BIOLOGIE MOLECULAIRE. 1 ère ANNEE PHARMACIE. Pr. Alami Ouahabi Naïma
|
|
|
- Valérie Sénéchal
- il y a 7 ans
- Total affichages :
Transcription
1 BIOLOGIE MOLECULAIRE 1 ère ANNEE PHARMACIE 2012 Pr. Alami Ouahabi Naïma 1
2 PLAN DU COURS Chapitre.1 Structure des acides nucléiques & relations structure / fonction. Chapitre. 2 Expression génétique & systèmes de régulation. Transcription & traduction Cycle cellulaire & réplication de l ADN Chapitre. 3 Systèmes de mutation & de réparation de l ADN in vivo.
3 Chapitre. 4 Biologie moléculaire du cancer (Proto-oncogènes, oncogènes, apoptose & agents anti-cancer). Chapitre. 5 Biologie moléculaire des virus, viroïdes & prions (Analyse moléculaire & moyens génétique de diagnostique & de traitement). Chapitre. 6 Outils, techniques & applications de biologie moléculaire.
4 OBJECTIFS DU COURS : Partie Biologie Moléculaire Transmettre à l étudiant : 1. Les bases essentielles de la Biologie Moléculaire. 2. Une compréhension spécifique des mécanismes moléculaires clef & communs à différent organismes. 3. La capacité d analyser ces différents processus moléculaires.
5 4. La capacité de distinguer les processus moléculaires ciblés à des fins thérapeutiques. 5. La capacité de sélectionner les outils moléculaires nécessaires à ce ciblage thérapeutique. 6. Une compréhension globale de l importance des techniques de Biologie moléculaire dans la pharmacologie, le diagnostique ainsi que la thérapie présente & future.
6 Chapitre. 1. Structure des acides nucléiques & relations structure / fonction. Objectifs: 1. Connaître la structure des acides nucléiques; 2. Savoir reconnaître les molécules simples dont ils sont constitués; 3. Connaître leurs fonctions dans l expression génétique.
7 I. Structure des acides nucléiques Les acides nucléiques ont été isolés initialement des noyaux des cellules. On peut en distinguer deux grands types: les acides désoxyribonucléiques (ADN): essentiellement localisés dans le noyau des cellules les acides ribonucléiques (ARN): essentiellement localisés dans le cytoplasme cellulaire. Ces molécules biologiques (les acides nucléiques) contiennent l information génétique. Les acides nucléiques (ADN et ARN) sont des macromolécules composés de molécules simples et comportent des sous-unités appelées nucléotides. Un nucléotide comporte trois composants: un ose, une base & de l acide phosphorique.
8 1. 1. Ribose, désoxyribose
9 Bases puriques
10 Bases pyrimidiques
11 Nucléosides
12 Nucléotide AMP
13 Nucléotide GMP
14 Nucléotide CMP
15 Nucléotide UMP
16 Les Acides Nucléiques Acide ribonucléique
17 Hybridation A - T
18 Hybridation G - C
19 Les polymères de nucléotides: La molécule d ADN est constituée en règle de deux chaînes (ou brins) de nucléotides. Les molécules d ARN sont le plus souvent sous forme d un seul brin
20 Structure de l ARN en forme d Epingle à cheveux (hair-pin loops) Les nucléotides des acides ribonucléiques peuvent quelquefois s autohybrider en formant des structures secondaires
21 Acide désoxyribonucléique
22 La double hélice Dans l espace les deux brin d ADN antiparallèles & complémentaire présentent une configuration hélicoïdale. Elles s enroulent autour d un axe imaginaire pour constituer une double hélice à rotation droite (dans les formes A et B de l ADN)
23 Sens de lecture d un acide nucléique. Par convention, on lit toujours un acide nucléique de l extrémité 5 (groupement phosphate) vers l extrémité 3 (OH libre). on indique seulement La séquence des bases d un ADN (A, T, G ou C), en précisant les extrémités 5 et 3. Les 4 nucléotides de l ADN avec les 4 bases (A, T, G ou C), se combinent en une infinité de séquences de la même manière que les 7 notes de musique composent une infinité de symphonies.
