient de la multiplication d'une cellule préexistante (cellule
Biologie LA MITOSE CUEEP - USTL DÉPARTEMENT SCIENCES BAHIJA DELATTRE Février 2006 I. L'INTRODUCTION Chaque cellule d'un organisme supérieur provient de la multiplication d'une cellule préexistante (cellule
Les tests thyroïdiens
 Les tests thyroïdiens Mise à jour de juin 2007 par S. Fatio Réf: les images et les graphiques sont tirés de www.endotext.org (textbook d endocrinologie online et libre d accès) QuickTime et un décompresseur
Les tests thyroïdiens Mise à jour de juin 2007 par S. Fatio Réf: les images et les graphiques sont tirés de www.endotext.org (textbook d endocrinologie online et libre d accès) QuickTime et un décompresseur
Les grandes voies de transmission du signal en cancérologie : où en sommes-nous?
 Les grandes voies de transmission du signal en cancérologie : où en sommes-nous? Dynamique des réseaux de signalisation et réponse aux thérapies ciblées Pascal GAUDUCHON Unité "Biologie et Thérapies Innovantes
Les grandes voies de transmission du signal en cancérologie : où en sommes-nous? Dynamique des réseaux de signalisation et réponse aux thérapies ciblées Pascal GAUDUCHON Unité "Biologie et Thérapies Innovantes
TEST DE DÉTECTION DE LA PRODUCTION D INTERFÉRON γ POUR LE DIAGNOSTIC DES INFECTIONS TUBERCULEUSES
 TEST DE DÉTECTION DE LA PRODUCTION D INTERFÉRON γ POUR LE DIAGNOSTIC DES INFECTIONS TUBERCULEUSES Classement NABM : non inscrit code : non codé DÉCEMBRE 2006 Service évaluation des actes professionnels
TEST DE DÉTECTION DE LA PRODUCTION D INTERFÉRON γ POUR LE DIAGNOSTIC DES INFECTIONS TUBERCULEUSES Classement NABM : non inscrit code : non codé DÉCEMBRE 2006 Service évaluation des actes professionnels
Enseignement de Virologie (part. 2) Pr. Y. BAKRI Plan du cours
 Pr. Y. BAKRI Plan du cours") Université Mohammed V-Agdal Département de Biologie Faculté des Sciences Filière SVI Semestre 4 Année Universitaire : 2004-2005 Module optionnel: Biologie Humaine (M 16.1) Elément : Parasitologie - Virologie
Université Mohammed V-Agdal Département de Biologie Faculté des Sciences Filière SVI Semestre 4 Année Universitaire : 2004-2005 Module optionnel: Biologie Humaine (M 16.1) Elément : Parasitologie - Virologie
INTRODUCTION À L'ENZYMOLOGIE
 INTRODUCTION À L'ENZYMOLOGIE Les enzymes sont des macromolécules spécialisées qui - catalysent les réactions biologiques - transforment différentes formes d'énergie. Les enzymes diffèrent des catalyseurs
INTRODUCTION À L'ENZYMOLOGIE Les enzymes sont des macromolécules spécialisées qui - catalysent les réactions biologiques - transforment différentes formes d'énergie. Les enzymes diffèrent des catalyseurs
les deux premières années du Bachelor of Science en sciences pharmaceutiques
 UNIVERSITÉ DE FRIBOURG SUISSE FACULTÉ DES SCIENCES UNIVERSITÄT FREIBURG SCHWEIZ MATHEMATISCH-NATURWISSENSCHAFTLICHE FAKULTÄT Plan d'études pour les deux premières années du Bachelor of Science en sciences
UNIVERSITÉ DE FRIBOURG SUISSE FACULTÉ DES SCIENCES UNIVERSITÄT FREIBURG SCHWEIZ MATHEMATISCH-NATURWISSENSCHAFTLICHE FAKULTÄT Plan d'études pour les deux premières années du Bachelor of Science en sciences
L INSUFFISANCE CARDIAQUE
