Thesis. Reference. Rôle et régulation de MD-2, la protéine réceptrice de l'endotoxine bactérienne. TISSIÈRES, Pierre
|
|
|
- Frédéric Gamache
- il y a 8 ans
- Total affichages :
Transcription
1 Thesis Rôle et régulation de MD-2, la protéine réceptrice de l'endotoxine bactérienne TISSIÈRES, Pierre Abstract Notre travail de recherche a pour thématique l'étude du rôle de MD-2 et la régulation de son expression. MD-2, une glycoprotéine intimement liée à TLR4, joue un rôle primordial dans la reconnaissance par l'hôte du lipopolysaccharide (LPS)le principal constituant de la paroi externe des bactéries à Gram négatif. Dans la première partie, nous avons pu démontrer que la forme soluble de MD-2 est une protéine de la phase aiguë, une opsonine qui lie les bactéries à Gram négatif et facilite leur phagocytose. Cette découverte participe au concept que l'inflammation aiguë induite par les bactéries est essentielle pour la résolution d'infections à bactéries à Gram négatif. Dans la deuxième partie, nous avons étudié la régulation de l'expression du gène MD-2 lors de l'induction d'une réponse de phase aiguë, et démontré le rôle central de l'interaction entre deux facteurs de transcription : PU.1 ET C/EBPB. Reference TISSIÈRES, Pierre. Rôle et régulation de MD-2, la protéine réceptrice de l'endotoxine bactérienne. Thèse de doctorat : Univ. Genève, 2009, no. Sc URN : urn:nbn:ch:unige Available at: Disclaimer: layout of this document may differ from the published version. [ Downloaded 29/06/2016 at 15:07:25 ]
2 UNIVERSITE DE GENEVE Département de Biologie Moléculaire FACULTE DES SCIENCES Professeur Ulrich Schibler Département d Anesthésiologie, Pharmacologie et Soins Intensifs FACULTE DE MEDECINE Professeur Jérôme Pugin Rôle et régulation de MD-2, la protéine réceptrice de l endotoxine bactérienne THESE présentée à la Faculté des Sciences de l Université de Genève pour obtenir le grade de Docteur ès Sciences, mention biologie par Pierre TISSIERES d Orsières, Valais Thèse n 4120 Genève ReproMail 2009
3 LISTE DES ABBREVIATIONS Akt Protein kinase B AP-1 Activated protein 1 AND/DNA Acide deoxyribo-nucléique ARN/RNA Acide ribo-nucléique CD14 Cluster of differentiation 14 C/EBP CAAT/enhancer binding protein ChIP Chromatin immunoprecipitation ERK Extracellular signal regulated kinase HMGB1 High mobility group box-1 IFN Interféron IKK Inhibitor of Nuclear factor-kappab kinase IRAK Interleukin-1 receptor-associated kinase IRF3 Interferon regulatory factor-3 JNK c-jun N-terminal protein kinase LBP LPS-binding protein LPS Lipopolysaccharide, endotoxine LRR Leucine-rich repeat MAPK Mitogen-activated protein kinase MD-2 Myeloid differentiation factor-2 smd-2 Soluble MD-2 MyD88 Myeloid differentiation factor 88 NBS-LRR Nucleotide binding-site leucine-rich repeats NFκB Nuclear factor-kappab NOD Nucleotide-binding oligomerization domain PCR Polymerase chain reaction qpcr Quantitative polymerase chain reaction RAGE Receptor for advanced glycation end-product sirna Short-interfering RNA TAK1 Transforming growth factor β activated kinase 1 TIRAP Toll/IL-1R domain-containing adaptor protein TLR Toll-like receptors TNF Tumor necrosis factor TRAF6 TNF receptor-associated factor 6 TRAM TRIF-related adaptor molecule TRIF TIR-containing adaptor-inducing IFNβ 2
4 TABLE DES MATIERES I. REMERCIEMENTS... 4 II. RESUME... 5 III. INTRODUCTION... 6 III.1 Infection systémique et susceptibilité de l hôte aux infections... 6 III.2 Immunité Innée... 8 III.3 Récepteurs Toll-like III.4 Structure et fonction de MD III.5 Reconnaissance des bactéries à Gram négatif par TLR4:MD IV. RESULTATS IV.1 MD-2 est une protéine de phase aiguë et une opsonine pour les bactéries à Gram négatif IV.1.1 Article 1: MD-2 is an acute phase protein and an opsonin for Gramnegative bacteria IV.1.2 Article 2: Role of MD-2 in the opsonophagocytosis of Gram-negative bacteria IV.2 Article 3: Cooperation between PU.1 and CAAT/Enhancer binding protein β is necessary to induce the expression of MD-2 gene V. DISCUSSIONS ET PERSPECTIVES V.1 Est-ce qu un déficit en MD-2 altère la réponse de l hôte aux infections Gram- négatives? V.2 Quel sont les mécanismes de régulation de l expression de MD-2? V.3 Réponse de l épithélium respiratoire lors du sepsis VI. CONCLUSION VII. REFERENCES
5 I. REMERCIEMENTS Pour conclure ces cinq années de thèse, je souhaiterai remercier tous ceux qui y ont participé directement ou indirectement. Je remercie Séverine, mon épouse, et mes enfants Arthur et Charlotte, pour leur patience, et leur soutien. Je remercie Jérôme Pugin pour m avoir fait confiance, et m avoir tant appris, professionnellement et scientifiquement, me préparant comme personne n aurait pu le faire à ma future carrière. Je remercie mes amis et mes collègues de laboratoire : Tanguy, Pierre-Emmanuel, Laurent, Vincent, Julien, Alex, Stephan, Irène, Thierry, Raphaël, Agnieszka, Geneviève, Angela, Evelyne, Clara, Vandack, Isabelle, Rachel, et Pascale. Je remercie, le Professeur Schibler d avoir accepté de co-diriger ma thèse. Je remercie la Professeure Régine Landmann d avoir accepté de participer à mon Jury de Thèse, ainsi que le Docteur Thierry Roger, qui a suivit le cheminement de ma thèse dès le début, pour ces nombreux conseils, et pour avoir accepté de participer à mon Jury. Je remercie Denis Devictor, mon mentor, qui depuis 15 ans m accompagne dans ma carrière. Je remercie Michel Berner et Maurice Beghetti pour leur soutien durant ma formation. Je remercie Patrick Linder et Dominique Belin au contact desquels j ai développé mon goût pour la biologie fondamentale. Je remercie aussi Peter Frey et Jacques-Antoine Haefliger qui ont dirigé mes premiers pas en recherche et m ont transmis leur enthousiasme. Je remercie la Faculté de Médecine de Genève, la Roche Research Foundation, la Novartis Consumer Health Foundation, la Novartis Foundation, la Fondation Boninchi, la Fondation Gertrud von Meissner pour leur soutien financier. Je tiens à rendre hommage à mes oncles, Paul et Alfred, qui par leur exemple ont orienté mon parcours vers la biologie. Pour conclure, je remercie mes parents, pour leur soutien indéfectible, durant ces nombreuses années d études. 4
6 II. RESUME Notre travail de recherche a pour thématique l étude du rôle de MD-2 et la régulation de son expression. MD-2, une glycoprotéine intimement liée à TLR4, joue un rôle primordial dans la reconnaissance par l hôte du lipopolysaccharide (LPS) le principal constituant de la paroi externe des bactéries à Gram négatif. Les travaux présentés dans cette thèse ont été effectués durant les 5 années passées dans le laboratoire du Professeur Jérôme Pugin au sein du Département de Microbiologie et Médecine Moléculaire de la Faculté de Médecine de l Université de Genève. Dans la première partie, nous avons pu démontrer que la forme soluble de MD-2 est une protéine de la phase aiguë, une opsonine qui lie les bactéries à Gram négatif et facilite leur phagocytose. (Tissières et al. Blood 2008) 1. Ce travail a été par la suite confirmé par d autres auteurs. 2 Cette découverte participe au concept que l inflammation aiguë induite par les bactéries est essentielle pour la résolution d infections à bactéries à Gram négatif (Tissières et Pugin, Curr Opin Infect Dis 2009) 3. Dans la deuxième partie, nous avons étudié la régulation de l expression du gène MD-2 lors de l induction d une réponse de phase aiguë, c'est-à-dire médiée par l IL-6, et montré que : 1) cette régulation nécessitait la coopération de 2 facteurs de transcription : C/EBPβ et PU.1 et 2) un promoteur alternatif régulait l expression de MD-2 chez l homme et la souris. Outre l intérêt en terme de mécanisme de régulation transcriptionnelle impliquant la coopération de deux facteurs permettant le recrutement de la machine de transcription («recrutement non-tata»), et l identification de promoteurs spécifiques aux lignées myéloïdes et non-myéloïdes, ce travail a permis de mettre en évidence pour la première fois le rôle central du facteur de transcription PU.1 dans une lignée nonmyéloïde, l hépatocyte. PU.1 a initialement été reconnu comme un facteur de transcription essentiel dans le mécanisme de différenciation des lignées myélo-monocytaires. Nous avons démontré que PU.1 est un facteur de transcription essentiel dans l induction de l expression de phase aiguë de MD- 2. (Tissières et al. J Biol Chem 2009) 4. Ces travaux sont introduits par une description détaillée des mécanismes innés de reconnaissance de pathogènes par l hôte, de la description de la famille des récepteurs Toll-like, et plus spécifiquement du complexe-récepteur de reconnaissance du LPS et des bactéries à Gram négatif. Nous concluons sur une mise en perspective de nos travaux, principalement axée sur les mécanismes de régulation de l expression de MD-2 et son importance fonctionnelle dans diverses situations physiopathologiques impliquant la réponse de l hôte lors d infection à bactéries à Gram négatif. 5
7 III. INTRODUCTION III.1 Infection systémique et susceptibilité de l hôte aux infections Le sepsis sévère et le choc septique sont les formes les plus sévères d infection bactérienne chez le mammifère et l homme en particulier. On compte plus de cas chaque année aux Etats-Unis, correspondant à 2-11% de l ensemble des hospitalisations. 5-7 Le sepsis sévère et le choc septique constituent la principale cause de mortalité en réanimation pouvant atteindre 40 à 70%. 6 Cette mortalité est d autant plus importante que l infection est associée à une dysfonction d organes, survient chez un hôte immuno-compromis, âgé, et/ou ayant subi une chirurgie à haut risque. 8 La réponse de l hôte à une infection implique de nombreux acteurs : bactéries, facteurs liés à l hôte, facteurs liés aux soins (antibiothérapie, iatrogénicité). La pathogénicité bactérienne inclus de multiples aspects allant des facteurs de virulence, des résistances aux antibiotiques, de la charge bactérienne, et du site d infection. Les facteurs liés à l hôte peuvent être catégorisés en cinq groupes que nous allons décrire. Une composante génétique influence la réponse de l hôte. Le travail de Sorensen et al. publié en 1988 en est une brillante illustration. 9 Dans ce travail, intitulé «Influences génétiques et environnementales influençant la mort prématurée d adulte», l évaluation du risque relatif (RR) de décès avant l âge de 50 ans, chez 960 sujets ayant été adoptés étant enfants, d une cause identique à celle de leur parents biologiques. Ce travail a mis en évidence que la première cause de décès liée à un facteur familial sont les infections (RR 5.81, 95%CI ), avec un risque relatif plus élevé que les causes cardiovasculaires (RR 4.52, 95%CI : ), ou les cancers (RR 1.19 ; ). Ce travail démontre l existence d une susceptibilité génétique aux infections et notamment aux infections graves mortelles. 9 Au même titre, l origine ethnique peut, dans certains cas, influencer la réponse de l hôte aux bactéries. L un des exemples est celui des Eskimos, ou des Apaches des «White Mountains» qui présentent un risque de développer une infection sévère à pneumocoque huit fois plus élevé que le reste de la population des Etats-Unis. 10,11 Au même titre, l incidence, sur la base d étude de population («population-based incidence») dans une très grande cohorte (> 71 millions de patients admis dans 68 hôpitaux aux Etats-Unis), a identifié un risque accru de sepsis, ajusté aux conditions socio-économiques, dans la population afro-américaine par rapport à la population caucasienne ou hispanique (respectivement, 6.08/1000 ; 4.06/1000 et 3.58/1000). 12. La susceptibilité génétique aux infections peut être théoriquement due à une modification du niveau d expression, ou d expression «tronquée» de protéines impliquées dans la reconnaissance des bactéries, la signalisation cellulaire, la réponse inflammatoire ou la clearance bactérienne. 13 La liste des protéines pour lesquelles des mutations altèrent la réponse de l hôte en le rendant susceptible aux infections et au développement de sepsis est longue. Nous n en évoquerons que certaines afin d illustrer les différents niveaux d atteinte dans la réponse de l hôte. Schématiquement, nous pouvons considérer que la réponse de l hôte aux bactéries nécessite les étapes suivantes : reconnaissance de 6
8 la présence d un agent pathogène «non-soi», signalisation et induction d une réponse inflammatoire. A chacun de ces niveaux, des mutations de la protéine elle-même ou d éléments de contrôle de son expression peuvent affecter cette réponse. Par exemple, des mutations touchant les récepteurs de l immunité innée anti-microbienne TLR1, TLR2, TLR3 et TLR4 ont été largement décrites dans de nombreuse situations physiopathologiques et nous en reparlerons plus loin L atteinte de protéines essentielles dans la transduction de l un des signaux induits par les TLRs, telles que les mutations d Interleukin-1 receptor-associated kinase (IRAK) -1 et -4 sont aussi clairement associées à une propension à développer des infections bactériennes récurrentes, ainsi qu un pronostic plus sombre en cas de sepsis bactérien. 17,18 Bien que l âge de l hôte soit communément reconnu comme un élément (dépendant de nombreux cofacteurs) influençant la réponse immunitaire aux infections, le sexe l est moins. Une étude récente évaluant une cohorte de 1608 patients, dont 608 femmes, a pourtant clairement montré que le risque de décès durant leur séjour à l hôpital d un sepsis sévère était inférieur chez les femmes que chez les hommes (OR 0.69, 95%CI ). 19 Ce travail, confirmé dans une étude à moindre effectifs, suggère une différence de mortalité - dans des cas de sepsis chirurgicaux - d environ 30% en faveur des femmes. 20 Bien que le status hormonal puisse jouer un rôle, la relative protection du sexe féminin reste pour l heure mystérieuse. L état immunologique de l hôte (inflammation chronique, immunosuppression), au même titre que différentes co-morbidités tels que le diabète ou des infections virales peuvent influer sur la réponse de l hôte. Il existe des données épidémiologiques corrélant la survenue d infection bactérienne sévère et d infection virale. Par exemple le rythme saisonnier d infection invasive à pneumocoque corrèle fidèlement à celui d infections virales à influenza et au virus respiratoire syncitial (VRS). 21,22 Les mécanismes liant infections virale et bactérienne sont multiples, et nous en discuterons certains plus loin (voir III. discussion et perspectives). Le hasard pourrait être lui aussi un facteur influençant la réponse de l hôte. Ainsi, comment expliquer le concept de dose létale 50 (LD50), c'est-à-dire la dose nécessaire pour tuer 50% d une population? Par exemple, comment expliquer que chez des souris parfaitement identiques sur le plan génétique (animaux syngéniques), une concentration identique du même pathogène induit la mort de 50% d animaux, alors que l autre moitié échappera à l infection? Il a été montré que parmi des souris Balb/c, avec le même patrimoine génétique, 19% des gènes ont un niveau d expression variant de 1.5 fois entre animaux, c est ce que l on nomme la variation génétique normale, ou un effet stochastique. 23 La reconnaissance par l hôte de bactéries et l activation de leucocytes (les macrophages, les polynucléaires neutrophiles (PMN), les monocytes, les cellules dendritiques, les cellules NK) va avoir des conséquences physiopathologiques multiples (Figure 1). Ainsi, bien que leur fonction initiale soit d identifier et de détruire les agents pathogènes, ces cellules activées de l immunité innée vont induire une cascade d atteinte cellulaire et tissulaire, de troubles de la coagulation et potentiellement d une dysfonction d organes. Ces conséquences sont induites par l intermédiaire de la sécrétion de 7