24 La forme B de l ADN La forme biologique la plus importante de l ADN; 10 paires de bases par tour de spire; Le pas de l hélice 3,4 nm; Le diamètre de l hélice est de 2,4 nm; Les bases puriques et pyrimidiques sont à l intérieur de l hélice Les groupements phosphates et les désoxyriboses sont à l extérieur; Deux types de sillons appelés : sillon majeur (1,2 nm de large) et sillon mineur (0,6 nm de large).
25 La forme Z de l ADN Double hélice à rotation gauche; 12 paires de bases par tour d hélice; Le pas d hélice est 4,6 nm; Diamètre de l hélice est plus petit 1,8 nm; Les bases sont enchaînées avec une alternance de conformation; Le Z-ADN a été décelé dans des chromosomes de mammifères; Fonction précise mal connue, (régulation structurale de l expression génétique).
26 Propriétés physico-chimiques de l ADN. A la température T m (de fusion), l ADN est dénaturé; La dénaturation est réversible dans certaines conditions; Elle entraîne des modifications physico-chimiques: Augmentation de l absorption dans l ultra-violet, diminution de la viscosité, augmentation de la densité. T m varie selon le pourcentage de bases (G+C) de l ADN étudié; La présence des bases puriques et pyrimidiques permet aux acides nucléiques (ADN et ARN) d absorber dans l ultra-violet (UV) à 260 nm.
27 I LA CONFORMATION DES ADN LES TOPOISOMÈRES. Les topoisomères sont deux molécules d ADN qui ont la même séquence & diffèrent uniquement par le nombre d enlacements ( superenroulement ou le nombre de tours que fait l un des brins autour de l autre brin). Il existe Différents états des topoisomères. L ADN peut exister: à l état relâché avec une contrainte minimale dans la molécule. C est la forme la plus stable de la molécule. Cependant, l axe de la double hélice d ADN peut s enrouler sur lui-même en formant un super enroulement.
28 Deux formes de superenroulement sont alors possibles: -Un superenroulement qui correspond à: une augmentation du nombre d enroulements dans la même direction que la rotation de l hélice B (rotation droite). On parle de superenroulement positif. -Un superenroulement de l ADN autour de son axe dans la direction opposée au sens des aiguilles d une montre. Il y a donc au niveau de l ADN relâchement de la pression de torsion. On parle de superenroulement négatif. Le superenroulement de l ADN a des conséquences importantes: - Il permet de le rendre plus compact et diminuer ainsi le volume occupé dans la cellule. - Ces modifications du degré d enroulement de la double hélice d ADN influencent les interactions de l ADN avec d autres molécules (protéines qui régulent l expression génétique).
29
30 LES TOPOISOMÉRASES Les topoisomérases sont des enzymes qui modifient le nombre d enlacements. Elles augmentent ou diminuent le nombre de supertours dans les molécules d ADN double brin. Les topoisomérases de type I: Coupent transitoirement et ressoudent un seul brin d ADN double brin. Ils peuvent agir sur de l ADN superenroulé positif ou négatif. Les topoisomérases II : coupent de manière transitoire les deux brins de l ADN, puis les ressoudent et agissent uniquement sur l ADN superenroulé négatif (exemple: gyrase bactérienne). Les topoisomérases I & II permettent de désenrouler (relacher) l ADN.
31 Rôle des topoisomérases Les deux types de topoisomérases ont une importance capitale dans la réplication, la transcription et la recombinaison de l ADN. chez les procaryotes et les eucaryotes. Ces enzymes sont également la cible d agents médicamenteux, soit par exemple : les agents anti-bactériens de la classe des quinolones qui inhibent les gyrases bactériennes ou les agents anti-cancéreux, (le taxol )qui agissent en tant que anti topo isomérases I. Exemple de poison anti topoisomérase I : ts HMG
32 STRUCTURE DE LA CHROMATINE Condensation de la chromatine dans les cellules somatiques la molécule d ADN nucléaire est fortement associée à des protéines pour constituer la chromatine. Les complexes protéines-adn sont appelés nucléosomes. Le nucléosome contient de l ADN enroulé autour d un «core» constitué de 8 protéines appelées histones 2x (H2A H2B, H3 et H4).