 L INSUFFISANCE CARDIAQUE EXISTE-T-IL DES TRAITEMENTS À BASE DE CELLULES SOUCHES POUR L INSUFFISANCE CARDIAQUE? Bien qu aucun traitement à base de cellules souches pour l insuffisance cardiaque n ait encore
L INSUFFISANCE CARDIAQUE EXISTE-T-IL DES TRAITEMENTS À BASE DE CELLULES SOUCHES POUR L INSUFFISANCE CARDIAQUE? Bien qu aucun traitement à base de cellules souches pour l insuffisance cardiaque n ait encore
Respiration Mitochondriale
 Université Pierre et Marie Curie Respiration Mitochondriale Objectifs au cours de Révisions Biochimie PCEM2 Révisions Biochimie Métabolique 2004-2005 Pr. A. Raisonnier (alain.raisonnier@upmc.fr) Mise à
Université Pierre et Marie Curie Respiration Mitochondriale Objectifs au cours de Révisions Biochimie PCEM2 Révisions Biochimie Métabolique 2004-2005 Pr. A. Raisonnier (alain.raisonnier@upmc.fr) Mise à
MASTER (LMD) PARCOURS MICROORGANISMES, HÔTES, ENVIRONNEMENTS (MHE)
 PARCOURS MICROORGANISMES, HÔTES, ENVIRONNEMENTS (MHE)") MASTER (LMD) PARCOURS MICROORGANISMES, HÔTES, ENVIRONNEMENTS (MHE) RÉSUMÉ DE LA FORMATION Type de diplôme : Master (LMD) Domaine ministériel : Sciences, Technologies, Santé Mention : BIOLOGIE DES PLANTES
MASTER (LMD) PARCOURS MICROORGANISMES, HÔTES, ENVIRONNEMENTS (MHE) RÉSUMÉ DE LA FORMATION Type de diplôme : Master (LMD) Domaine ministériel : Sciences, Technologies, Santé Mention : BIOLOGIE DES PLANTES
Comité conseil en matière de prévention et sécurité des personnes et des biens Octobre 2013
 PROJET Encadrement du système de caméras de sécurité et Foire aux questions Comité conseil en matière de prévention et sécurité des personnes et des biens Octobre 2013 2 3 Table des matières 1. CADRE JURIDIQUE...4
PROJET Encadrement du système de caméras de sécurité et Foire aux questions Comité conseil en matière de prévention et sécurité des personnes et des biens Octobre 2013 2 3 Table des matières 1. CADRE JURIDIQUE...4
La Greffe de Cellules Souches Hématopoïétiques
 La Greffe de Cellules Souches Hématopoïétiques Professeur Ibrahim Yakoub-Agha CHRU de LILLE (Illustration de J. Cloup, extraite du CD-Rom «greffe de Moelle» réalisé par la société K Noë) La moelle osseuse
La Greffe de Cellules Souches Hématopoïétiques Professeur Ibrahim Yakoub-Agha CHRU de LILLE (Illustration de J. Cloup, extraite du CD-Rom «greffe de Moelle» réalisé par la société K Noë) La moelle osseuse
Le programme de soutien des soins primaires pour diabétiques de London : Des soins pour diabétiques différents
 Le programme de soutien des soins primaires pour diabétiques de London : Des soins pour diabétiques différents S U C C E S S S T O R Y 1 Diabetes Care with a Difference: The London Primary Care Diabetes
Le programme de soutien des soins primaires pour diabétiques de London : Des soins pour diabétiques différents S U C C E S S S T O R Y 1 Diabetes Care with a Difference: The London Primary Care Diabetes
Information génétique
 chapitre 3 Information génétique et division cellulaire L étude de la division cellulaire est abordée pour découvrir comment est transmise et conservée l information génétique portée par les chromosomes.
chapitre 3 Information génétique et division cellulaire L étude de la division cellulaire est abordée pour découvrir comment est transmise et conservée l information génétique portée par les chromosomes.