 Inflammation (TNF, IL-1, IL-8, PAF) Anti-inflammation (IL-10, stnfrs, IL-1ra) Coagulation")
9 cytokines pro- et anti-inflammatoires, de l expression de protéines de surface, et de protéines circulantes. 24 Bactéries Reconnaissance innée des bactéries Cellules de l immunité innée (macrophages, PMNs, endothélium) Inflammation (TNF, IL-1, IL-8, PAF) Anti-inflammation (IL-10, stnfrs, IL-1ra) Coagulation (tissue factor, thrombin, PAI-1) Vasodilatation (inos - NO) Dysfonction Métabolique (mitochondrie) Lésions tissulaires Immuno-suppression CIVD (microthromboses, saignement) Choc (hypoperfusion tissulaire) Dysfonction d organes Figure 1. La reconnaissance innée des bactéries par les cellules de l immunité innée va induire une réponse systémique de l hôte et potentiellement mener à une dysfonction d organes. La sécrétion de cytokines pro- et anti-inflammatoires va générer des lésions tissulaires et provoquer, dans un deuxième temps, un état d immuno-paralysie. L activation de la coagulation secondaire à une dysrégulation de l équilibre hémostatique, peut mener au développement d un état de coagulation intravasculaire disséminée (CIVD). La sécrétion de substances vasodilatatrices, l augmentation de la perméabilité capillaire, le développement d une insuffisance circulatoire associée à des perturbations du métabolisme tissulaire vont mener à une hypoperfusion tissulaire et une anoxie cellulaire. 24,25 III.2 Immunité Innée L intensité et la qualité de la réponse de l hôte aux bactéries dépendent avant tout des mécanismes de reconnaissance, qu ils soient spécifiques ou non. Ces dernières font référence au système immunitaire adaptatif et au système immunitaire inné. 8
10 L une des spécificités immunologiques uniques aux vertébrés est la présence d un système capable de générer une réponse immunitaire spécifique, et d induire une mémoire immunologique de l infection. Ce système, appelé «immunité adaptative», est capable d adapter sa capacité de réponse à chaque contact successif d un agent pathogène donné avec l hôte. Les composants principaux de l immunité adaptative sont les lymphocytes et leurs produits, les anticorps. Le mécanisme de reconnaissance par le système immunitaire adaptatif est basé sur la reconnaissance spécifique de fragments de structure moléculaire (protéines, peptides, hydrates de carbones) par des anticorps. Ces anticorps obtenus après réarrangement de segments de gènes et production clonale permet d induire une réaction de l hôte retardée secondaire à l activation d effecteur. Ainsi, trois phases sont décrites : la phase de reconnaissance, phase d activation et phase effectrice. La phase de reconnaissance consiste en la liaison d antigènes à des récepteurs spécifiques (anticorps) liés ou non à des lymphocytes matures. Les lymphocytes B expriment des anticorps à leur surface cellulaire qui lient aussi bien des protéines «étrangères» que des sucres, des lipides ou autres structures chimiques. Les lymphocytes T expriment à leur surface des récepteurs capables de reconnaître de courtes séquences peptidiques issues des antigènes. Par ailleurs, les lymphocytes T ont la caractéristique unique de pouvoir reconnaître et réagir uniquement à des peptides présentés par d autres cellules (cellules présentatrices d antigènes, ou APC). La phase d activation de la réponse immunitaire adaptative consiste en la séquence d évènements induits sur le lymphocyte par la reconnaissance spécifique de l antigène. Tous les lymphocytes subissent une phase de prolifération permettant une expansion clonale, suivie d une différentiation vers une cellule effectrice (dont la finalité est d éliminer l antigène), ou une cellule «mémoire». Certains lymphocytes ne vont pas se différencier et vont mourir. Durant la phase effectrice de la réponse immunitaire adaptative, les lymphocytes préalablement activés vont induire les mécanismes d élimination de l antigène. Au contraire de l immunité adaptative constituée d un système hautement sophistiqué retrouvé uniquement chez les vertébrés et nécessitant un réarrangement génétique, l immunité innée est elle phylogénétiquement conservée et est présente chez quasi tous les organismes multicellulaire. 26 Le système immunitaire inné est caractérisé par une action rapide de molécules effectrices et de leucocytes permettant de limiter la multiplication de bactéries pathogéniques et de les détruire. 27,28 Toutefois, l immunité innée et l immunité adaptative interagissent continuellement. Certains effecteurs sont communs : Par exemple, la liaison d antigènes par des anticorps induit la phagocytose, et l activation du système du complément. Les lymphocytes T activés sécrètent des cytokines qui vont induire une réponse inflammatoire, et ce au même titre que la réponse immunitaire innée. Ainsi, bien que les anticorps soient habituellement considérés comme faisant partie du système immunitaire adaptatif, les anticorps naturels (NAbs) représentent une classe à part d anticorps car ils sont capables de détecter des antigènes que l hôte n a «jamais vu». 29 Parmi les premières descriptions d NAbs, notons les travaux de Landsteiner (prix Nobel de Médecine en 1930) qui décrivit la présence d agglutinines anti-a et anti-b dans le sérum humain de groupe sanguin alternatif (1). Ce sont (1) 9
11 principalement des IgM qui sont produites, prioritairement par les lymphocytes B1 (CD5+). Les NAbs sont codés par les gènes de type «germline variable» (V) avec peu de mutations somatiques, 80% possède un gène V H de la famille V H3 et > 90% comportent une chaîne légère lambda. Ces anticorps ont donc un répertoire restreint mais qui est hautement conservé dans l évolution 30. Les NAbs ont la capacité de neutraliser directement un pathogène, mais aussi d activer le système du complément et de former des complexes antigène-anticorps secondairement éliminés par le système réticuloendothélial permettant ainsi de prévenir la prolifération de pathogènes et d induire une réponse immunitaire dans des organes lymphoïdes secondaires. Les NAbs ont clairement montré, du moins chez l animal, un rôle essentiel dans la réponse de l hôte à des pathogènes tels que Streptococcus pneumoniae, Nesseiria meningitidis, Salmonella typhimurium, mais aussi contre différents virus 31. Le système du complément ainsi que les NAbs constituent les deux systèmes de défense centraux dans la réponse dites «humorale» (la fameuse «balle magique» en référence aux travaux de Behring, Kitsao, et Ehrlich 32 ), déterminant ainsi un lien entre l immunité adaptative et innée. Le système du complément est apparu tôt dans l évolution, probablement il y a plus de 500 millions d années 33. C est un système complexe, constitué de plus de 30 protéines sériques et récepteurs cellulaires et qui joue un rôle central dans la réponse de l hôte aux infections. Il y a trois manières d activer le système du complément : 1) la «voie classique», par l intermédiaire d anticorps (IgM), de pentraxines (Pentraxin 3, PTX3 ; protéine C-réactive, CRP ; serum amyloïde protéine, SAP) et du C1q, 2) la «voie des lectines» à partir de complexes multimériques constitués de lectines (tels que le mannose-binding lectine, MBL ; les protéines du surfactant A et D, SP-A et SP-D), de ficolines (L-, M-, et H-ficoline) et d oligosaccharide d origine bactérienne (mannose, N-acetylglucosamine, fucose) et 3) la «voie alterne», activée spontanément à partir de l hydrolyse de C3. 34,35,36,37 La reconnaissance par le système du complément initie une cascade protéolytique résultant en l induction d une réponse inflammatoire (anaphylatoxines, C3a, C4a, C5a), de la constitution d un complexe d attaque membranaire (MAC : C5b6789n), de l initiation de la phagocytose (MBL, C1q, C3b, ic3b, C4b) et de la stimulation de lymphocytes B (C3d). 36 Les NAbs ainsi que le système du complément sont ainsi à la «croisée» de la réponse immunitaire innée et adaptative, mêlant phagocytose, inflammation, lyse cellulaire et apoptose, présentation des antigènes, et stimulation des lymphocytes B. Il y a plus d un siècle, Metchnikoff recevait le Prix Nobel de Médecine (1908) pour l identification des phagocytes et la caractérisation de la phagocytose initiant la compréhension des mécanismes cellulaire de l immunité innée (2). La théorie de la phagocytose développée par Metchnikoff a pour origine une expérience effectuée à Messine en A cette occasion, il démontra qu après ponction d une larve d étoile de mer avec une épine de rose, des cellules mobiles, issues de la larve, s agglutinaient autour de l épine comme si elles tentaient de l ingérer. Cette expérience représenta une révolution conceptuelle par laquelle des particules «étrangères» ou des agents pathogènes étaient capables d induire chez l hôte une réaction inflammatoire aboutissant à la phagocytose de ceux-ci. Bien que ce ne soit pas Metchnikoff, mais Osler qui décrivit le premier la phagocytose bactérienne 38, Metchnikoff porte le crédit d avoir compris le rôle central de la phagocytose dans les (2) 10
12 mécanismes de défense de l hôte contre les infections et est considéré comme le père de l immunité innée. Dans une publication datant de 1884, Metchnikoff rapporte l observation selon laquelle des cellules amoeboïdes s accumulent autour d acrospores ayant pénétré le tube digestif de daphnea, les mouches d eau, et la capacité de ces cellules de phagocyter et digérer ces spores. 39. Le conflit entre «humoralistes» et «cellularistes» comme Metchnikoff sera sujet à une violente polémique, magnifiquement illustrée par les discours de Nobel de Metchnikoff en Ce seront Wright et Douglas qui réconcilieront les théories humorale et cellulaire, en montrant que des facteurs (protéines de l hôte) accélèrent et favorisent le processus de phagocytose. Nous leur devons le concept des opsonines et de l opsonophagocytose (du grec «opsono» signifiant «préparer des victuailles pour») 39. Ainsi, le système immunitaire inné constitue le premier échelon des défenses contre l infection. Il est activé rapidement, aussitôt qu un pathogène traverse les barrières naturelles de l hôte, tels que la peau ou les épithéliums. 27 Il est constitué d une part de protéines solubles (protéines de la phase aiguë, cytokines, protéines du complément, lectines, etc ) et d autre part de cellules dites «sentinelles» que sont les monocytes, les macrophages, les polynucléaires neutrophiles, les cellules dendritiques et les cellules NK). Il y a 20 ans, Charles A. Janeway proposa l existence d un mécanisme de reconnaissance immun inné capable d identifier des structures moléculaires, ou motifs, conservés exprimés par divers pathogènes (bactéries, champignon, virus) et non par les cellules de l hôte, qu il intitula pathogen-associated molecular patterns, ou PAMPs. 40 Ces motifs aussi variés que la flagelline, le lipopolysaccharide (LPS), le peptidoglycan (PGN), l acide lipotechoique (LTA), les lipopeptides di- ou tri-acylés, l ADN non-méthylé avec des motifs CpG, l ARN simple-brin (sbarn), ou double-brin (dbarn), etc.. sont autant de structures chimiques spécifiquement reconnues par des récepteurs nommés pattern recognition receptors, ou PRRs. 41,42 Parmi les PRRs reconnaissant des motifs bactériens nous en citerons quatre types. La famille des scavenger receptors (SR) est composée d un grand nombre de protéines membranaires (par exemple, MSR1 ou SR-A, MARCO) participant à l endocytose de ligands polyanioniques, telles que des lipoprotéines ou lipopolysaccharides (LTA, LPS). La famille des C-type lectin receptors (CLR) regroupe 17 groupes de récepteurs membranaires (tels que le mannose receptor, macrophage galactose CLR, Dectin 1 et 2, Langerin) ou solubles (fibronectine, vitronectine). Une discussion détaillée des SR et des CLR est faite dans l article 2 (voir III.1.2). Les Toll-like receptors (TLRs) ainsi que les nucleotide-binding oligomerization domain (Nod) proteins constituent d importants groupes de PRRs. Les TLRs sont situés à la surface cellulaire ou dans différents compartiments intra-cellulaires, tels que le Golgi ou le réticulum endoplasmique (par exemple le TLR3), les protéines Nods ne se trouvant que dans le cytoplasme. 27,43 Nous discuterons spécifiquement la fonction des TLRs dans le chapitre suivant (voir II.3). Les protéines «Nod» sont ainsi capables de reconnaître des bactéries dans le compartiment cytoplasmique par le biais de constituant bactériens de surface. Les deux protéines «Nod» les mieux connues sont Nod1 et Nod2, deux membres de la famille des mammalian nucleotide binding-site 11