33 Les histones sont des protéines de petit poids moléculaire (11-14 k Da); Les histones sont riches en acides aminés basiques (+); L histone H1 n appartient pas au nucléosome, mais permet le contact entre deux nucléosomes. L enroulements successif de 6 nucléosomes forme des structures de type solénoïde. Les solénoïdes forment des domaines (boucles de solénoïdes) de 60 kb ( pb). Les boucles de solénoïdes s enroulent ou s empilent afin de former une mini bande chromosomale Les mini bandes s empilent pour former un chromosome
34 Chez l homme : pb, gènes, 46 chromosomes (2 x 23) Le projet du Génome Humain le séquençage complet des pb en juin 2001
35 Condensation de la chromatine dans les spermatides la spermiogenèse (maturation de la spermatide en spermatozoïde); Changements morphologiques et physiologiques de la spermatide; s accompagnent de changements drastiques au niveau nucléaire; Les histones somatiques sont remplacées par des protéines tshmg et des protamines; Changement de la structure de la chromatine; Augmentation de la condensation de l ADN spermatique; Inhibition de l expression génétique transcription & traduction; Autre model de la structure de la chromatine; Assure transport sans danger du patrimoine génétique vers l ovule.
36 Les travaux de recherche in vitro effectués sur la protéine tshmg par N. Alami-Ouahabi & al, ont montré que cette nouvelle protéine nucléaire est impliquée dans: La condensation et superenroulement de l ADN; L inhibition de la transcription donc de l expression génétique & de l activité de division cellulaire; Ces travaux ont également montrés que tshmg agit comme agent anti topoisomerase I; Pourrait être un potentiel agent anticancer pour les traitements par Thérapie Génique.
37 LES TÉLOMÈRES. Les télomères constituent les extrémités des chromosomes eucaryotes. Ils sont formés par des séquences répétitives d ADN. A l extrémité 3 des chromosomes, on retrouve des copies répétées de séquences de type TTGGGG (retrouvées chez un protozoaire cilié: Tétrahymena) ou TTAGGG (retrouvées chez l Homme). A l extrémité 5, on a les séquences complémentaires riches en cytosine. Le problème majeur lors de la réplication de l ADN est l élimination potentielle de l amorce d ARN la plus externe ce qui pourrait entraîner un raccourcissement de l ADN à chaque cycle de réplication. La protection des extrémités des chromosomes des eucaryotes est assurée par une enzyme spécifique: la télomérase.
38 TÉLOMÈRASE La réplication de l ADN à l extrémité des chromosomes eucaryotes fait donc intervenir une enzyme particulière appelée télomérase. Cette enzyme ajoute des séquences spécifiques en 3 d un brin d ADN. La télomérase va se positionner à l extrémité 3 du brin d ADN. La télomérase possède une matrice qui lui est propre et qui est une matrice d ARN, elle va se fixer à l extrémité 3 : (3 brin parental + télomérase) Une répétition de motifs: TTGGGG est synthétisée. Elle constitue le télomère.
39 Pour synthétiser le brin télomère complémentaire riche en C, une primase interviend avec synthèse d une amorce d ARN par copie de l extrémité 3. Puis l ADN polymérase allonge la chaîne à partir de l amorce d ARN. Finalement, une ligase assure la soudure finale. L activité des télomérases est très réduite ou nulle dans les cellules normales en culture. Après plusieurs réplications, les chromosomes perdent leurs télomères et deviennent instables. Par contre dans les cellules cancéreuses, l activité des télomérases est augmentée. Les télomères ainsi que les gènes & les activités des télomérases ont un intérêt primordial dans la recherche des phénomènes moléculaires de vieillissement, de longévité & d immortalité cellulaire.
40 TERMES CLE EN BIOLOGIE MOLECULAIRE
41 I.6. PSEUDOGENES Certains gènes apparus par duplication au cours de l évolution ont subi des mutations génétiques (substitutions, insertions, délétions, etc...) qui empêchent la transcription ou la traduction; Exemple: (codon Stop prématuré) Il en résulte que même si la transcription conduit à un ARN messager, celui-ci ne peut être traduit Ce codon stop prématuré à transformé le gène fonctionnel en pseudo gène non traduit.