Physiopathologie : de la Molécule à l'homme
 Mention Sciences du Vivant Spécialité de Master : Physiopathologie : de la Molécule à l'homme Pourquoi, comment, combien, contourner, greffer, restaurer, inhiber, suppléer Responsables : Dr Gilles Prévost
Mention Sciences du Vivant Spécialité de Master : Physiopathologie : de la Molécule à l'homme Pourquoi, comment, combien, contourner, greffer, restaurer, inhiber, suppléer Responsables : Dr Gilles Prévost
LIGNES DIRECTRICES CLINIQUES TOUT AU LONG DU CONTINUUM DE SOINS : Objectif de ce chapitre. 6.1 Introduction 86
 LIGNES DIRECTRICES CLINIQUES TOUT AU LONG DU CONTINUUM DE SOINS : ÉTABLISSEMENT DE LIENS ENTRE LES PERSONNES CHEZ QUI UN DIAGNOSTIC D INFECTION À VIH A ÉTÉ POSÉ ET LES SERVICES DE SOINS ET DE TRAITEMENT
LIGNES DIRECTRICES CLINIQUES TOUT AU LONG DU CONTINUUM DE SOINS : ÉTABLISSEMENT DE LIENS ENTRE LES PERSONNES CHEZ QUI UN DIAGNOSTIC D INFECTION À VIH A ÉTÉ POSÉ ET LES SERVICES DE SOINS ET DE TRAITEMENT
La filtration glomérulaire et sa régulation
 UE3-2 - Physiologie rénale Chapitre 4 : La filtration glomérulaire et sa régulation Professeur Diane GODIN-RIBUOT Année universitaire 2010/2011 Université Joseph Fourier de Grenoble - Tous droits réservés.
UE3-2 - Physiologie rénale Chapitre 4 : La filtration glomérulaire et sa régulation Professeur Diane GODIN-RIBUOT Année universitaire 2010/2011 Université Joseph Fourier de Grenoble - Tous droits réservés.
Saadi KHOCHBIN. Directeur de recherche CNRS. médaille d Argent. Institut des sciences biologiques. Institut Albert Bonniot (IAB)
") Saadi KHOCHBIN Directeur de recherche CNRS Institut Albert Bonniot (IAB) Inserm / UJF Institut des sciences biologiques médaille d Argent Les projets développés par Saadi Khochbin s inscrivent essentiellement
Saadi KHOCHBIN Directeur de recherche CNRS Institut Albert Bonniot (IAB) Inserm / UJF Institut des sciences biologiques médaille d Argent Les projets développés par Saadi Khochbin s inscrivent essentiellement
Gènes du développement et structuration de l organisme
 03-Gènes du développement_00/maquette imports 19/01/10 10:45 Page99 Revue des Questions scientifiques, 2010, 181 (1) : 99-110 Gènes du développement et structuration de l organisme René Rezsohazy Unité
03-Gènes du développement_00/maquette imports 19/01/10 10:45 Page99 Revue des Questions scientifiques, 2010, 181 (1) : 99-110 Gènes du développement et structuration de l organisme René Rezsohazy Unité
UE : GENE 2208. Responsable : Enseignant : ECUE 1. Enseignant : ECUE 2. Dr COULIBALY Foungotin Hamidou
 UE : GENE 2208 Responsable : Enseignant : ECUE 1 Enseignant : ECUE 2 Dr COULIBALY Foungotin Hamidou Spécialités : Génétique Humaine Biologie de la Procréation Cytogénétique Spermiologie Essais Cliniques
UE : GENE 2208 Responsable : Enseignant : ECUE 1 Enseignant : ECUE 2 Dr COULIBALY Foungotin Hamidou Spécialités : Génétique Humaine Biologie de la Procréation Cytogénétique Spermiologie Essais Cliniques
- 2 - faire industriel dans la mise au point des produits biologiques. L Institut Roche de Recherche et Médecine Translationnelle (IRRMT, basé à
 Information presse Roche, l Agence Nationale de Recherches sur le Sida et les hépatites virales (ANRS), le Baylor Research Institute (BRI) et Inserm Transfert mettent en place une coopération stratégique
Information presse Roche, l Agence Nationale de Recherches sur le Sida et les hépatites virales (ANRS), le Baylor Research Institute (BRI) et Inserm Transfert mettent en place une coopération stratégique
EXERCICES : MECANISMES DE L IMMUNITE : pages 406 407 408 409 410
 EXERCICES : MECANISMES DE L IMMUNITE : pages 406 407 408 409 410 EXERCICE 1 PAGE 406 : EXPERIENCES A INTERPRETER Question : rôles respectifs du thymus et de la moelle osseuse dans la production des lymphocytes.