13 leucine-rich repeat (NBS-LRR). Les protéines du groupe NBS-LRR, se caractérisent par la présence d un segment riche en leucine et sont représentées outre Nod1 et Nod2, par IPAF, et les protéines Nalp (Nalp 1-14), ainsi que le MHC class II transactivator (CIITA). Une certaine homologie se retrouve avec les protéines du groupe des hélicases, tels que RIG-1, et Mda-5. L ensemble des NBS-LRR et des hélicases constitue la famille des Nod-like receptors (NLRs). Cette famille de récepteurs est capable de reconnaître de nombreux ligands bactériens (tels que les peptidoglycan (PGN), la flagelline, la toxine de l anthrax et l ARN bactérien), l ADN viral double-brin (dsdna), mais aussi des ligands non-bactériens, tels que des cristaux d acide urique, mais aussi d asbeste et d hydroxyde d aluminium. 44,45 L une des caractéristiques des NLRs est de posséder un domaine de signalisation, tels que caspase-activating and recruitment domain (CARD) et PYRIN domain. Par ailleurs, à l exception de RIG-1 et de Mda-5, les NLRs contiennent un domaine «Nod». Nod1 est ubiquitaire dans les tissus humains et possède un domaine CARD. Nod1 reconnaît spécifiquement un muropeptide issu de peptidoglycans provenant de bactéries à Gram négatif contenant un motif tripeptidique N-acetylglucosamine-N-acetylmuramic acid (GlcNAc-MurNAc). 46 Nod2 est principalement exprimé dans les cellules myélomonocytaire. Il est structurellement proche de Nod1 à l exception qu il contient deux domaines CARD. 47 Nod2 reconnait un muramly dipeptide (MDP) dérivé du peptidoglycan : MurNAc-L-Ala-D-isoGln, que l on retrouve aussi bien chez les bactéries à Gram négatif qu à Gram positif. Ces deux récepteurs ont un rôle important dans la reconnaissance de nombreux pathogènes. Ainsi, Nod1 a été identifié comme un récepteur des peptidoglycans de Listeria monocytogenes, Pseudomonas aeruginosa, Shigella flexneri, Chlamydia pneumoniae, et Helicobacter pylori Pour sa part, Nod2 reconnait les peptidoglycans de Staphylococcus aureus, Mycobacterium tuberculosis, Streptococcus pneumoniae et Salmonella typhimurium Par ailleurs, la flagelline de Salmonella typhimurium est reconnue par IPAF et ce indépendamment de TLR5 (voir II.3). 54,55 L importance des NLRs dans la réponse de l hôte aux bactéries est illustrée par la corrélation entre une mutation de NOD2 (frameshift et missense) et la maladie de Crohn (30% des patients souffrant de cette maladie ont une mutation de NOD2). 47,56 III.3 Récepteurs Toll-like Le premier membre de la famille des récepteurs Toll, «Drosophila Toll, dtoll», a été découvert comme étant l un des 12 gènes maternels nécessaires pour la formation de l axe dorso-ventral chez l embryon de la mouche de vinaigre. 57 Il a été ensuite découvert que dtoll était le récepteur d une protéine «Spätzle». La stimulation du récepteur transmembranaire dtoll par Spätzle active une serine/thréonine kinase cytoplasmique, Pelle, via la protéine adaptatrice, Tube. L activation de Pelle permet la dégradation du complexe Cactus-Dorsal. Une fois que Cactus est dégradé, Dorsal, un facteur de transcription, est transloqué dans le noyau activant l expression de gènes nécessaires à la formation de l axe dorso-ventral de la mouche. Ces kinases sont les équivalents chez la mouche des kinases impliquées dans la voie de signalisation «NF- B» chez les mammifères. C est en 1996, que le rôle de dtoll dans la réponse contre un pathogène fut identifiée, initialement contre les 12
14 champignons (Aspergillus fumigatus), puis contre les bactéries à Gram positif. 58,59 Les mouches déficientes en dtoll sont incapables de se défendre contre ces pathogènes et meurent d une infection généralisée. 58,59 Rapidement, des protéines homologues à dtoll sont identifiées chez l homme et d autres mammifères et intitulées TLRs pour «Toll-like receptors». 60 Les TLRs sont des glycoprotéines transmembranaires de type 1, et sur la base d une grande homologie de leur région cytoplasmique, font partie d une superfamille contenant le récepteur à l IL-1 (IL-1R). Toutefois la région extracellulaire des TLRs et de IL-1R diffèrent : les TLRs sont essentiellement constitués d une région riche en leucine (LRR), tandis que la région extracellulaire de IL-1R contient trois domaines successifs de type immunoglobuline-like. Le domaine extracellulaire des TLRs est constitué de 16 à 28 répétions du motif LRR. Ces répétitions «leucine-rich» forment un brin-β (β-strand) et une hélice-α (α-helix) reliés par une boucle, constituant ainsi une structure en forme de «fer-à-cheval» directement impliqué dans la reconnaissance des ligands (voir II.5). 61,62 Des LRRs se retrouvent dans plus de 6'000 protéines (3) impliquées dans la réponse immunitaire (TLR, NLR, CD14, IL1-R, etc.), la transduction de signaux intracellulaires, la régulation du cycle cellulaire, etc.. 63 L analyse des séquences et des structures cristallines des LRRs a permis de les classifier en sept sous-familles. 64 Les TLRs constituent une sous-famille contenant des modules LRR constitués du motif suivant : XLXXLXXLXLXXNXLXXLPXXXFX. La structure cristalline des domaines LRR permet de distinguer deux groupes de TLRs. TLR1, 2 et 4 ont une structure conformationelle légèrement différente des autres TLRs. 65,66 Ils présentent deux transitions structurelles sur le brin-β, le divisant ainsi en trois sous-domaines : N-terminal, central et C-terminal. De plus le domaine central ne possède pas l ossature constituée d asparagine (au même titre que TLR6 et TLR10) permettant ainsi une «distorsion» de la structure, observée pour TLR1, TLR2 et TLR4. L extrémité intracytoplasmique des TLRs contient une région d environ 150 acides aminés avec un fort degré d homologie avec celle du récepteur à IL-1 (IL-1R). Cette région a été nommée domaine «TIR» pour «Toll/IL-1R région». Les structures cristallines des domaines TIR ont été décrites pour TLR1, TLR2, et TLR Elles ont en commun la présence de cinq brins-β parallèles entourées par cinq hélices-α. L analyse par mutation de cette région a permis de mettre en évidence le rôle central de la boucle BB («BB loop») connectant le second brin-β (brin β-b) et la seconde hélice-α (hélice α- B). 68,69 La boucle BB joue un rôle dans la dimérisation des domaines TIR et/ou le recrutement des protéines adaptatrices, essentielles dans l activation des voies de signalisation intra-cellulaires. 69,70 Actuellement treize TLRs différents ont été décrits chez les mammifères, dont 10 chez l homme (TLR1 à -10). Nous venons de voir que la famille de TLRs est structurellement caractérisée par la présence d un domaine extracellulaire LRR, et d une région intracytoplasmique contenant un domaine TIR. L analyse des séquences d acides aminés ainsi que de la structure génomique des TLRs chez l homme permet de les subdiviser en cinq sous-familles. La première, contenant TLR2, TLR1, TLR6 et (3) 13
15 TLR10 se caractérise par la présence d un seul exon codant (TLR2 possède trois exons, mais seul un est codant). Par ailleurs, TLR6 et TLR1 sont localisé en tandem sur le chromosome 4p14 et présente une très haute similarité dans les séquences d acides aminés, ceci suggérant que TLR1 et TLR6 pourrait être issu d une duplication au cours de l évolution. 71 Il est par ailleurs intéressant de noter que cette sous-famille est aussi caractérisée par l absence d une ossature asparagine (voir ci-dessus) rendant la structure du domaine LRR moins rigide. Les TLRs de la sous-famille contenant TLR7, TLR8 et TLR9 sont codés par deux exons. TLR7 et TLR8 sont aussi structurellement très proches et localisés en tandem sur le chromosome Xp22. 72,73 TLR4 et TLR5 sont respectivement codés par quatre et cinq exons. TLR3 possède 5 exons, mais n est codé que par les exons 2 à Le premier TLR à avoir été identifié chez l homme fut TLR4 par le groupe de Janeway, préalablement appelé htoll pour «human Toll». 60. Toutefois ce n est qu une année plus tard en 1998, que Poltorak et al. identifièrent Tlr4 comme étant impliqué dans la reconnaissance du LPS. 74 Basé sur l observation selon laquelle les souris C3H/HeJ et C57BL/10ScCr ne répondent pas au LPS, mais présente une très grande susceptibilité aux bactéries à Gram négatif, des mutations (missense et mutation null, respectivement) touchant Tlr4 ont été identifiées suggérant le rôle de TLR4 dans la reconnaissance du LPS. 74 Poltorak et Beutler ont en effet démontré qu une proline mutée dans le domaine TIR de TLR4 des souris C3H/HeJ expliquait leur résistance au LPS, et qu une délétion d une partie de TLR4 était responsable de l insensibilité des souris C57BL/10ScCr au LPS. 74 TLR4 participe aussi à la reconnaissance de protéines virales, fongiques, et parasitaires. Il a été montré que TLR4 en association avec CD14 est capable de reconnaître la protéine de fusion du virus respiratoire syncitial (VRS). 75 Les polysaccharides capsulaires issus de Cryptococcus neoformans, les mannans de Candida albicans, les glycoinositolphospholipides de Trypanosoma cruzi seraient également des ligands microbiens de TLR4. TLR2 reconnaît et lie les lipopeptides bactériens. La capacité de TLR2 de reconnaître les peptidoglycans (PGN), un constituant important des parois bactériennes, reste discutée dans la mesure où une contamination des préparations de PGN par d autres ligands de TLR2 (LTA, lipoprotéines) est possible. 76 TLR2 forme un heterodimère avec TLR1 ou TLR6. TLR2:TLR1 reconnaît les lipopetides di- et tri-acylés, alors que TLR2:TLR6 ne reconnaît que les lipopeptides di-acylés Récemment la structure cristalline des domaines extracellulaires de TLR2 en association avec TLR1 et le lipopeptide synthétique tri-acylé Pam 3 CSK 4 a été caractérisée. 65 L hétérodimérisation de TLR2 et TLR1 nécessite la présence du ligand, et forme une structure constituant un «M» avec les extrémités N-terminal des deux TLRs sur les extrémités. Cette structure est identique à celle des homodimères TLR3 et du dimère TLR4 associé à LPS et MD-2. 64,81 TLR2 a ainsi la capacité de reconnaître une grande variété de microorganismes par le biais de molécules de surface bactériennes conservées. Ceci inclus les lipoprotéines de bactéries à Gram négatif, mycoplasmes et spirochètes, les peptidoglycans et l acide lipotechoïque des bactéries à Gram négatif, le lipoarabinomannan des mycobactéries, les glycoinositolphospholipides (protéine TC52) de Trypanosoma cruzi, des lysophosphatidylsérine shistosomales, le zymosan et des phopholipomannanes des champignons, 14
16 des glycolipides de Treponema maltophilum, des porines de Nesseiria meningitidis et Hemophilus influenzae. 28,41 De plus TLR2 semble reconnaître le LPS de Legionella pneumophila, Leptospira interrogans (LPS penta-acylé) et Porphyromonas gingivalis. 82,83 Toutefois le risque d une contamination des extraits de LPS par des lipopeptides est réel. 84 De plus la capacité de reconnaissance de bactéries à Gram négatif par TLR2 (que se soit à partir de lipopeptides ou de LPS) semble être limitée en comparaison à la reconnaissance dépendante de MD-2:TLR4, dans la mesure ou des concentrations bactériennes beaucoup plus importantes sont nécessaires pour activer la réponse TLR2-induite par rapport à TLR4. 85 TLR2 est capable de reconnaître aussi des PAMPs issus de virus, tels que les hémagglutinines du virus de la rougeole, et l herpès simplex-1 (HSV-1). 86,87 TLR5 reconnaît spécifiquement la flagelline, le principal constituant protéique des flagelles bactériens. 88,89 Les flagelles s étendent de la paroi externe des bactéries à Gram négatif, leur permettant de se propulser, mais aussi de se lier aux cellules hôtes, participant ainsi à l invasion des tissus. TLR3 reconnaît l ARN double-brin (dbarn) produit par bon nombre de virus durant leur cycle de réplication. L acide polyinosinique-polycytidylique ou poly(i :C), un ARN double brin synthétique, a lui aussi la capacité de lier TLR TLR3 a clairement été lié à la réponse de l hôte contre HSV La structure cristalline de TLR3 lié à de l dbarn a récemment été caractérisée. 91 L ARN double-brin interagit avec les extrémités N-terminal et C-terminal de TLR3. A la différence de la liaison entre TLR2:TLR1 et les lipopeptides ou les interactions hydrophobiques jouent un rôle majeur, l ARN double brin se lie à TLR3 par des liaisons ioniques et hydrogènes. 65,91 Alors que TLR3 reconnaît l ARN double-brin, TLR7 et TLR8 reconnaissent l ARN simple-brin 92 et semble être essentiel dans la réponse de l hôte contre le virus influenza A. 93 Des données récentes suggèrent que le TLR7, 8 et 9 pourraient participer à la pathogénèse de maladie auto-immune, tel que le lupus érythémateux disséminé. 94,95 TLR9 détecte les motifs CpG de l ADN non-méthylé bactérien, fongique, ainsi que l haemazoine issu de Plasmodium. 96,97,98,99 Par ailleurs TLR9 reconnaît aussi Herpes simplex Le ligand de TLR10 reste pour l instant inconnu. L activation des TLRs entraine l expression de nombreuses cytokines et chémokines. La nature de cette réponse dépend de la voie de signalisation activée, laquelle est déterminée par des protéines adaptatrices qui se lient directement au domaine «TIR» des TLRs : MyD88, MAL(ou TIRAP), TRIF, TRAM et SARM. Ces protéines adaptatrices, au nombre de cinq, sont couplées en aval à des protéines kinases qui vont phosphoryler des cibles moléculaires en cascade pour finalement entrainer l activation de facteurs de transcriptions tel que Nuclear Factor kappab (NF-κB) et des membres de la famille des interferon (IFN) regulatory factors (IRFs). Ces cinq protéines adaptatrices contiennent, tout comme les TLRs, un domaine «TIR». 41,101,102 La plupart des réponses pro-inflammatoires induites par les TLRs (TLR1, 2, 6, 4, 7, 8, 9) sont issues de la stimulation d une voie commune, médiée par la protéine adaptatrice MyD88 (Figure 2). Après stimulation d un TLR, MyD88 se lie au domaine «TIR» et recrute différentes isoformes de l IL-1 receptor associated kinase (IRAK). IRAK4 est par exemple particulièrement important dans la réponse 15
17 induite par l IL-1. Comme précédemment mentionné, un déficit en IRAK4 a été reporté chez des patients souffrant d infections bactériennes à répétition. 18 L activation d IRAK entraine une liaison avec Tumor Necrosis Factor (TNF) Receptor-Associated Factor 6 (TRAF6), et l activation de différentes voies d activation intracellulaire : a) c-jun N-terminal protein kinase (JNK) par les mitogenactivated protein kinase kinase (MKK) -4 et -7, b) p38 (via MKK3/6), c) ERK1/2 (via MKK1/2), d) des voies d activation dépendantes du facteur de transcription nuclear factor-kappab (NF-κB), ainsi que e) l induction par interferon (IFN) regulatory factor (IRF) 1, 5, 7. Ces derniers vont permettre l expression d IFN de type 1 et de nombreux gènes de cytokines impliqués dans la réponse inflammatoire (TNF-α, IL-8, IL-1, etc..). Il est intéressant de noter que l induction de p38 et JNK induite par la stimulation TLRs:MyD88-dépendante va aussi participer à la stabilisation des ARN messagers de TNF-α. 103 Figure 2. Protéines adaptatrices impliquées dans la signalisation des TLRs. TLR, Toll like receptor ; TIRAP, Toll/IL-1R domain-containing adaptor protein ; TRIF, TIR-containing adaptorinducing IFN-β ; TRAM, TRIF-related adaptor molecule; SARM, sterile α- and armadillo-motifcontaining protein. TIRAP (Toll/IL-1R domain-containing adaptor protein), aussi connu sous l acronyme «Mal» (MyD88- adaptor-like), coopère avec MyD88 et est nécessaire pour la signalisation induite par TLR2 et TLR4 (Figure 3). 104 La voie de signalisation MyD88-indépendante liée à l activation LPS:MD-2:TLR4 et dbarn:tlr3 permet l activation du facteur de transcription IRF3 et l expression d IFN-β. 105 TRIF (TIR-containing adaptor-inducing IFN-β) est nécessaire pour l activation de la signalisation MyD88- indépendante et l activation d IRF Il est intéressant de noter que les souris doublement déficientes en MyD88 et en Trif perdent toute capacité de réponse au LPS, suggérant que les deux voies de signalisation sont les seules liées à TLR TRAM (TRIF-related adaptor molecule) est un co-adapteur de la signalisation MyD88-indépendante avec TRIF et IRF SARM (sterile α- and armadillo-motif-containing protein) est la protéine adaptatrice la plus ancienne sur le plan de l évolution, étant la seule présente chez le ver Caenorhabiditis elegans. 109,110 SARM, à l inverse des quatre autres protéines adaptatrices, est un régulateur négatif de l activation de NF-κB et des IRFs. Ainsi, SARM permet le blocage spécifique de l activation des facteurs de transcription induite par TRIF, sans altérer les voies de signalisation MyD88-dépendantes. 111 De nombreux signaux (NLR, 16