42
43 Au cours de l évolution des espèces, le génome s agrandit continuellement par suite de duplications de gènes et de transpositions. Les duplications: se produisent en tandem, une séquence se trouvant répétée deux fois dans le génome à la suite de la duplication. Cette duplication permet à chacune des copies de la séquence d évoluer pour son propre compte ce qui augmente les chances de produire des caractères nouveaux. Les transpositions: résultent de l incorporation d acides nucléiques synthétisés hors du génome : soit par incorporation du DNA d un virus, soit par transcription reverse d un RNA de la cellule ou d un virus, au moyen d une reverse-transcriptase qui produit un DNA complémentaire (cdna) et d une transposase qui l incorpore au génome de la cellule.
44 Conversion de gène: Les séquences de gènes peuvent être transformées par échange d exons complets (par une transposition à partir d un autre gène). La transposition d un exon peut ajouter un La transposition d un exon peut ajouter un domaine nouveau dans la structure d une protéine ou restaurer un domaine devenu non fonctionnel au cours de l évolution.
45 Les phénomènes de transformation de gènes en Pseudo gènes, de transpositions & de coversions de gènes Jouent un rôle primordial dans l évolution, l adaptation & la diversité génétique. effet positif sur l expression génétique. Peuvent avoir un effet néfaste sur la régulation de l expression génétique. En effet un transposon peut lors de son mouvement interrompre un gène codant pour un facteur important de la régulation du cycle cellulaire ou un facteur biochimique vital & peut ainsi causer des métastases ou des pathologies.
46 LES ACIDES RIBONUCLÉIQUES (OU ARN) CARACTÉRISTIQUES GÉNÉRALES Les ARN sont caractérisés par: -L ose= ribose: remplace le 2 -désoxyribose présent dans les ADN. -Les bases pyrimidiques et puriques rencontrées sont: A, C, G et U (cette dernière est remplacée par T dans les ADN). Une seule chaîne nucléotidique (au lieu de deux dans les ADN). Cette unique chaîne est plus courte que les chaînes d ADN. Les règles d appariement entre les bases complémentaires peuvent s observer dans les ARN. Dans certains cas: une même chaîne d ARN peut s auto hybrider où les portions bi caténaire ont un appariement suivant la règle: A apparié avec U (deux liaisons hydrogène) et C apparié avec G (trois liaisons hydrogène). Ainsi les régions appariées donnent des tiges & les régions non appariées donnent des boucles (structure en cruciforme).
47 LES DIFFÉRENTS TYPES D ARN. Les cellules contiennent essentiellement quatre types d ARN: - Les ARN ribosomiques (ou rarn). - Les ARN de transfert (ou tarn). - Les ARN messagers (ou marn). - Les ARN nucléaires de petite taille (ou snrna)(ou small nuclear RNA)(snRNA) Dans la cellule, les rarn sont les plus abondants (au moins 80% de l ensemble des ARN de la cellule) et les snrna les moins abondants.
48 LES ARN RIBOSOMIQUES. Ils constituent les ribosomes; Les ribosomes sont des organites intra-cellulaires situés dans le cytoplasme (également dans les mitochondries) et qui sont l usine de fabrication des protéines de la cellule. On peut distinguer les rarn des procaryotes (E. coli) et les rarn des eucaryotes. Les rarn des procaryotes (E. coli): constituent les ribosomes dits 70 S (S = unité de sédimentation ou Svedberg) qui sont formés de deux sous-unités, une grande sous-unité (50 S) et une petite sous-unité (30 S). Chaque sous-unité comporte des protéines dites protéines ribosomales (ou r-protéines) et des rarn.
49 Les rarn des eucaryotes. Chez les eucaryotes, les ribosomes sont plus gros (80 S) avec également une structure à deux sous-unités (40 S et 60 S). La sous-unité 60 S contient trois rarn différents (5 S, 5,8 S et 28 S), et la petite sous-unité 40 S ne contient qu un seul rarn 18 S). Le nombre de protéines ribosomales à l intérieur de chaque sous-unité est plus important que chez les procaryotes..