EXERCICES : MECANISMES DE L IMMUNITE : pages 406 407 408 409 410 EXERCICE 1 PAGE 406 : EXPERIENCES A INTERPRETER Question : rôles respectifs du thymus et de la moelle osseuse dans la production des lymphocytes.
3: Clonage d un gène dans un plasmide
 3: Clonage d un gène dans un plasmide Le clonage moléculaire est une des bases du génie génétique. Il consiste à insérer un fragment d'adn (dénommé insert) dans un vecteur approprié comme un plasmide par
3: Clonage d un gène dans un plasmide Le clonage moléculaire est une des bases du génie génétique. Il consiste à insérer un fragment d'adn (dénommé insert) dans un vecteur approprié comme un plasmide par
Plateforme Transgenèse/Zootechnie/Exploration Fonctionnelle IBiSA. «Anexplo» Service Transgenèse. Catalogue des prestations
 Plateforme Transgenèse/Zootechnie/Exploration Fonctionnelle IBiSA «Anexplo» Service Transgenèse Catalogue des prestations 04/01/12 - Page 1 sur 8 Présentation du service de Transgenèse Le service de Transgenèse
Plateforme Transgenèse/Zootechnie/Exploration Fonctionnelle IBiSA «Anexplo» Service Transgenèse Catalogue des prestations 04/01/12 - Page 1 sur 8 Présentation du service de Transgenèse Le service de Transgenèse
Direction des bibliothèques. Sondage Ithaka S+R. Questionnaire français Université de Montréal
 Direction des bibliothèques Sondage Ithaka S+R Questionnaire français Université de Montréal Décembre 2014 Modules retenus Découverte Activités de recherche numérique Compétences informationnelles des
Direction des bibliothèques Sondage Ithaka S+R Questionnaire français Université de Montréal Décembre 2014 Modules retenus Découverte Activités de recherche numérique Compétences informationnelles des
Rôle des acides biliaires dans la régulation de l homéostasie du glucose : implication de FXR dans la cellule bêta-pancréatique
 Rôle des acides biliaires dans la régulation de l homéostasie du glucose : implication de FXR dans la cellule bêta-pancréatique Tuteur : Anne Muhr-Tailleux cardiovasculaires et diabète (Equipe 1) Institut
Rôle des acides biliaires dans la régulation de l homéostasie du glucose : implication de FXR dans la cellule bêta-pancréatique Tuteur : Anne Muhr-Tailleux cardiovasculaires et diabète (Equipe 1) Institut
Master de Bioinformatique et Biologie des Systèmes Toulouse http://m2pbioinfo.biotoul.fr Responsable : Pr. Gwennaele Fichant
 Master de Bioinformatique et Biologie des Systèmes Toulouse http://m2pbioinfo.biotoul.fr Responsable : Pr. Gwennaele Fichant Parcours: Master 1 : Bioinformatique et biologie des Systèmes dans le Master
Master de Bioinformatique et Biologie des Systèmes Toulouse http://m2pbioinfo.biotoul.fr Responsable : Pr. Gwennaele Fichant Parcours: Master 1 : Bioinformatique et biologie des Systèmes dans le Master
ANTIVIRAUX ET INTERFERONS LES ANTIVIRAUX
 ANTIVIRAUX ET INTERFERONS LES ANTIVIRAUX La thérapeutique antivirale progresse avec lenteur car elle s'attaque à des micro-organismes ne se multipliant qu'à l'intérieur des cellules vivantes dont ils détournent
ANTIVIRAUX ET INTERFERONS LES ANTIVIRAUX La thérapeutique antivirale progresse avec lenteur car elle s'attaque à des micro-organismes ne se multipliant qu'à l'intérieur des cellules vivantes dont ils détournent
Les tests génétiques à des fins médicales
 Les tests génétiques à des fins médicales Les tests génétiques à des fins médicales Nous avons tous hérité d une combinaison unique de gènes de la part de nos parents. Cette constitution originale et l
Les tests génétiques à des fins médicales Les tests génétiques à des fins médicales Nous avons tous hérité d une combinaison unique de gènes de la part de nos parents. Cette constitution originale et l
ECOLE PRATIQUE DES HAUTES ETUDES Sciences de la Vie et de la Terre