18 CLR Dectin-1, tyrosine kinase et phosphatase) interagissent avec la signalisation des TLRs (voir article 2). 3,102 Figure 3. Représentation schématique de la signalisation MyD88 dépendante et indépendante par TLR4. LPS, lipopolysaccharide ; TLR4, Toll-like receptor 4 ; TIRAP, Toll/IL-1R domain-containing adaptor protein ; IRAK, IL-1 Receptor-Associated Kinase; TRAF, Tumor Necrosis Factor Receptor-Associated Factor 6; TAK1, Transforming growth factor β activated kinase 1; TRIF, TIR-containing adaptor-inducing IFN-β ; TRAM, TRIF-related adaptor molecule; TBK1, Tumor necrosis factor receptor associated factor (TRAF) family member associated Nuclear factor-kappab (NF- B) activator binding kinase 1; SARM, sterile α- and armadillo-motif-containing protein; IRF, interferon regulatory factor; IKK, inhibitor of NFκB kinase; Akt, protein kinase B ; MAP3K, Mitogen-activated protein kinase kinase kinase; ERK, extracellular signal regulated kinase; JNK, Jun N terminal kinase; AP-1, activated protein-1. La réponse «TLR» est régulée par certaines protéines accessoires agissant au niveau de la reconnaissance bactérienne, de la signalisation, et de la réponse. 112 Parmi celles-ci, RP105, une 17
19 protéine transmembranaire de type 1 (comme les TLRs) contenant un domaine extracellulaire de type LRR. RP105 se trouve à la surface des lymphocytes B, et est associé à MD-1 (qui présente une grande homologie avec MD-2). MD-1 est nécessaire à l expression de surface de RP105, dont le complexe MD-1:RP105 interagit directement avec MD-2:TLR4 et TLR2 113 Le rôle de MD-1:RP105 semble varier selon le type cellulaire. Il régule positivement la réponse TLR2 et TLR4 dans les lymphocytes B, et négativement dans les macrophages et les cellules dendritiques. 114 Cette fonction régulatrice pourrait avoir un rôle dans le développement du lupus érythémateux disséminé à anticorps antinucléaires négatifs 115 UNC93B est une protéine conservée chez les mammifères et est située dans le réticulum endoplasmique. UNC93B interagit spécifiquement avec TLR3,7 et 9 permettant leur transfert du réticulum endoplasmique aux endolysosomes. 116 PRAT4A est aussi une protéine du réticulum endoplasmique, indispensable à l expression membranaire de TLR4 et TLR1, mais aussi endolysosomiale de TLR9, TLR7, mais pas de TLR La glycoprotéine 96 (gp96) est indispensable à la maturation réticulo-endoplasmique des TLRs. 118 CD36 fait partie des récepteurs «scavenger» (voir article 2) 3. Parallèlement à sa fonction dans les mécanismes de phagocytose, CD36 régule la réponse TLR2/TLR6 aux lipopeptides di-acylés, ainsi qu au LTA. 119 CD36 forme une liaison hétérotopique avec TLR2/TLR6 et leurs ligands. 120 HMGB1 (high mobility group box-1) est une protéine nucléaire libérée lors de la nécrose et/ou souffrance cellulaire. HMGB1 dans le milieu extracellulaire est capable d induire une réponse inflammatoire, la réparation tissulaire, de recruter des cellules pluripotentes et d induire leur prolifération. 121 HMGB1 est un co-facteur de TLR9, capable de lier l ADN microbien ainsi que le receptor for advanced glycation end-products (RAGEs). HMGB1 est ainsi recruté dans des vésicules contenant TLR9 et permet d accélérer la redistribution de TLR9 du réticulum endoplasmique aux endosomes précoces et la formation de complexe CpG ADN:TLR9. 98 LL37 est un peptide antimicrobien participant au mécanisme auto-immun observé lors de psoriasis. En effet, LL37 est capable de convertir de l ADN endogène amorphe en un inducteur TLR9-dépendant reconnu par les cellules dendritiques plasmocytoïdes. 122 III.4 Structure et fonction de MD-2 Au même titre que CD14, dont nous parlerons plus loin, MD-2 est une protéine accessoire essentielle à la reconnaissance des ligands de TLR4. Par ailleurs MD-2 est une protéine chaperonne de TLR4 permettant sa glycosylation et son transport intracellulaire («cell trafficking»), en tout cas chez la souris. 123, MD-2, pour myeloid differentiation factor-2, anciennement appelé Ly96, est une glycoprotéine de 160 acides aminés dont le gène se trouve sur le chromosome 8q Le gène MD-2 est constitué chez tous les vertébrés de 5 exons et de 3 introns. MD-2 existe lié à TLR4 à la surface cellulaire et sous forme soluble dans les liquides biologiques. La forme soluble recombinante de MD-2 est un mélange en solution de monomères, dimères et multimères Toutefois, seule la forme monomérique est capable de se lier à TLR4 et de jouer son rôle de co-facteur dans la réponse au LPS. 131 La raison de 18
20 la tendance de MD-2 à ce multimériser (ponts di-sulfure) est inconnue. Le MD-2 produit par bacculovirus est principalement monomérique et dimérique. 132 MD-2 soluble migre sur SDS-PAGE en condition réduite en deux bandes (25 et 30 KDa). Un traitement par endoglycosidase réduit la migration d une seule bande à 18KDa. 130 MD-2 a deux sites de N-glycosylation (N 26 et N 114 ), confirmé par analyse de mutant. 123,133 MD-2 monoglycosylé ou aglycosylé interagissent avec TLR4, sont capable de lier LPS et d induire une signalisation, suggérant que le niveau de glycosylation n est pas directement impliqué dans la reconnaissance du LPS et l initiation de la signalisation. La première publication suggérant que MD-2 lie le LPS est celle de Viriyakosol et al. 134 Figure 4. Structure du complexe MD-2/TLR4 humain. 81 La dimérisation de l hétérodimère constitué par MD-2/TLR4 forme un «M». Ce complexe est stabilisé par des interactions hydrophobes, hydrogènes et des ponts phosphates entre MD-2, et MD-2:LPS et TLR4* de l hétérodimère complémentaire, ainsi qu entre LPS et TLR4, TLR4* et MD-2. (4) (4) Crystal Structure of the Human Tlr4-Human Md-2-E.Coli Lps Ra Complex. [MMDB ID:70004; PDB ID: 3FXI]. 19
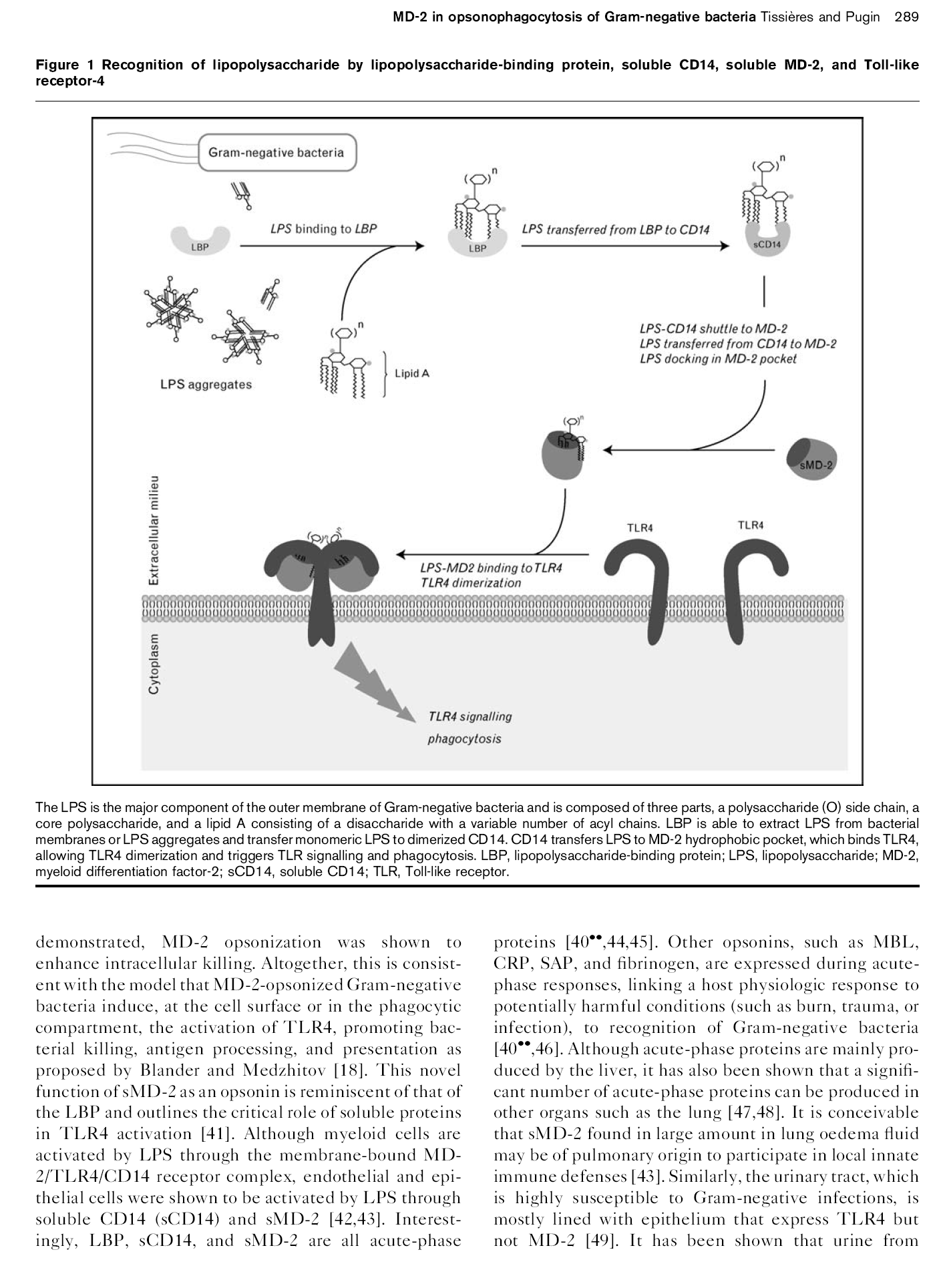
21 La caractérisation de liaison du LPS au MD-2 est récente. 66,135 L activité endotoxinique maximale a été identifiée avec un LPS dont le lipide A est hexa-acylé (6 chaînes d acides gras) et contient 2 groupes phosphates, alors que le lipide A penta- ou hepta-acylé est 100 fois moins actif. 136,137 L Eritoran (E5564) est un dérivé synthétique tétra-acylé du lipide A issu du LPS non-pathogénique de Rhodobacter sphaeroides 138. L Eritoran est un puissant antagoniste MD-2/TLR4 bloquant la réponse au LPS, actuellement en étude de phase III dans le traitement du sepsis sévère (5). Un autre Lipide A tétra-acylé, le lipide IVa, possède une activité antagoniste avec MD-2:TLR4 humain, mais partiel chez la souris. La structure cristalline de TLR4-VLR avec Eritoran et de MD-2 avec le Lipide IVa a récemment été publiée. 66,135 MD-2 possède une poche constituée par deux feuillets β antiparallèles, reliés sur l un des côtés, avec des résidus hydrophobes sur leur face interne («hydrophobic groove»). Cette poche hydrophobe est bordée en son ouverture de résidus chargés positivement. Ainsi la structure et les caractéristiques électrostatiques sont favorables pour des ligands ampiphatiques, tel que le LPS. La poche hydrophobique du MD-2 est capable de contenir les quatre chaînes acylaires du Lipid IVa et 5 des 6 chaînes d un LPS «commun». MD-2 interagit avec TLR4 principalement à partir de liaisons ioniques et hydrogènes. Ces deux travaux ont ainsi montré que le récepteur du LPS est principalement MD-2 et que LPS ne lie pas directement à TLR4. 66,135 Le mécanisme de dimérisation et d activation de MD-2:TLR4 a été récemment caractérisé. 81 Ainsi, cinq des six chaînes acylaires du LPS s insèrent dans la poche hydrophobique, la sixième chaîne formant une liaison hydrophobe avec TLR4*. L insertion des cinq chaînes acylaires dans la poche hydrophobique provoque une modification structurelle de MD-2 permettant son interaction avec TLR4* et provoquant ainsi la dimérisation des complexes LPS:MD-2:TLR4. 81 Une vue schématique des interactions moléculaires LPS:MD-2:TLR4 est résumée dans la Figure 4. III.5 Reconnaissance des bactéries à Gram négatif par TLR4:MD-2 Les infections à bactéries à Gram négatif ont de particulier que le principal constituant de leur paroi externe, l endotoxine bactérienne ou LPS, est capable d induire chez l humain une réaction systémique majeure avec lésions tissulaires, coagulation intra-vasculaire et état de choc. Le système immunitaire inné est capable de détecter des concentrations de LPS de l ordre du picomolaire et d induire l expression de centaines de gènes. 139,140 La reconnaissance du LPS par les cellules des mammifères implique plusieurs protéines principalement le LPS-binding protein (LBP), CD14, MD-2 et TLR4. Les lipopolysaccharides sont constitués d un lipide A et d une partie polysaccharidique débordant de la membrane externe (Figure 5). Le lipide A est constitué de deux sucres aminés (glucosamine, 3- aminoglucosamine) liés à des acides gras hydroxylés et non-hydroxylés. La nature des sucres aminés (5) ClinicalTrials.gov identifier number: NCT
22 et des acides gras, ainsi que le nombre de chaînes lipidiques varient selon l espèce bactérienne. La fraction polysaccharidique est constituée du noyau («core») interne (constitué d heptose et d acide 3- désoxy-d-manno-2-octulosnique, ou KDO) lié au lipide A, et d un noyau externe (nombre variable d hexose et hexosamine) lié à une chaîne polysaccharidique terminale, appelée chaîne O spécifique ou antigène O («O-antigen»). Chez certaines bactéries à Gram négatif, telles que Neisseria et certains Haemophilus, le LPS est remplacé par un lipo-oligosaccharide constitué uniquement du lipide A et des noyaux interne et externe. La chaîne O spécifique est la partie la plus variable du LPS dont la taille va définir le LPS rugueux («rough LPS» = chaîne courte ou absente) et le LPS lisse («smooth LPS» = chaîne longue). Ainsi, le LPS est une molécule amphiphile formant de larges agrégats en solution, des plans ressemblant à des membranes, voire des micelles. Figure 5. La paroi bactérienne est constituée, de l intérieur à l extérieur, d une membrane plasmique, séparée de peptidoglycans présents dans l espace périplasmique, puis de la membrane externe, dont le feuillet externe est constitué à > 90% de lipopolysaccharide (LPS), et de diverses protéines. Le LPS est constitué d un lipide A, d un core et d un antigène O d une longueur 21
23 varaiable. Le core ainsi que l antigène O sont constitués de divers polysaccharides. KDO, acide 3- désoxy-d-manno-2-octulosnique ; Hep, heptose ; Gal, galactose ; Glu, glucose ; GluNAc, acide N acetylglutamic ; Rha, rhamnose ; Man, mannose ; Abe, abedose. La caractérisation récente de la structure cristalline du complexe LPS:MD-2:TLR4 suggère que seul la forme monomérique du LPS lie MD-2:TLR4. 81 La «désagrégation» du LPS nécessite LBP. LBP est une glycoprotéine sérique de kda. C est une protéine de phase aiguë de type I synthétisée par le foie, mais aussi par les cellules épithéliales et musculaires. 141 Le LBP appartient à la famille des lipid transfer/lbp comprenant aussi BPI (bactericidal/permeability-increasing protein), CETP (cholesteryl ester-binding protein), et PLTP (phospholipid transfer protein). La cristallisation de la structure de BPI a permis (homologie >40% avec LBP) d identifier à l extrémité N-terminale de LBP une niche de résidus cationique nécessaire pour la désagrégation et la reconnaissance du LPS L extrémité C-terminale est elle nécessaire à l interaction entre LBP et CD Le LBP a une double fonction lors de son interaction avec LPS : à faible concentration, LBP accentue la signalisation en l extrayant des parois bactériennes et en formant un complexe LBP:LPS monomérique, alors qu à haute concentration, LBP inhibe la réponse en transportant le LPS à des lipoprotéines (HDL) et en formant des agrégats. 146,147 Le CD14 («cluster of differentiation 14») est une glycoprotéine qui existe aussi bien ancrée à la membrane sous forme «GPI-anchored», que sous forme soluble. Le CD14 est nécessaire pour la reconnaissance de faibles concentrations de LPS. 148 CD14 est un PRR capable de lier des phospholides anioniques et de participer à leur transport La structure cristalline de CD14 montre qu il s agit d une protéine de la famille des LRR, ressemblant à un «fer à cheval», et constituée d un brin β sur la surface concave et d hélice α sur la surface convexe. 152 Le segment aminoterminal, impliqué dans la liaison au LPS, circoncit une poche hydrophobique permettant d y recevoir les chaînes acylaires du lipide A. 153 CD14 est aussi capable de lier les chaînes polysacchardiques du LPS et des peptidoglycans. 154,155 CD14 est une protéine accessoire influençant la réponse des TLRs. 156 Récemment, il a été montré que le CD14 était nécessaire pour l activation par le LPS lisse de la signalisation TLR4-MyD88 indépendant. 157 Ainsi, sans CD14, le lipide A active TLR4 et permet le recrutement uniquement de MyD88 et MAL, alors qu en présence de LPS lisse, l absence de CD14 ne permet pas l activation de TLR4. Toutefois, en présence de CD14, aussi bien le LPS rugueux que lisse sont capables de recruter la voie de signalisation MyD88:MAL et TRIF:TRAM. CD14 est aussi capable d amplifier la réponse TLR2, l activation de TLR2:TLR6 (conformation du dimère permettant le recrutement de MyD88) nécessitant la présence de CD Ainsi, le LPS lie LBP et le CD14 avec une grande affinité (constantes d association de l ordre du nanomolaire), permettant ainsi son transport jusqu à MD-2, la protéine réceptrice de l endotoxine, qui va avec TLR4 former un double hétérodimère (LPS:MD-2:TLR4:LPS*:MD-2*:TLR4*). Cette dimérisation semble être dépendante du ligand bactérien LPS. 66,156,158 De plus il semble aussi que c est cette dimérisation («ligand-dependent») qui rapproche les domaines TIR des 2 molécules de 22
24 TLR4, induisant la cascade de signalisation. 81 La haute spécificité de la reconnaissance du LPS par le complexe MD-2/TLR4 est en ligne avec le fait que ce récepteur joue un rôle central dans la réponse de l hôte aux bactéries à Gram négatif. Ainsi, l endotoxine bactérienne peut être reconnue aussi bien par des cellules immunes «professionnelles» tels que les macrophages, les monocytes et les neutrophiles, que par des cellules endothéliales ou des cellules épithéliales exprimant une ou plusieurs protéines constituant le récepteur à l endotoxine. Les protéines «manquantes» (CD14, MD- 2) peuvent être «amenées» par le plasma par exemple ou elles se trouvent sous forme soluble (Figure 6). Figure 6. Représentation schématique de la reconnaissance par les cellules myélo-monocytaires, l endothélium et les cellules épithéliales de l endotoxine bactérienne. LPS, lipopolysaccharide/ endotoxine ; LBP, LPS-binding protein ; scd14, CD14 soluble ; smd-2, MD-2 soluble ; TLR4, Tolllike receptor. Les mécanismes de reconnaissance varient entre différents tissus et ce afin de répondre à des contraintes physiologiques spécifiques. Par exemple, certains tissus épithéliaux sont exposés en permanence à une charge bactérienne importante (épithélium intestinal, épithélium respiratoire) alors 23
25 que d autres (épithélium urinaire ou méningé) se situent dans un environnement physiologiquement stérile. Il a été ainsi montré que les cellules d origine myélo-monocytaires répondaient au LPS par la présence à leur surface de MD-2 lié au récepteur TLR4, et de CD Au même titre, l endothélium possède le complexe MD-2:TLR4 à sa surface, toutefois l activation par le LPS nécessite du CD14 sous forme soluble, présent dans le plasma et dans certains liquides biologiques. compréhension des mécanismes de réponse de l épithélium à l endotoxine bactérienne permet de mettre en perspective bon nombre de pathologies infectieuses, telles que les infections bactériennes respiratoires, digestives, méningées et urinaires. L épithélium, au sens histologique, ne peut pas en situation physiologique répondre à l endotoxine bactérienne sans exposer l hôte à un état inflammatoire continu. Il n exprime donc pas de CD14, ni de MD-2, mais peut dans certaines circonstances exprimer TLR4. Ainsi, l épithélium respiratoire peut exprimer à sa surface TLR4 en réponse à différents stimuli, lors d une infection virale, par exemple. 160 C est cette inductibilité de TLR4 qui permet, potentiellement, à l épithélium de répondre au LPS, et ce seulement si les cofacteurs solubles CD14 et MD-2 sont présents La La fonction de la forme soluble de MD-2 (smd-2) a récemment pris une perspective supplémentaire. Il a été montré que le plasma, ainsi que les liquides biologiques (œdème pulmonaire et urine) de patients souffrant de sepsis contenaient du smd-2 capable d induire une réponse au LPS et aux bactéries à Gram négatif par des cellules épithéliales n exprimant que TLR4. 162,164,165 Ces travaux suggèrent que l induction de MD-2, lors d une infection systémique, a un rôle physiologique central dans les mécanismes de défenses contre les bactéries à Gram négatif. 3 Dès lors, l étude de la fonction et de la régulation de l expression de MD-2, la protéine réceptrice de l endotoxine bactérienne, prend tout son sens. 24
26 IV. IV.1 RESULTATS MD-2 est une protéine de phase aiguë et une opsonine pour les bactéries à Gram négatif L augmentation des concentrations de MD-2 soluble dans le plasma de patients au cours d un épisode infectieux grave suggère que cette protéine pourrait être sécrétée en réponse à une réaction de type «phase aiguë». 162 La réaction de phase aiguë est caractérisée par la sécrétion, principalement par le foie, de diverses protéines en réponse à une agression telle qu un traumatisme, une brûlure étendue, ou une infection. 166 Cette sécrétion est principalement induite par deux cytokines, l IL-1β et l IL-6. Un grand nombre de protéines circulantes sont des protéines de la phase aiguë. Parmi celles-ci citons les protéines du compléments C3, C4, C9, le facteur B, le C1 inhibiteur, et le mannose-binding lectin (MBL), des protéines majeures de la coagulation comme le fibrinogène, le plasminogène, le plasminogen-activator inhibitor 1 (PAI-1), mais aussi un grand nombres d anti-protéases, de protéines de transport (haptoglobine, céruloplasmine) et des protéines elles-mêmes directement liées avec la réponse inflammatoire tels que la phospholipase A2, l interleukin-1 receptor antagonist (IL1-RA), ou le granulocyte colony-stimulating factor (G-CSF). Plusieurs protéines de phase aiguë jouent un rôle dans l opsonophagocytose de bactéries, tels que la protéine sérum amyloïde A (SAA), la fibronectine, le MBL, le C3b, la protéine C-réactive, ainsi que deux protéines du complexe de reconnaissance du LPS : le LBP et le CD14 soluble. 167,168 Notre hypothèse expérimentale initiale était de déterminer si MD-2 soluble est une protéine de phase aiguë. 25
27 IV.1.1 Article 1: MD-2 is an acute phase protein and an opsonin for Gram-negative bacteria. (MD-2 est une protéine de phase aiguë et une opsonine pour les bactéries à Gram négatif) Nous avons initialement établi que l induction in vivo chez la souris C57BL/6 d une réaction de phase aiguë par injection sous-cutanée de turpentine, induisait la production hépatocytaire de MD-2 (figure 3). L inductibilité dite «de phase aiguë», c'est-à-dire dépendante d IL-6, a été démontrée en induisant une réaction de phase aiguë chez des souris C57BL/6 wild type et des souris C57BL/6 IL-6 -/- (IL-6 k.o.). L absence d induction de l expression de MD-2 lors d une induction de phase aiguë chez les souris IL6 -/- et non chez les wild-type suggère que le MD-2 soluble est une protéine de phase aiguë. Nous avons ensuite montré par qpcr, western blot, et gène reporteur assay sur différentes lignées cellulaires (THP-1, HepG2), que MD-2 étant induit par IL-6 mais non par l IL-1β, permettant de conclure que MD-2 est une protéine phase aiguë de type II (type I : inductible par IL-6 et IL.1β). Nous avons décrit la cinétique de sécrétion de smd-2 et avons déterminé la fonction du MD-2 soluble. Nous nous sommes d abord intéressés à la capacité de MD-2 soluble de se lier à des bactéries à Gram négatif. En utilisant un MD-2 humain soluble «taggé» His («6xHis tag») à des concentrations croissantes, nous avons montré par cytométrie en flux (FACS) que smd-2 se liait de manière dosedépendante à des bactéries à Gram négatif (E.coli), mais pas à des bactéries à Gram positif (S.aureus). Trois fonctions possibles de MD-2 soluble ont été testées : un potentiel effet bactéricide de smd-2, sa capacité d induire une inflammation en présence de bactéries à Gram négatif, et un rôle putatif comme opsonine. En utilisant un test de croissance bactérienne avec ou sans smd-2, aucune capacité bactéricide n a pu être mise en évidence. Toutefois, le smd-2 permet la reconnaissance de bactéries à Gram négatif par des cellules épithéliales n exprimant que TLR4 (HEK293 TLR4+/MD2-) et d induire une réaction inflammatoire. La fonction opsonophagocytaire du MD-2 soluble a été démontrée en utilisant un test dans lequel la phagocytose de bactéries à Gram négatif fluorescentes par des granulocytes humains a été mesurée par FACS après 5, 10 ou 20 minutes d opsonisation avec du MD-2 soluble. Ce travail a ainsi permis de décrire pour la première fois la fonction opsonisante de MD-2 soluble et sa caractérisation comme étant une protéine de phase aiguë de type II. Tissières P, Dunn-Siegrist I, Nobre V, Elson G, Schappi M, Comte R, Pugin J. Soluble MD-2 is an acute phase protein and an opsonin for Gram negative bacteria. Blood 2008;111(4):
28 27
29 28
30 29
31 30
32 31
33 32
34 33
35 34
36 35

37 36
38 IV.1.2 Article 2: Role of MD-2 in the opsonophagocytosis of Gram-negative bacteria. (Rôle de MD-2 dans l opsonophagocytose de bactéries à Gram négatif) Le rôle de MD-2 dans la phagocytose de bactéries à Gram négatif et de fait dans le contrôle de l infection, change fondamentalement la compréhension de la fonction des mécanismes de reconnaissance innée des pathogènes par les TLRs. Il devient apparent que les TLRs, et dans notre cas MD-2:TLR4, sont des éléments essentiels dans les mécanismes de phagocytose, de clearance bactérienne, et de présentation des antigènes au système immunitaire adaptatif. 169 Ainsi, l observation faite selon laquelle les souris Md-2 -/- ont une résistance augmentée à l endotoxine, mais une susceptibilité accrue aux infections à bactéries à Gram négatif, prend toute sa valeur. 170 La mise en perspective de l interaction de MD-2 soluble, une nouvelle opsonine, avec les mécanismes de phagocytose habituels (IgG, système du complément, etc.), a été récemment résumée dans l article suivant. Tissières P, Pugin J. The role of MD-2 in the opsonophagocytosis of Gram-negative bacteria. Curr Opin Infect Dis 2009;22(3):
39 38
40 39
41 40
42 41
43 42

44 43
COUSIN Fabien KERGOURLAY Gilles. 19 octobre 2007. de l hôte par les. Master 2 MFA Responsable : UE Incidence des paramètres environnementaux
 COUSIN Fabien KERGOURLAY Gilles 19 octobre 2007 Inhibition des défenses de l hôte par les bactéries pathogènes Master 2 MFA Responsable : UE Incidence des paramètres environnementaux Gwennola ERMEL I Détection
COUSIN Fabien KERGOURLAY Gilles 19 octobre 2007 Inhibition des défenses de l hôte par les bactéries pathogènes Master 2 MFA Responsable : UE Incidence des paramètres environnementaux Gwennola ERMEL I Détection
Les cytokines et leurs récepteurs. Laurence Guglielmi laurence.guglielmi@univ-montp1.frli
 Les cytokines et leurs récepteurs Laurence Guglielmi laurence.guglielmi@univ-montp1.frli l i@ i 1 Les cytokines et leurs récepteurs 2 mécanismes principaux d interactions cellulaires : - contact membranaire
Les cytokines et leurs récepteurs Laurence Guglielmi laurence.guglielmi@univ-montp1.frli l i@ i 1 Les cytokines et leurs récepteurs 2 mécanismes principaux d interactions cellulaires : - contact membranaire
Vaccinologie et Stratégie de Développement des Vaccins
 Vaccinologie et Stratégie de Développement des Vaccins Vaccinologie 1977 - Terme utilisé par Jonas Salk. 1983 - Rassemble tous les aspects des vaccinations : biologiques, immunologiques et cliniques ainsi
Vaccinologie et Stratégie de Développement des Vaccins Vaccinologie 1977 - Terme utilisé par Jonas Salk. 1983 - Rassemble tous les aspects des vaccinations : biologiques, immunologiques et cliniques ainsi
Le rôle de l endocytose dans les processus pathologiques
 UE7 Cours n 9 C. LAMAZE 24.11.11 Elise GODEAU (partie1) Guillaume MERGENTHALER (partie2) Le rôle de l endocytose dans les processus pathologiques SOMMAIRE : I. L endocytose à récepteurs : la voie des clathrines
UE7 Cours n 9 C. LAMAZE 24.11.11 Elise GODEAU (partie1) Guillaume MERGENTHALER (partie2) Le rôle de l endocytose dans les processus pathologiques SOMMAIRE : I. L endocytose à récepteurs : la voie des clathrines
Observer : l'absence de noyau des hématies la petite taille des plaquettes la forme et la taille des noyaux (leucocytes) ACTIVITES ELEVES TS
 ACTIVITES ELEVES TS") 4.1 La réaction inflammatoire, un exemple de réponse innée 4.1.1 Les cellules de défense au microscope optique : Par l'observation d'une goutte de sang humain au microscope, repérer chaque type de cellule
4.1 La réaction inflammatoire, un exemple de réponse innée 4.1.1 Les cellules de défense au microscope optique : Par l'observation d'une goutte de sang humain au microscope, repérer chaque type de cellule
Explorations des réponses Immunitaires. L3 Médecine
 2012 Explorations des réponses Immunitaires L3 Médecine Rappel sur les réponses Immunitaires DIFFERENTS TYPES DE REPONSES IMMUNITAIRES Naturelle = innée Adaptative Non spécifique Spécifique Immédiate Barrière
2012 Explorations des réponses Immunitaires L3 Médecine Rappel sur les réponses Immunitaires DIFFERENTS TYPES DE REPONSES IMMUNITAIRES Naturelle = innée Adaptative Non spécifique Spécifique Immédiate Barrière
Cibles et mécanismes d action des traitements par cytokines et anti-cytokines
 Cibles et mécanismes d action des traitements par cytokines et anti-cytokines Jean Daniel Lelièvre, Yves Lévy, Pierre Miossec I-Introduction... 2 II-Les interférons... 2 II-1.L interféron... 3 II-1-a.
Cibles et mécanismes d action des traitements par cytokines et anti-cytokines Jean Daniel Lelièvre, Yves Lévy, Pierre Miossec I-Introduction... 2 II-Les interférons... 2 II-1.L interféron... 3 II-1-a.
IMMUNOLOGIE. La spécificité des immunoglobulines et des récepteurs T. Informations scientifiques
 IMMUNOLOGIE La spécificité des immunoglobulines et des récepteurs T Informations scientifiques L infection par le VIH entraîne des réactions immunitaires de l organisme qui se traduisent par la production
IMMUNOLOGIE La spécificité des immunoglobulines et des récepteurs T Informations scientifiques L infection par le VIH entraîne des réactions immunitaires de l organisme qui se traduisent par la production
Vaccins du futur Atelier «Croisière dans l archipel des nouveaux vaccins»
 Vaccins du futur Atelier «Croisière dans l archipel des nouveaux vaccins» Bernard Duval Institut National de Santé Publique du Québec 18 avril 2002 Objectifs Grands axes de la réponse immunitaire Conjugaison
Vaccins du futur Atelier «Croisière dans l archipel des nouveaux vaccins» Bernard Duval Institut National de Santé Publique du Québec 18 avril 2002 Objectifs Grands axes de la réponse immunitaire Conjugaison
EXERCICES : MECANISMES DE L IMMUNITE : pages 406 407 408 409 410
 EXERCICES : MECANISMES DE L IMMUNITE : pages 406 407 408 409 410 EXERCICE 1 PAGE 406 : EXPERIENCES A INTERPRETER Question : rôles respectifs du thymus et de la moelle osseuse dans la production des lymphocytes.
EXERCICES : MECANISMES DE L IMMUNITE : pages 406 407 408 409 410 EXERCICE 1 PAGE 406 : EXPERIENCES A INTERPRETER Question : rôles respectifs du thymus et de la moelle osseuse dans la production des lymphocytes.
Contrôle et modulation de la réponse immunitaire par Brucella abortus
 AIX-MARSEILLE UNIVERSITE Faculté des sciences de Luminy Ecole Doctorale des Sciences de la Vie et de la Santé THESE DE DOCTORAT Biologie - Spécialité : Immunologie En vue d'obtenir le titre de DOCTEUR
AIX-MARSEILLE UNIVERSITE Faculté des sciences de Luminy Ecole Doctorale des Sciences de la Vie et de la Santé THESE DE DOCTORAT Biologie - Spécialité : Immunologie En vue d'obtenir le titre de DOCTEUR
Cellules procaryotes Service histologie Pr.k.mebarek
 Cellules procaryotes Service histologie Pr.k.mebarek I) Les cellules procaryotes II) Les cellules eucaryotes o 1) Caractéristiques générales des cellules eucaryotes o 2) Organisation des cellules eucaryotes
Cellules procaryotes Service histologie Pr.k.mebarek I) Les cellules procaryotes II) Les cellules eucaryotes o 1) Caractéristiques générales des cellules eucaryotes o 2) Organisation des cellules eucaryotes
Le Monde des insectes, son importance pour l Homme et l apport de l étude des insectes aux sciences du vivant.