 MINISTERE DE L EDUCATION NATIONALE ECOLE PRATIQUE DES HAUTES ETUDES Sciences de la Vie et de la Terre MEMOIRE Présenté par LE PAVEC GWENAELLE Pour l obtention du diplôme de l Ecole Pratique des Hautes
MINISTERE DE L EDUCATION NATIONALE ECOLE PRATIQUE DES HAUTES ETUDES Sciences de la Vie et de la Terre MEMOIRE Présenté par LE PAVEC GWENAELLE Pour l obtention du diplôme de l Ecole Pratique des Hautes
Section «Maturité fédérale» EXAMENS D'ADMISSION Session de février 2014 RÉCAPITULATIFS DES MATIÈRES EXAMINÉES. Formation visée
 EXAMENS D'ADMISSION Admission RÉCAPITULATIFS DES MATIÈRES EXAMINÉES MATIÈRES Préparation en 3 ou 4 semestres Formation visée Préparation complète en 1 an 2 ème partiel (semestriel) Niveau Durée de l examen
EXAMENS D'ADMISSION Admission RÉCAPITULATIFS DES MATIÈRES EXAMINÉES MATIÈRES Préparation en 3 ou 4 semestres Formation visée Préparation complète en 1 an 2 ème partiel (semestriel) Niveau Durée de l examen
Les bases de données transcriptionnelles en ligne
 Les bases de données transcriptionnelles en ligne Différents concepts en régulation transcriptionnelle sites de fixation - in vitro/vivo? - quelle technique? - degré de confiance? facteur de transcription
Les bases de données transcriptionnelles en ligne Différents concepts en régulation transcriptionnelle sites de fixation - in vitro/vivo? - quelle technique? - degré de confiance? facteur de transcription
EVALUATION DES TECHNOLOGIES DE SANTÉ ANALYSE MÉDICO-ÉCONOMIQUE. Efficacité et efficience des hypolipémiants Une analyse centrée sur les statines
 EVALUATION DES TECHNOLOGIES DE SANTÉ ANALYSE MÉDICO-ÉCONOMIQUE Efficacité et efficience des hypolipémiants Une analyse centrée sur les statines Juillet 2010 Mise à jour Septembre 2010 1 Le rapport complet
EVALUATION DES TECHNOLOGIES DE SANTÉ ANALYSE MÉDICO-ÉCONOMIQUE Efficacité et efficience des hypolipémiants Une analyse centrée sur les statines Juillet 2010 Mise à jour Septembre 2010 1 Le rapport complet
MAB Solut. vos projets. MABLife Génopole Campus 1 5 rue Henri Desbruères 91030 Evry Cedex. www.mabsolut.com. intervient à chaque étape de
 Mabsolut-DEF-HI:Mise en page 1 17/11/11 17:45 Page1 le département prestataire de services de MABLife de la conception à la validation MAB Solut intervient à chaque étape de vos projets Création d anticorps
Mabsolut-DEF-HI:Mise en page 1 17/11/11 17:45 Page1 le département prestataire de services de MABLife de la conception à la validation MAB Solut intervient à chaque étape de vos projets Création d anticorps
Transgene accorde une option de licence exclusive pour le développement et la commercialisation de son produit d immunothérapie TG4010
 Parc d Innovation d Illkirch, France, le 10 mars 2010 Transgene accorde une option de licence exclusive pour le développement et la commercialisation de son produit d immunothérapie TG4010 Transgene (Euronext
Parc d Innovation d Illkirch, France, le 10 mars 2010 Transgene accorde une option de licence exclusive pour le développement et la commercialisation de son produit d immunothérapie TG4010 Transgene (Euronext
L immunoenzymologie. Technique puissante couramment utilisée e en recherche et en diagnostic cificité des anticorps pour leurs nes
 L immunoenzymologie Technique puissante couramment utilisée e en recherche et en diagnostic Basée e sur la très s grande spécificit cificité des anticorps pour leurs antigènes nes Test qualitatif Détection
L immunoenzymologie Technique puissante couramment utilisée e en recherche et en diagnostic Basée e sur la très s grande spécificit cificité des anticorps pour leurs antigènes nes Test qualitatif Détection
DIAPOSITIVE 1 Cette présentation a trait à la réglementation sur les thérapies cellulaires.