 MEDAILLE D OR DU CNRS 2011 Le Monde des insectes, son importance pour l Homme et l apport de l étude des insectes aux sciences du vivant. Extrait de l allocution prononcée le 19 juin 2007 par Jules Hoffmann,
MEDAILLE D OR DU CNRS 2011 Le Monde des insectes, son importance pour l Homme et l apport de l étude des insectes aux sciences du vivant. Extrait de l allocution prononcée le 19 juin 2007 par Jules Hoffmann,
DIAPOSITIVE 1 Cette présentation a trait à la réglementation sur les thérapies cellulaires.
 Produits de thérapie cellulaire DIAPOSITIVE 1 Cette présentation a trait à la réglementation sur les thérapies cellulaires. DIAPOSITIVE 2 La fabrication des thérapies cellulaires est examinée par la Division
Produits de thérapie cellulaire DIAPOSITIVE 1 Cette présentation a trait à la réglementation sur les thérapies cellulaires. DIAPOSITIVE 2 La fabrication des thérapies cellulaires est examinée par la Division
Hépatite chronique B Moyens thérapeutiques
 Hépatite chronique B Moyens thérapeutiques Dr Olfa BAHRI Laboratoire de Virologie Clinique Institut Pasteur de Tunis INTRODUCTION Plus de 300. 10 6 porteurs chroniques de VHB dans le monde Hépatite chronique
Hépatite chronique B Moyens thérapeutiques Dr Olfa BAHRI Laboratoire de Virologie Clinique Institut Pasteur de Tunis INTRODUCTION Plus de 300. 10 6 porteurs chroniques de VHB dans le monde Hépatite chronique
Rôle de l'acétylation du facteur de transcription IRF3
 Université de Montréal Rôle de l'acétylation du facteur de transcription IRF3 par Tasheen Wissanji Département de Pharmacie Faculté de Pharmacie Mémoire présenté à la Faculté des études supérieures en
Université de Montréal Rôle de l'acétylation du facteur de transcription IRF3 par Tasheen Wissanji Département de Pharmacie Faculté de Pharmacie Mémoire présenté à la Faculté des études supérieures en
Dr E. CHEVRET UE2.1 2013-2014. Aperçu général sur l architecture et les fonctions cellulaires
 Aperçu général sur l architecture et les fonctions cellulaires I. Introduction II. Les microscopes 1. Le microscope optique 2. Le microscope à fluorescence 3. Le microscope confocal 4. Le microscope électronique
Aperçu général sur l architecture et les fonctions cellulaires I. Introduction II. Les microscopes 1. Le microscope optique 2. Le microscope à fluorescence 3. Le microscope confocal 4. Le microscope électronique
Identification de mécanismes de régulation des fonctions des interférons: Rôle de la palmitoylation du récepteur de l interféron de type I
 Identification de mécanismes de régulation des fonctions des interférons: Rôle de la palmitoylation du récepteur de l interféron de type I Julie Claudinon To cite this version: Julie Claudinon. Identification
Identification de mécanismes de régulation des fonctions des interférons: Rôle de la palmitoylation du récepteur de l interféron de type I Julie Claudinon To cite this version: Julie Claudinon. Identification
Transfusions sanguines, greffes et transplantations
 Transfusions sanguines, greffes et transplantations Chiffres clés en 2008 La greffe d organes est pratiquée depuis plus de 50 ans. 4 620 malades ont été greffés. 1 563 personnes ont été prélevées. 222
Transfusions sanguines, greffes et transplantations Chiffres clés en 2008 La greffe d organes est pratiquée depuis plus de 50 ans. 4 620 malades ont été greffés. 1 563 personnes ont été prélevées. 222
- 2 - faire industriel dans la mise au point des produits biologiques. L Institut Roche de Recherche et Médecine Translationnelle (IRRMT, basé à
 Information presse Roche, l Agence Nationale de Recherches sur le Sida et les hépatites virales (ANRS), le Baylor Research Institute (BRI) et Inserm Transfert mettent en place une coopération stratégique
Information presse Roche, l Agence Nationale de Recherches sur le Sida et les hépatites virales (ANRS), le Baylor Research Institute (BRI) et Inserm Transfert mettent en place une coopération stratégique
Rôle des acides biliaires dans la régulation de l homéostasie du glucose : implication de FXR dans la cellule bêta-pancréatique
 Rôle des acides biliaires dans la régulation de l homéostasie du glucose : implication de FXR dans la cellule bêta-pancréatique Tuteur : Anne Muhr-Tailleux cardiovasculaires et diabète (Equipe 1) Institut
Rôle des acides biliaires dans la régulation de l homéostasie du glucose : implication de FXR dans la cellule bêta-pancréatique Tuteur : Anne Muhr-Tailleux cardiovasculaires et diabète (Equipe 1) Institut
Dr Pascale Vergne-Salle Service de Rhumatologie, CHU de Limoges. Membre enseignant chercheur EA 4021
 Dr Pascale Vergne-Salle Service de Rhumatologie, CHU de Limoges Membre enseignant chercheur EA 4021 Rhumatisme inflammatoire chronique Prévalence des SA ies en Europe = 1 à 2% Age moy : 26 ans, ratio homme/femme:
Dr Pascale Vergne-Salle Service de Rhumatologie, CHU de Limoges Membre enseignant chercheur EA 4021 Rhumatisme inflammatoire chronique Prévalence des SA ies en Europe = 1 à 2% Age moy : 26 ans, ratio homme/femme:
Les Infections Associées aux Soins
 Les Infections Associées aux Soins Dr Catherine Sartor EOH Conception Assistance Publique Hôpitaux de Marseille Ecole IFSI, Marseille, 12 septembre 2014 Définitions 2007 Infection Nosocomiale (IN) 1999
Les Infections Associées aux Soins Dr Catherine Sartor EOH Conception Assistance Publique Hôpitaux de Marseille Ecole IFSI, Marseille, 12 septembre 2014 Définitions 2007 Infection Nosocomiale (IN) 1999
POURQUOI L HYGIENE HYGIENE = PROPRETE HYGIENE = PREVENTION DES INFECTIONS COMMUNAUTAIRES ET DES INFECTIONS ASSOCIEES AUX SOINS
 Prise en charge d un «résident septique» en Etablissements Médico Sociaux PRÉVENTION DES RISQUES INFECTIEUX EN ETABLISSEMENTS MÉDICO SOCIAUX INFECTIONS NOSOCOMIALES ET COMMUNAUTAIRES ASSOCIÉES AUX SOINS
Prise en charge d un «résident septique» en Etablissements Médico Sociaux PRÉVENTION DES RISQUES INFECTIEUX EN ETABLISSEMENTS MÉDICO SOCIAUX INFECTIONS NOSOCOMIALES ET COMMUNAUTAIRES ASSOCIÉES AUX SOINS
Travaux dirigés de Microbiologie Master I Sciences des Génomes et des Organismes Janvier 2015
 Andrew Tolonen atolonen@genoscope.cns.fr Travaux dirigés de Microbiologie Master I Sciences des Génomes et des Organismes Janvier 2015 A- Généralités I- La vie sur terre telle que nous la connaissons ne
Andrew Tolonen atolonen@genoscope.cns.fr Travaux dirigés de Microbiologie Master I Sciences des Génomes et des Organismes Janvier 2015 A- Généralités I- La vie sur terre telle que nous la connaissons ne
Mécanisme des réactions inflammatoires
 01/04/2014 THOMASSIN Guillaume L2 Revêtement Cutané Dr. Sophie Deplat-Jégo Relecteur 4 8 pages Revêtement cutané Mécanisme des réactions inflammatoires cutanés Mécanisme des réactions inflammatoires Plan
01/04/2014 THOMASSIN Guillaume L2 Revêtement Cutané Dr. Sophie Deplat-Jégo Relecteur 4 8 pages Revêtement cutané Mécanisme des réactions inflammatoires cutanés Mécanisme des réactions inflammatoires Plan
Transgene accorde une option de licence exclusive pour le développement et la commercialisation de son produit d immunothérapie TG4010
 Parc d Innovation d Illkirch, France, le 10 mars 2010 Transgene accorde une option de licence exclusive pour le développement et la commercialisation de son produit d immunothérapie TG4010 Transgene (Euronext
Parc d Innovation d Illkirch, France, le 10 mars 2010 Transgene accorde une option de licence exclusive pour le développement et la commercialisation de son produit d immunothérapie TG4010 Transgene (Euronext
Chapitre 7 : Structure de la cellule Le noyau cellulaire
 UE2 : Structure générale de la cellule Chapitre 7 : Structure de la cellule Le noyau cellulaire Professeur Michel SEVE Année universitaire 2010/2011 Université Joseph Fourier de Grenoble - Tous droits
UE2 : Structure générale de la cellule Chapitre 7 : Structure de la cellule Le noyau cellulaire Professeur Michel SEVE Année universitaire 2010/2011 Université Joseph Fourier de Grenoble - Tous droits
Cytokines & Chimiokines
 Cytokines & Chimiokines I. (D après Förster, R. et al. (1999) Cell 99:23) Dans le but d étudier la régulation de la circulation des leucocytes dans l organisme, des souris déficientes pour le récepteur
Cytokines & Chimiokines I. (D après Förster, R. et al. (1999) Cell 99:23) Dans le but d étudier la régulation de la circulation des leucocytes dans l organisme, des souris déficientes pour le récepteur
DÉFICITS IMMUNITAIRE COMMUN VARIABLE
 DÉFICITS IMMUNITAIRE COMMUN VARIABLE Le présent livret a été rédigé à l attention des patients et de leurs familles. Il ne doit pas remplacer les conseils d un spécialiste en immunologie. 1 Egalement Disponible
DÉFICITS IMMUNITAIRE COMMUN VARIABLE Le présent livret a été rédigé à l attention des patients et de leurs familles. Il ne doit pas remplacer les conseils d un spécialiste en immunologie. 1 Egalement Disponible
Première partie: Restitution + Compréhension (08 points)
") Lycée M hamdia Année scolaire : 2011/2012 Prof : Saïd Mounir Date : 17/05/2012 Première partie: Restitution + Compréhension (08 points) EXERCIE N O 1: (4 points) : 1 : a-b 2 : b 3 : a-b 4 : d 5 : d 6 :
Lycée M hamdia Année scolaire : 2011/2012 Prof : Saïd Mounir Date : 17/05/2012 Première partie: Restitution + Compréhension (08 points) EXERCIE N O 1: (4 points) : 1 : a-b 2 : b 3 : a-b 4 : d 5 : d 6 :
Cytokines ; Chimiokines
 Cytokines ; Chimiokines I. Dans le but d étudier la régulation de la circulation des leucocytes dans l'organisme, des souris déficientes pour le récepteur CCR7 de chimiokine ont été générées par recombinaison
Cytokines ; Chimiokines I. Dans le but d étudier la régulation de la circulation des leucocytes dans l'organisme, des souris déficientes pour le récepteur CCR7 de chimiokine ont été générées par recombinaison
Les renseignements suivants sont destinés uniquement aux personnes qui ont reçu un diagnostic de cancer
 Information importante pour les personnes atteintes d un cancer du poumon non à petites cellules de stade avancé Les renseignements suivants sont destinés uniquement aux personnes qui ont reçu un diagnostic
Information importante pour les personnes atteintes d un cancer du poumon non à petites cellules de stade avancé Les renseignements suivants sont destinés uniquement aux personnes qui ont reçu un diagnostic
Anticorps, vaccins, immunothérapies allergéniques tout savoir sur les progrès de l immunothérapie en 20 questions
 Anticorps, vaccins, immunothérapies allergéniques tout savoir sur les progrès de l immunothérapie en 20 questions De quoi se compose le système immunitaire? Chaque jour, des substances étrangères, appelées
Anticorps, vaccins, immunothérapies allergéniques tout savoir sur les progrès de l immunothérapie en 20 questions De quoi se compose le système immunitaire? Chaque jour, des substances étrangères, appelées
Sommaire de la séquence 8
 Sommaire de la séquence 8 Nous avons découvert dans la séquence 7 que les micro-organismes sont présents partout dans notre environnement et qu à la faveur d une lésion, ils sont capables de franchir nos
Sommaire de la séquence 8 Nous avons découvert dans la séquence 7 que les micro-organismes sont présents partout dans notre environnement et qu à la faveur d une lésion, ils sont capables de franchir nos
Séquence 10. Le maintien de l intégrité de l organisme : quelques aspects de la réaction immunitaire. Sommaire
 Séquence 10 Le maintien de l intégrité de l organisme : quelques aspects de la réaction immunitaire Sommaire Chapitre 1. Pré-requis Chapitre 2. La réaction inflammatoire, un exemple de réponse innée Chapitre
Séquence 10 Le maintien de l intégrité de l organisme : quelques aspects de la réaction immunitaire Sommaire Chapitre 1. Pré-requis Chapitre 2. La réaction inflammatoire, un exemple de réponse innée Chapitre
Enseignement de Virologie (part. 2) Pr. Y. BAKRI Plan du cours
 Pr. Y. BAKRI Plan du cours") Université Mohammed V-Agdal Département de Biologie Faculté des Sciences Filière SVI Semestre 4 Année Universitaire : 2004-2005 Module optionnel: Biologie Humaine (M 16.1) Elément : Parasitologie - Virologie
Université Mohammed V-Agdal Département de Biologie Faculté des Sciences Filière SVI Semestre 4 Année Universitaire : 2004-2005 Module optionnel: Biologie Humaine (M 16.1) Elément : Parasitologie - Virologie
Chapitre III Le phénotype immunitaire au cours de la vie
 Chapitre III Le phénotype immunitaire au cours de la vie Le phénotype immunitaire d un individu caractérise sa capacité à répondre, grâce aux effecteurs de l immunité adaptative, aux différents agents
Chapitre III Le phénotype immunitaire au cours de la vie Le phénotype immunitaire d un individu caractérise sa capacité à répondre, grâce aux effecteurs de l immunité adaptative, aux différents agents
Sciences du vivant et Société. L inflammation silencieuse
 Sciences du vivant et Société L inflammation silencieuse Colloque du vendredi 12 décembre 2014 Présentent Dans le cadre du cycle de conférences «Sciences du Vivant et Société» L inflammation silencieuse
Sciences du vivant et Société L inflammation silencieuse Colloque du vendredi 12 décembre 2014 Présentent Dans le cadre du cycle de conférences «Sciences du Vivant et Société» L inflammation silencieuse
ECOLE PRATIQUE DES HAUTES ETUDES. MEMOIRE Présenté par Caroline FLAMENT
 MINISTERE DE L EDUCATION NATIONALE ECOLE PRATIQUE DES HAUTES ETUDES Science de la vie et de la terre MEMOIRE Présenté par Caroline FLAMENT Pour l obtention du diplôme de l Ecole Pratique des Hautes Etudes
MINISTERE DE L EDUCATION NATIONALE ECOLE PRATIQUE DES HAUTES ETUDES Science de la vie et de la terre MEMOIRE Présenté par Caroline FLAMENT Pour l obtention du diplôme de l Ecole Pratique des Hautes Etudes
De la physico-chimie à la radiobiologie: nouveaux acquis (I)
") De la physico-chimie à la radiobiologie: nouveaux acquis (I) Collaboration: - Laboratoire de Radiotoxicologie et Oncologie (L. Sabatier) CEA, DSV - Laboratoire de Génotoxicité et Modulation de l Expression
De la physico-chimie à la radiobiologie: nouveaux acquis (I) Collaboration: - Laboratoire de Radiotoxicologie et Oncologie (L. Sabatier) CEA, DSV - Laboratoire de Génotoxicité et Modulation de l Expression
ULBI 101 Biologie Cellulaire L1. Le Système Membranaire Interne
 ULBI 101 Biologie Cellulaire L1 Le Système Membranaire Interne De la nécessité d un SMI Le volume augmente comme le cube de la dimension linéaire, alors que la surface n'est augmentée que du carré Une
ULBI 101 Biologie Cellulaire L1 Le Système Membranaire Interne De la nécessité d un SMI Le volume augmente comme le cube de la dimension linéaire, alors que la surface n'est augmentée que du carré Une
Sérodiagnostic de la polyarthrite rhumatoïde
 1 ETSL Sérodiagnostic de la polyarthrite rhumatoïde TP 1 GABIN-GAUTHIER 13/11/2009 I. LA MALADIE... 2 II. TECHNIQUES QUALITATIVES... 2 1. PRINCIPE... 2 2. MODE OPERATOIRE... 3 2.1. WRST ou Waaler Rose
1 ETSL Sérodiagnostic de la polyarthrite rhumatoïde TP 1 GABIN-GAUTHIER 13/11/2009 I. LA MALADIE... 2 II. TECHNIQUES QUALITATIVES... 2 1. PRINCIPE... 2 2. MODE OPERATOIRE... 3 2.1. WRST ou Waaler Rose
Il est bien établi que le réseau d eau hospitalier peut
 Les micro-organismes de l eau impliqués dans les infections nosocomiales Depuis 2001 chez Pall, en charge de l information scientifique et réglementaire dans les domaines d applications couverts par la
Les micro-organismes de l eau impliqués dans les infections nosocomiales Depuis 2001 chez Pall, en charge de l information scientifique et réglementaire dans les domaines d applications couverts par la
CHAPITRE 3 LA SYNTHESE DES PROTEINES
 CHAITRE 3 LA SYNTHESE DES ROTEINES On sait qu un gène détient dans sa séquence nucléotidique, l information permettant la synthèse d un polypeptide. Ce dernier caractérisé par sa séquence d acides aminés
CHAITRE 3 LA SYNTHESE DES ROTEINES On sait qu un gène détient dans sa séquence nucléotidique, l information permettant la synthèse d un polypeptide. Ce dernier caractérisé par sa séquence d acides aminés
voies de signalisation
 Olivier Lascols Faculté de Médecine ierre et Marie Curie Généralités s sur les voies de signalisation 1 I. Caractérisation risation d une d voie de transduction d un d signal Exemple du Signal Mitogénique
Olivier Lascols Faculté de Médecine ierre et Marie Curie Généralités s sur les voies de signalisation 1 I. Caractérisation risation d une d voie de transduction d un d signal Exemple du Signal Mitogénique
STRUCTURE ET FONCTION DES PLURICELLULAIRES
 Plan de cours STRUCTURE ET FONCTION DES PLURICELLULAIRES 101-FYA-PT Pondération 3-1-2 Gilles Bourbonnais (C360) gilles.bourbonnais@dgpc.ulaval.ca Sciences de la Nature / PASC@L http://ici.cegep-ste-foy.qc.ca/profs/gbourbonnais/
Plan de cours STRUCTURE ET FONCTION DES PLURICELLULAIRES 101-FYA-PT Pondération 3-1-2 Gilles Bourbonnais (C360) gilles.bourbonnais@dgpc.ulaval.ca Sciences de la Nature / PASC@L http://ici.cegep-ste-foy.qc.ca/profs/gbourbonnais/
Médicaments du futur : Tendances et enjeux. Nicolas PY, Debiopharm International forumofac.14 26/09/2014
 Médicaments du futur : Tendances et enjeux Nicolas PY, Debiopharm International forumofac.14 26/09/2014 Quelques mots sur Debiopharm Groupe fondé en 1979 Siège à Lausanne 350 collaborateurs Financièrement
Médicaments du futur : Tendances et enjeux Nicolas PY, Debiopharm International forumofac.14 26/09/2014 Quelques mots sur Debiopharm Groupe fondé en 1979 Siège à Lausanne 350 collaborateurs Financièrement
Les nouveaux traitements du psoriasis
 Les nouveaux traitements du psoriasis Ci-après, vous trouverez 3 articles, parus récemment, parmi d'autres tout aussi intéressants, dans notre bulletin trimestriel Pso Magazine: - un extrait de l'exposé
Les nouveaux traitements du psoriasis Ci-après, vous trouverez 3 articles, parus récemment, parmi d'autres tout aussi intéressants, dans notre bulletin trimestriel Pso Magazine: - un extrait de l'exposé
PARTIE II : RISQUE INFECTIEUX ET PROTECTION DE L ORGANISME. Chapitre 1 : L Homme confronté aux microbes de son environnement
 PARTIE II : RISQUE INFECTIEUX ET PROTECTION DE L ORGANISME Chapitre 1 : L Homme confronté aux microbes de son environnement I- Les microbes dans notre environnement Qu est-ce qu un microbe? Où se trouvent-ils?
PARTIE II : RISQUE INFECTIEUX ET PROTECTION DE L ORGANISME Chapitre 1 : L Homme confronté aux microbes de son environnement I- Les microbes dans notre environnement Qu est-ce qu un microbe? Où se trouvent-ils?
NOCOSPRAY CARACTÉRISTIQUES TECHNIQUES UN FONCTIONNEMENT TRÈS SIMPLE DE MULTIPLES OPTIONS PERMETTANT DE S ADAPTER À CHAQUE SITUATION
 NOCOSPRAY Notre concept est né de l association Nocospray + Nocolyse (machine/produit) pour une désinfection automatisée sous forme gazeuse. Il permet de traiter de façon automatisée l air et les surfaces
NOCOSPRAY Notre concept est né de l association Nocospray + Nocolyse (machine/produit) pour une désinfection automatisée sous forme gazeuse. Il permet de traiter de façon automatisée l air et les surfaces
INSUFFISANCE HÉPATIQUE
 INSUFFISANCE HÉPATIQUE EXISTE-T-IL DES TRAITEMENTS À BASE DE CELLULES SOUCHES POUR L INSUFFISANCE HÉPATIQUE? Bien qu aucun traitement à base de cellules souches pour l insuffisance hépatique ne soit approuvé
INSUFFISANCE HÉPATIQUE EXISTE-T-IL DES TRAITEMENTS À BASE DE CELLULES SOUCHES POUR L INSUFFISANCE HÉPATIQUE? Bien qu aucun traitement à base de cellules souches pour l insuffisance hépatique ne soit approuvé
MINISTERE DE LA SANTE ET DES SOLIDARITES DIRECTION GENERALE DE LA SANTE- DDASS DE SEINE MARITIME
 Département des situations d urgence sanitaire Personne chargée du dossier : Evelyne FALIP/Nicole BOHIC Tél : 01 40 56 59 65/02 32 18 31 66 evelyne.falip@sante.gouv.fr MINISTERE DE LA SANTE ET DES SOLIDARITES
Département des situations d urgence sanitaire Personne chargée du dossier : Evelyne FALIP/Nicole BOHIC Tél : 01 40 56 59 65/02 32 18 31 66 evelyne.falip@sante.gouv.fr MINISTERE DE LA SANTE ET DES SOLIDARITES
PRINCIPALES ETAPES DE L'HEMATOPOIESE MEDULLAIRE (lignées, maturation, voies de domiciliation tissulaire) Partie II
 Partie II") Le 23/09/13 SOGHOMONIAN Astrid, L2 Tissu sanguin et système immunitaire Pr Baccini 22 pages PRINCIPALES ETAPES DE L'HEMATOPOIESE MEDULLAIRE (lignées, maturation, voies de domiciliation tissulaire) Partie
Le 23/09/13 SOGHOMONIAN Astrid, L2 Tissu sanguin et système immunitaire Pr Baccini 22 pages PRINCIPALES ETAPES DE L'HEMATOPOIESE MEDULLAIRE (lignées, maturation, voies de domiciliation tissulaire) Partie
Principales causes de décès selon le groupe d âge. 0 24 25 44 45 64 65 84 85 et plus
 Module 2 Exercice 1: Cellules souches hématopoïétiques 1. Causes de décès en Suisse (2010) La figure suivante montre les causes de décès les plus fréquentes en Suisse en 2010, telles qu elles ont été relevées
Module 2 Exercice 1: Cellules souches hématopoïétiques 1. Causes de décès en Suisse (2010) La figure suivante montre les causes de décès les plus fréquentes en Suisse en 2010, telles qu elles ont été relevées
LA MALADIE DE WALDENSTRÖM expliquée au néophyte
 LA MALADIE DE WALDENSTRÖM expliquée au néophyte Comment comprendre sa maladie de Waldenström lorsque l'on est ni médecin, ni biologiste? Bernard Cornillon, biochimiste à l'inserm, a rédigé ce document
LA MALADIE DE WALDENSTRÖM expliquée au néophyte Comment comprendre sa maladie de Waldenström lorsque l'on est ni médecin, ni biologiste? Bernard Cornillon, biochimiste à l'inserm, a rédigé ce document
CONTROVERSE : IDR OU QUANTIFERON LORS D'UN CONTAGE EN EHPAD?
 CONTROVERSE : IDR OU QUANTIFERON LORS D'UN CONTAGE EN EHPAD? Hélène MANGEARD François MALADRY Tuberculose : infection mycobactérienne Infection mycobactérienne chronique (M. Tuberculosis ++ ou bacille
CONTROVERSE : IDR OU QUANTIFERON LORS D'UN CONTAGE EN EHPAD? Hélène MANGEARD François MALADRY Tuberculose : infection mycobactérienne Infection mycobactérienne chronique (M. Tuberculosis ++ ou bacille
La lutte contre la tuberculose est régie par l arrêté royal du 17 octobre 2002.
 Le diagnostic de la tuberculose bovine La lutte contre la tuberculose est régie par l arrêté royal du 17 octobre 2002. 1. Tuberculination Dans la première phase d une infection de tuberculose bovine (Mycobacterium
Le diagnostic de la tuberculose bovine La lutte contre la tuberculose est régie par l arrêté royal du 17 octobre 2002. 1. Tuberculination Dans la première phase d une infection de tuberculose bovine (Mycobacterium
Mécanismes de l alloréactivité, des rejets de greffe et de la réaction du greffon contre l hôte.
 Mécanismes de l alloréactivité, des rejets de greffe et de la réaction du greffon contre l hôte. Marcelo de Carvalho Bittencourt, Christophe Baron, Gilles Blancho, Myriam Labalette, Hélène Moins Teisserenc
Mécanismes de l alloréactivité, des rejets de greffe et de la réaction du greffon contre l hôte. Marcelo de Carvalho Bittencourt, Christophe Baron, Gilles Blancho, Myriam Labalette, Hélène Moins Teisserenc
vaccin pneumococcique polyosidique conjugué (13-valent, adsorbé)
") EMA/90006/2015 EMEA/H/C/001104 Résumé EPAR à l intention du public vaccin pneumococcique polyosidique conjugué (13-valent, adsorbé) Le présent document est un résumé du rapport européen public d évaluation
EMA/90006/2015 EMEA/H/C/001104 Résumé EPAR à l intention du public vaccin pneumococcique polyosidique conjugué (13-valent, adsorbé) Le présent document est un résumé du rapport européen public d évaluation
HEPATITES VIRALES 22/09/09. Infectieux. Mme Daumas
 HEPATITES VIRALES 22/09/09 Mme Daumas Infectieux Introduction I. Hépatite aigu II. Hépatite chronique III. Les différents types d hépatites A. Hépatite A 1. Prévention de la transmission 2. Vaccination
HEPATITES VIRALES 22/09/09 Mme Daumas Infectieux Introduction I. Hépatite aigu II. Hépatite chronique III. Les différents types d hépatites A. Hépatite A 1. Prévention de la transmission 2. Vaccination
Les OGM. 5 décembre 2008. Nicole Mounier
 Les OGM 5 décembre 2008 Nicole Mounier Université Claude Bernard Lyon 1 CGMC, bâtiment Gregor Mendel 43, boulevard du 11 Novembre 1918 69622 Villeurbanne Cedex OGM Organismes Génétiquement Modifiés Transfert
Les OGM 5 décembre 2008 Nicole Mounier Université Claude Bernard Lyon 1 CGMC, bâtiment Gregor Mendel 43, boulevard du 11 Novembre 1918 69622 Villeurbanne Cedex OGM Organismes Génétiquement Modifiés Transfert
Mutations du gène CARD15 et maladie de Crohn
 Mutations du gène CARD15 et maladie de Crohn J.-P. HUGOT (Paris) Tirés à part : Jean-Pierre Hugot Hôpital Robert Debré, 48, boulevard Serrurier, 75019 Paris. Le rôle du gène CARD15/NOD2 dans la maladie
Mutations du gène CARD15 et maladie de Crohn J.-P. HUGOT (Paris) Tirés à part : Jean-Pierre Hugot Hôpital Robert Debré, 48, boulevard Serrurier, 75019 Paris. Le rôle du gène CARD15/NOD2 dans la maladie
GRANULOMATOSE SEPTIQUE CHRONIQUE
 GRANULOMATOSE SEPTIQUE CHRONIQUE Le présent livret a été rédigé à l attention des patients et de leurs familles. Il ne doit pas remplacer les conseils d un spécialiste en immunologie. 1 Egalement Disponible:
GRANULOMATOSE SEPTIQUE CHRONIQUE Le présent livret a été rédigé à l attention des patients et de leurs familles. Il ne doit pas remplacer les conseils d un spécialiste en immunologie. 1 Egalement Disponible:
Mécanismes de Subversion de l Immunité Innée par le Virus de l Hépatite C (VHC)
") Université de Montréal Mécanismes de Subversion de l Immunité Innée par le Virus de l Hépatite C (VHC) par Loubna Jouan Département de sciences biomédicales Faculté de médecine Thèse présentée à la Faculté
Université de Montréal Mécanismes de Subversion de l Immunité Innée par le Virus de l Hépatite C (VHC) par Loubna Jouan Département de sciences biomédicales Faculté de médecine Thèse présentée à la Faculté
Le VIH et votre foie
 Le VIH et votre foie Le VIH et votre foie Que dois-je savoir au sujet de mon foie? Votre foie joue un rôle incroyablement important. Il filtre votre sang en éliminant les substances nocives (toxiques)
Le VIH et votre foie Le VIH et votre foie Que dois-je savoir au sujet de mon foie? Votre foie joue un rôle incroyablement important. Il filtre votre sang en éliminant les substances nocives (toxiques)
MODULE DE VIROLOGIE 2010 IMMUNITE INNEE ANTIVIRALE CHEZ LES VERTEBRES
 MODULE DE VIROLOGIE 2010 IMMUNITE INNEE ANTIVIRALE CHEZ LES VERTEBRES Marc DALOD CIML Téléphone 04-91-26-94-51 email: dalod@ciml.univ-mrs.fr INTRODUCTION BRAINSTORMING PAR PETITS GROUPES (10 MIN) 1) Qu
MODULE DE VIROLOGIE 2010 IMMUNITE INNEE ANTIVIRALE CHEZ LES VERTEBRES Marc DALOD CIML Téléphone 04-91-26-94-51 email: dalod@ciml.univ-mrs.fr INTRODUCTION BRAINSTORMING PAR PETITS GROUPES (10 MIN) 1) Qu
Information génétique
 chapitre 3 Information génétique et division cellulaire L étude de la division cellulaire est abordée pour découvrir comment est transmise et conservée l information génétique portée par les chromosomes.
chapitre 3 Information génétique et division cellulaire L étude de la division cellulaire est abordée pour découvrir comment est transmise et conservée l information génétique portée par les chromosomes.