 Produits de thérapie cellulaire DIAPOSITIVE 1 Cette présentation a trait à la réglementation sur les thérapies cellulaires. DIAPOSITIVE 2 La fabrication des thérapies cellulaires est examinée par la Division
Produits de thérapie cellulaire DIAPOSITIVE 1 Cette présentation a trait à la réglementation sur les thérapies cellulaires. DIAPOSITIVE 2 La fabrication des thérapies cellulaires est examinée par la Division
ANTICORPS POLYCLONAUX ANTI IMMUNOGLOBULINES
 L OUTIL IDEAL POUR TOUTES LES DETECTIONS IMMUNOCHIMIQUES pour toutes les techniques immunodosages (EIA/ELISA) dot/ westernblot immunohistochimie immunocytochimie cytométrie en flux quel que soit le système
L OUTIL IDEAL POUR TOUTES LES DETECTIONS IMMUNOCHIMIQUES pour toutes les techniques immunodosages (EIA/ELISA) dot/ westernblot immunohistochimie immunocytochimie cytométrie en flux quel que soit le système
La reconnaissance moléculaire: la base du design rationnel Modélisation moléculaire: Introduction Hiver 2006
 La reconnaissance moléculaire: la base du design rationnel En 1890 Emil Fisher a proposé le modèle "serrure et clé" pour expliquer la façon de fonctionner des systèmes biologiques. Un substrat rentre et
La reconnaissance moléculaire: la base du design rationnel En 1890 Emil Fisher a proposé le modèle "serrure et clé" pour expliquer la façon de fonctionner des systèmes biologiques. Un substrat rentre et
Innovations thérapeutiques en transplantation
 Innovations thérapeutiques en transplantation 3èmes Assises de transplantation pulmonaire de la région Est Le 16 octobre 2010 Dr Armelle Schuller CHU Strasbourg Etat des lieux en transplantation : 2010
Innovations thérapeutiques en transplantation 3èmes Assises de transplantation pulmonaire de la région Est Le 16 octobre 2010 Dr Armelle Schuller CHU Strasbourg Etat des lieux en transplantation : 2010
Université Louis Pasteur Strasbourg I Ecole Doctorale des Sciences de la Vie et de la Santé THESE. En vue de l obtention du grade de
 Université Louis Pasteur Strasbourg I Ecole Doctorale des Sciences de la Vie et de la Santé THESE En vue de l obtention du grade de Docteur de l Université Louis Pasteur de Strasbourg Discipline : Aspects
Université Louis Pasteur Strasbourg I Ecole Doctorale des Sciences de la Vie et de la Santé THESE En vue de l obtention du grade de Docteur de l Université Louis Pasteur de Strasbourg Discipline : Aspects
Considérations sur la crise et le marché intérieur 1
 Considérations sur la crise et le marché intérieur 1 Auteurs : Jacques Potdevin : Président de la Fédération des Experts comptables Européens (FEE - Federation of European Accountants 2 ) de 2007 à 2008.
Considérations sur la crise et le marché intérieur 1 Auteurs : Jacques Potdevin : Président de la Fédération des Experts comptables Européens (FEE - Federation of European Accountants 2 ) de 2007 à 2008.