MICROBIOLOGIE. 1. Strep A et Urine Slide. 2. Coloration de Gram 3. Virologie (HCV, HBV, HIV)
") MICROBIOLOGIE 1. Strep A et Urine Slide (page 1) 2. Coloration de Gram (page 1) 3. Virologie (HCV, HBV, HIV) (page 1) 4. UKNEQAS Microbiology (page 2) 5. QCMD (page 4) Les programmes 1, 2 et 3 sont organisés
MICROBIOLOGIE 1. Strep A et Urine Slide (page 1) 2. Coloration de Gram (page 1) 3. Virologie (HCV, HBV, HIV) (page 1) 4. UKNEQAS Microbiology (page 2) 5. QCMD (page 4) Les programmes 1, 2 et 3 sont organisés
Exposé sur la Transfusion Sanguine
 Sommaire : I Histoire de la transfusion sanguine A/ Découverte des groupes sanguins B/ Premières transfusions sanguines C/ Les journées du sang II Les groupes sanguins A/ Les différents groupes B/ Déterminer
Sommaire : I Histoire de la transfusion sanguine A/ Découverte des groupes sanguins B/ Premières transfusions sanguines C/ Les journées du sang II Les groupes sanguins A/ Les différents groupes B/ Déterminer
Y a-t-il une place pour un vaccin thérapeutique contre l hépatite B? H. Fontaine, Unité d Hépatologie Médicale, Hôpital Cochin
 Y a-t-il une place pour un vaccin thérapeutique contre l hépatite B? H. Fontaine, Unité d Hépatologie Médicale, Hôpital Cochin Objectifs thérapeutiques Inhibition de la réplication virale Négativation
Y a-t-il une place pour un vaccin thérapeutique contre l hépatite B? H. Fontaine, Unité d Hépatologie Médicale, Hôpital Cochin Objectifs thérapeutiques Inhibition de la réplication virale Négativation
Contrôle de l'expression génétique : Les régulations post-transcriptionnelles
 Contrôle de l'expression génétique : Les régulations post-transcriptionnelles http://perso.univ-rennes1.fr/serge.hardy/ utilisateur : biochimie mot de passe : 2007 L'ARNm, simple intermédiaire entre le
Contrôle de l'expression génétique : Les régulations post-transcriptionnelles http://perso.univ-rennes1.fr/serge.hardy/ utilisateur : biochimie mot de passe : 2007 L'ARNm, simple intermédiaire entre le
TD de Biochimie 4 : Coloration.
 TD de Biochimie 4 : Coloration. Synthèse de l expérience 2 Les questions posées durant l expérience 2 Exposé sur les méthodes de coloration des molécules : Générique Spécifique Autres Questions Pourquoi
TD de Biochimie 4 : Coloration. Synthèse de l expérience 2 Les questions posées durant l expérience 2 Exposé sur les méthodes de coloration des molécules : Générique Spécifique Autres Questions Pourquoi
Laurence LEGOUT, Michel VALETTE, Henri MIGAUD, Luc DUBREUIL, Yazdan YAZDANPANAH et Eric SENNEVILLE
 Laurence LEGOUT, Michel VALETTE, Henri MIGAUD, Luc DUBREUIL, Yazdan YAZDANPANAH et Eric SENNEVILLE Service Régional Universitaires des Maladies Infectieuses et du Voyageur, Hôpital Gustave Dron 59208 TOURCOING
Laurence LEGOUT, Michel VALETTE, Henri MIGAUD, Luc DUBREUIL, Yazdan YAZDANPANAH et Eric SENNEVILLE Service Régional Universitaires des Maladies Infectieuses et du Voyageur, Hôpital Gustave Dron 59208 TOURCOING
La Greffe de Cellules Souches Hématopoïétiques
 La Greffe de Cellules Souches Hématopoïétiques Professeur Ibrahim Yakoub-Agha CHRU de LILLE (Illustration de J. Cloup, extraite du CD-Rom «greffe de Moelle» réalisé par la société K Noë) La moelle osseuse
La Greffe de Cellules Souches Hématopoïétiques Professeur Ibrahim Yakoub-Agha CHRU de LILLE (Illustration de J. Cloup, extraite du CD-Rom «greffe de Moelle» réalisé par la société K Noë) La moelle osseuse
Biomarqueurs en Cancérologie
 Biomarqueurs en Cancérologie Définition, détermination, usage Biomarqueurs et Cancer: définition Anomalie(s) quantitative(s) ou qualitative(s) Indicative(s) ou caractéristique(s) d un cancer ou de certaines
Biomarqueurs en Cancérologie Définition, détermination, usage Biomarqueurs et Cancer: définition Anomalie(s) quantitative(s) ou qualitative(s) Indicative(s) ou caractéristique(s) d un cancer ou de certaines
Hépatite B. Le virus Structure et caractéristiques 07/02/2013
 Hépatite B Le virus Structure et caractéristiques o o o Famille des Hepadnaviridae Genre orthohepadnavirus Enveloppé, capside icosaédrique, 42 nm 1 Le virus Structure et caractéristiques En microscopie
Hépatite B Le virus Structure et caractéristiques o o o Famille des Hepadnaviridae Genre orthohepadnavirus Enveloppé, capside icosaédrique, 42 nm 1 Le virus Structure et caractéristiques En microscopie
interféron ron β 1a dans le traitement des formes rémittentes de sclérose en plaques
 Le mécanisme m d action d et les effets de l interfl interféron ron β 1a dans le traitement des formes rémittentes de sclérose en plaques Présentation du domaine de terminologie Fanny Jarnet Le 13 avril
Le mécanisme m d action d et les effets de l interfl interféron ron β 1a dans le traitement des formes rémittentes de sclérose en plaques Présentation du domaine de terminologie Fanny Jarnet Le 13 avril
LA STOMATITE LYMPHOPLASMOCYTAIRE FELINE
 ANNEE 2006 THESE : 06 TOU 3 4103 LA STOMATITE LYMPHOPLASMOCYTAIRE FELINE MECANISMES IMMUNOPATHOLOGIQUES ET APPLICATIONS THERAPEUTIQUES THESE pour obtenir le grade de DOCTEUR VETERINAIRE DIPLOME D ETAT
ANNEE 2006 THESE : 06 TOU 3 4103 LA STOMATITE LYMPHOPLASMOCYTAIRE FELINE MECANISMES IMMUNOPATHOLOGIQUES ET APPLICATIONS THERAPEUTIQUES THESE pour obtenir le grade de DOCTEUR VETERINAIRE DIPLOME D ETAT
Notre système. Immunitaire
 Notre système Immunitaire Notre système Immunitaire Edito L association I.R.I.S. salue l initiative de Sara Lebien, qui en écrivant cet ouvrage et en proposant une illustration gaie et coloré a rendu l
Notre système Immunitaire Notre système Immunitaire Edito L association I.R.I.S. salue l initiative de Sara Lebien, qui en écrivant cet ouvrage et en proposant une illustration gaie et coloré a rendu l
L AUTOGREFFE QUELQUES EXPLICATIONS
 L AUTOGREFFE QUELQUES EXPLICATIONS Le traitement de votre maladie nécessite une Chimiothérapie intensive. Cette chimiothérapie qui utilise de fortes doses de médicaments antimitotiques est plus efficace
L AUTOGREFFE QUELQUES EXPLICATIONS Le traitement de votre maladie nécessite une Chimiothérapie intensive. Cette chimiothérapie qui utilise de fortes doses de médicaments antimitotiques est plus efficace
Vue d ensemble : Office of Cellular, Tissue and Gene Therapies
 Vue d ensemble : Office of Cellular, Tissue and Gene Therapies DIAPOSITIVE 1 Cette présentation fournit une vue d ensemble de l Office of Cellular, Tissue, and Gene Therapies (bureau des thérapies cellulaires,
Vue d ensemble : Office of Cellular, Tissue and Gene Therapies DIAPOSITIVE 1 Cette présentation fournit une vue d ensemble de l Office of Cellular, Tissue, and Gene Therapies (bureau des thérapies cellulaires,
82 collaborateurs 52 millions
 Édito Chiffre d affaires à fin Septembre 2007 Assemblée générale du 26 juin 2007 Réponses aux questions des actionnaires * Actualité R&D et produits I 2 : la science d Innate * Innate Pharma et ses actionnaires
Édito Chiffre d affaires à fin Septembre 2007 Assemblée générale du 26 juin 2007 Réponses aux questions des actionnaires * Actualité R&D et produits I 2 : la science d Innate * Innate Pharma et ses actionnaires
Communiqué. Abbott présente à Santé Canada une demande d homologation d HUMIRA pour le traitement du psoriasis POUR PUBLICATION IMMÉDIATE
 POUR PUBLICATION IMMÉDIATE Communiqué Abbott présente à Santé Canada une demande d homologation d HUMIRA pour le traitement du psoriasis Media: Sylvie Légaré (514) 832-7268 Abbott Gabrielle Collu 514)
POUR PUBLICATION IMMÉDIATE Communiqué Abbott présente à Santé Canada une demande d homologation d HUMIRA pour le traitement du psoriasis Media: Sylvie Légaré (514) 832-7268 Abbott Gabrielle Collu 514)
Risque infectieux et protection de l organisme
 Risque infectieux et protection de l organisme 1 La menace microbienne Comment définir les microbes? Activité 1 Les micro-organismes qui nous entourent Q1. Observer la préparation du bacille de Koch ou
Risque infectieux et protection de l organisme 1 La menace microbienne Comment définir les microbes? Activité 1 Les micro-organismes qui nous entourent Q1. Observer la préparation du bacille de Koch ou
Enseignement de Virologie (part. 2) Pr. Y. BAKRI Plan du cours
 Pr. Y. BAKRI Plan du cours") Université Mohammed V-Agdal Département de Biologie Faculté des Sciences Filière SVI Semestre 4 Année Universitaire : 2009-2010 Module optionnel: Biologie Humaine (M 16.1) Elément : Parasitologie - Virologie
Université Mohammed V-Agdal Département de Biologie Faculté des Sciences Filière SVI Semestre 4 Année Universitaire : 2009-2010 Module optionnel: Biologie Humaine (M 16.1) Elément : Parasitologie - Virologie
Charges virales basses sous traitement: définition impact virologique. Laurence Bocket Virologie CHRU de Lille
 XVIIe Journée Régionale de Pathologie Infectieuse 12 octobre 2010 Charges virales basses sous traitement: définition impact virologique Laurence Bocket Virologie CHRU de Lille conflits d intérêts subventions,
XVIIe Journée Régionale de Pathologie Infectieuse 12 octobre 2010 Charges virales basses sous traitement: définition impact virologique Laurence Bocket Virologie CHRU de Lille conflits d intérêts subventions,
Chapitre II La régulation de la glycémie
 Chapitre II La régulation de la glycémie Glycémie : concentration de glucose dans le sang valeur proche de 1g/L Hypoglycémie : perte de connaissance, troubles de la vue, voire coma. Hyperglycémie chronique
Chapitre II La régulation de la glycémie Glycémie : concentration de glucose dans le sang valeur proche de 1g/L Hypoglycémie : perte de connaissance, troubles de la vue, voire coma. Hyperglycémie chronique
Annales du Contrôle National de Qualité des Analyses de Biologie Médicale
 Annales du Contrôle National de Qualité des Analyses de Biologie Médicale ARN du virus de l hépatite C : ARN-VHC ARN-VHC 03VHC1 Novembre 2003 Edité : mars 2006 Annales ARN-VHC 03VHC1 1 / 8 ARN-VHC 03VHC1
Annales du Contrôle National de Qualité des Analyses de Biologie Médicale ARN du virus de l hépatite C : ARN-VHC ARN-VHC 03VHC1 Novembre 2003 Edité : mars 2006 Annales ARN-VHC 03VHC1 1 / 8 ARN-VHC 03VHC1
Université de Provence Aix-Marseille I
 Université de Provence Aix-Marseille I /_/_/_/_/_/_/_/_/_/_/ École Doctorale des Sciences Chimiques de Marseille THÈSE Présentée et soutenue publiquement le 12 décembre 2011 par : Nadège HANDKÉ Ingénieur
Université de Provence Aix-Marseille I /_/_/_/_/_/_/_/_/_/_/ École Doctorale des Sciences Chimiques de Marseille THÈSE Présentée et soutenue publiquement le 12 décembre 2011 par : Nadège HANDKÉ Ingénieur
Le don de moelle osseuse :
 DON DE MOELLE OSSEUSE Le don de moelle osseuse : se décider aujourd hui, s engager pour longtemps LA MOELLE OSSEUSE ET SA GREFFE La moelle osseuse C est le tissu mou dans le centre du corps des os qui
DON DE MOELLE OSSEUSE Le don de moelle osseuse : se décider aujourd hui, s engager pour longtemps LA MOELLE OSSEUSE ET SA GREFFE La moelle osseuse C est le tissu mou dans le centre du corps des os qui
THÈSE. Docteur. l Institut des Sciences et Industries du Vivant et de l Environnement (Agro Paris Tech) Leïla Périé. le 14/10/2009
 Leïla Périé. le 14/10/2009") THÈSE pour obtenir le grade de Docteur de l Institut des Sciences et Industries du Vivant et de l Environnement (Agro Paris Tech) Spécialité : Immunologie présentée et soutenue publiquement par Leïla Périé
THÈSE pour obtenir le grade de Docteur de l Institut des Sciences et Industries du Vivant et de l Environnement (Agro Paris Tech) Spécialité : Immunologie présentée et soutenue publiquement par Leïla Périé
VACCINS ANTIPNEUMOCOCCIQUES
 VACCINS ANTIPNEUMOCOCCIQUES Infections respiratoires SPIF - 15 Novembre 2014 Patrick Petitpretz Déclaration de liens d'intérêts J ai actuellement, ou j ai eu au cours des trois dernières années, une affiliation
VACCINS ANTIPNEUMOCOCCIQUES Infections respiratoires SPIF - 15 Novembre 2014 Patrick Petitpretz Déclaration de liens d'intérêts J ai actuellement, ou j ai eu au cours des trois dernières années, une affiliation
TITRE : On est tous séropositif!
 PRÉPARER UNE «SITUATION COMPLEXE DISCIPLINAIRE», SITUATION D APPRENTISSAGE ET/OU D ÉVALUATION TITRE : On est tous séropositif! classe : 3 ème durée : 30 min I - Choisir des objectifs dans les textes officiels.
PRÉPARER UNE «SITUATION COMPLEXE DISCIPLINAIRE», SITUATION D APPRENTISSAGE ET/OU D ÉVALUATION TITRE : On est tous séropositif! classe : 3 ème durée : 30 min I - Choisir des objectifs dans les textes officiels.
Subversion de l immunité innée et adaptative par les virus. Philippe Sansonetti Leçon #5 Collège de France 26 Janvier 2012
 Subversion de l immunité innée et adaptative par les virus Philippe Sansonetti Leçon #5 Collège de France 26 Janvier 2012 Des virus et des hommes... Les virus sont les pathogènes les plus divers, les plus
Subversion de l immunité innée et adaptative par les virus Philippe Sansonetti Leçon #5 Collège de France 26 Janvier 2012 Des virus et des hommes... Les virus sont les pathogènes les plus divers, les plus
Les interférons de type I et leur fonction antivirale
 Manuscrit déposé le 09/01/2006 Ann. Méd. Vét., 2006, 150, 73-107 formation continue - articles de synthèse Les interférons de type I et leur fonction antivirale LEROY M., DESMECHT D. Département de Morphologie
Manuscrit déposé le 09/01/2006 Ann. Méd. Vét., 2006, 150, 73-107 formation continue - articles de synthèse Les interférons de type I et leur fonction antivirale LEROY M., DESMECHT D. Département de Morphologie
MISSION PARTENARIALE IMMUNOLOGIE - VACCINATION & INFECTIOLOGIE A l occasion de BIOPHARM AMERICA 2015 BOSTON, ETATS-UNIS 14 au 18 septembre 2015
 SAVE THE DATE! MISSION PARTENARIALE IMMUNOLOGIE - VACCINATION & INFECTIOLOGIE A l occasion de BIOPHARM AMERICA 2015 BOSTON, ETATS-UNIS 14 au 18 2015 BioPharm America est le plus grand événement de partenariats
SAVE THE DATE! MISSION PARTENARIALE IMMUNOLOGIE - VACCINATION & INFECTIOLOGIE A l occasion de BIOPHARM AMERICA 2015 BOSTON, ETATS-UNIS 14 au 18 2015 BioPharm America est le plus grand événement de partenariats
La résistance d'agents infectieux aux médicaments antimicrobiens
 DECLARATION COMMUNE DES ACADEMIES DU G SCIENCE 2013 La résistance d'agents infectieux aux médicaments antimicrobiens Une menace globale pour l'humanité Depuis l introduction dans les années 40 du premier
DECLARATION COMMUNE DES ACADEMIES DU G SCIENCE 2013 La résistance d'agents infectieux aux médicaments antimicrobiens Une menace globale pour l'humanité Depuis l introduction dans les années 40 du premier