ORIGINE ET DIFFERENTS TYPES DE RYTHMES BIOLOGIQUES. Carine Bécamel carine.becamel@igf.cnrs.fr 04 67 14 29 83
 ORIGINE ET DIFFERENTS TYPES DE RYTHMES BIOLOGIQUES Carine Bécamel carine.becamel@igf.cnrs.fr 04 67 14 29 83 INTRODUCTION Rythmes biologiques: définition Tout être vivant (humain, animal, végétal) est soumis
ORIGINE ET DIFFERENTS TYPES DE RYTHMES BIOLOGIQUES Carine Bécamel carine.becamel@igf.cnrs.fr 04 67 14 29 83 INTRODUCTION Rythmes biologiques: définition Tout être vivant (humain, animal, végétal) est soumis
Enseignement de Virologie (part. 2) Pr. Y. BAKRI Plan du cours
 Pr. Y. BAKRI Plan du cours") Université Mohammed V-Agdal Département de Biologie Faculté des Sciences Filière SVI Semestre 4 Année Universitaire : 2009-2010 Module optionnel: Biologie Humaine (M 16.1) Elément : Parasitologie - Virologie
Université Mohammed V-Agdal Département de Biologie Faculté des Sciences Filière SVI Semestre 4 Année Universitaire : 2009-2010 Module optionnel: Biologie Humaine (M 16.1) Elément : Parasitologie - Virologie
TEST ELISA (ENZYME-LINKED IMMUNOSORBENT ASSEY)
") TEST ELISA (ENZYME-LINKED IMMUNOSORBENT ASSEY) Lise Vézina, technicienne de laboratoire Michel Lacroix, agronome-phytopathologiste Direction de l innovation scientifique et technologique Au Laboratoire
TEST ELISA (ENZYME-LINKED IMMUNOSORBENT ASSEY) Lise Vézina, technicienne de laboratoire Michel Lacroix, agronome-phytopathologiste Direction de l innovation scientifique et technologique Au Laboratoire
LA GESTION DES RESSOURCES HUMAINES Anne DIETRICH Frédérique PIGEYRE 2005, repères, La découverte
 LA GESTION DES RESSOURCES HUMAINES Anne DIETRICH Frédérique PIGEYRE 2005, repères, La découverte La GRH constitue une préoccupation permanente de toute entreprise, de tout dirigeant, qu il s agisse de
LA GESTION DES RESSOURCES HUMAINES Anne DIETRICH Frédérique PIGEYRE 2005, repères, La découverte La GRH constitue une préoccupation permanente de toute entreprise, de tout dirigeant, qu il s agisse de
Détection des duplications en tandem au niveau nucléique à l'aide de la théorie des flots
 Université Toulouse 3 Paul Sabatier(UT3 Paul Sabatier) Informatique Spécialité Bioinformatique Eric AUDEMARD lundi 28 novembre 2011 Détection des duplications en tandem au niveau nucléique à l'aide de
Université Toulouse 3 Paul Sabatier(UT3 Paul Sabatier) Informatique Spécialité Bioinformatique Eric AUDEMARD lundi 28 novembre 2011 Détection des duplications en tandem au niveau nucléique à l'aide de
Vue d ensemble : Office of Cellular, Tissue and Gene Therapies
 Vue d ensemble : Office of Cellular, Tissue and Gene Therapies DIAPOSITIVE 1 Cette présentation fournit une vue d ensemble de l Office of Cellular, Tissue, and Gene Therapies (bureau des thérapies cellulaires,
Vue d ensemble : Office of Cellular, Tissue and Gene Therapies DIAPOSITIVE 1 Cette présentation fournit une vue d ensemble de l Office of Cellular, Tissue, and Gene Therapies (bureau des thérapies cellulaires,
La PCR en temps réel: principes et applications
 Reviews in Biology and Biotechnology By The Moroccan Society of Biology in Canada Vol.2, No 2, December 2002. pp.2-11 Printed in Canada La PCR en temps réel: principes et applications Elyse Poitras et
Reviews in Biology and Biotechnology By The Moroccan Society of Biology in Canada Vol.2, No 2, December 2002. pp.2-11 Printed in Canada La PCR en temps réel: principes et applications Elyse Poitras et
Critères pour les méthodes de quantification des résidus potentiellement allergéniques de protéines de collage dans le vin (OIV-Oeno 427-2010)
") Méthode OIV- -MA-AS315-23 Type de méthode : critères Critères pour les méthodes de quantification des résidus potentiellement allergéniques de protéines de collage (OIV-Oeno 427-2010) 1 Définitions des
Méthode OIV- -MA-AS315-23 Type de méthode : critères Critères pour les méthodes de quantification des résidus potentiellement allergéniques de protéines de collage (OIV-Oeno 427-2010) 1 Définitions des
Le grand écart de la recherche fondamentale vers l application biomédicale
 Le grand écart de la recherche fondamentale vers l application biomédicale Ludger Johannes «Chemical Biology of Membranes and Therapeutic Delivery» unit U1143 INSERM UMR3666 CNRS «Endocytic Trafficking
Le grand écart de la recherche fondamentale vers l application biomédicale Ludger Johannes «Chemical Biology of Membranes and Therapeutic Delivery» unit U1143 INSERM UMR3666 CNRS «Endocytic Trafficking