50 LES ARN DE TRANSFERT (tarn). Les tarn sont les vecteurs qui vont transférer les acides aminés jusqu à l usine ribosome où s effectue la synthèse protéique. Structure des tarn. Les tarn présentent bien entendu la structure générale des ARN. Mais ils possèdent en plus quelques particularités propres. Des bases inhabituelles.tout d abord, ils contiennent des nucléotides inhabituels par les bases qu ils renferment. Ainsi, l hypoxanthine, dont le nucléotide correspondant est l IMP (I = inosine monophosphate), mais aussi la thymine ou des bases méthylées. La structure spatiale des tarn. Les chaînes de tarn sont constituées d une centaine environ de nucléotides. Surtout, il existe des zones d appariement selon la règle de complémentarité et des zones appelées boucles sans appariement où sont présentes les bases inhabituelles. La forme générale est celle d un L. En structure spatiale, on présente souvent les tarn sous forme de trèfle ou un cruciforme.
51 Les sites importants dans les tarn. Plusieurs sites sont importants dans les tarn. Tout d abord: leur extrémité 3 -OH. Il existe trois nucléotides caractéristiques -C- C-A-3 -OH. C est par cette extrémité que sera fixé l acide aminé qui sera véhiculé par le tarn. Puis, l anticodon qui correspond à un groupe de trois nucléotides (ou triplet) situé sur une boucle du tarn. Ce triplet présente un rôle très important puisqu il doit s apparier avec le codon correspondant présent sur l ARN messager. Cet appariement entre l anti-codon et le codon se fait par des liaisons hydrogène et suivant les règles de complémentarité. Le codon et l anti-codon sont disposés de manière antiparalllèle.
52
53 LES ARN MESSAGERS (marn). Les ARN messagers (ou marn) constituent le support essentiel de l information génétique entre l ADN et le ribosome où s effectuera la synthèse protéique. Leur durée de vie est très courte à la différence des tarn par exemple. Chez les bactéries, la durée de vie d un marn est de quelques minutes environ. Les marn sont très rapidement synthétisés et dégradés. Ils sont formés d une seule chaîne de nucléotides avec les bases communes aux ARN: A, U, C et G. Cette chaîne comporte une succession de triplets nucléotidiques. Chaque triplet nucléotidique constitue un codon spécifique d un acide aminé donné.
54 Il existe naturellement une vingtaine d acides aminés et on dénombre 61 codons différents! Ceci signifie qu un acide aminé pourra être véhiculé par plusieurs tarn spécifiques (différent par leur anticodon). Enfin, l extrémité 5 des tarn comporte un groupement phosphate. Les tarn jouent un rôle capital dans la biosynthèse protéique.
55 LES PETITS ARN NUCLEAIRES. Les petits ARN nucléaires (ou snrna) sont présents dans le noyau des cellules et sont impliqués dans certaines étapes de la régulation post transcriptionnelle (épissage alternatif).
56 II. Relation entre les structures des acides nucléiques et leurs fonctions lord de l expression génétique Les différentes structures des acides nucléiques & des protéines associées à la chromatine ont une grande influence sur la régulation de l expression génétique. En effet: La structure de la double hélice d ADN, sa forme A, B ou Z, sa charge négative et son degré de superenroulement jouent un rôle Déterminant dans l accessibilité des gènes au facteurs d initiation, D activation ou d inhibition de la transcription. Donc la structure de l ADN est déjà en soi un facteur régulateur de l expression génétique. Les interactions ADN ADN, ADN ARN, ADN Protéines & Protéines----Protéines sont également des éléments clef de La régulation de l expression des gènes.
57 INFORMATION GENETIQUE Expression Génétique : Transcription &Traduction Réplication & Cycle Cellulaire

58 Définitions : La double hélice Expression Génétique
59 L acide désoxyribonucléique (ADN ou DNA), constitué de deux chaînes de nucléotides monophosphates, liés chacun par une liaison ester entre son carbone 3 et le carbone 5 du nucléotide suivant. Ces deux chaînes de nucléotides sont unies entre elles par des liaisons hydrogènes pour former un hybride en forme de double hélice dont les deux brins sont : antiparallèles & complémentaires. Chaque adénine (A) d un des deux brins est liée par deux liaisons hydrogène avec une thymine (T) de l autre brin, et chaque guanine (G) d un brin est liée par trois liaisons hydrogène avec une cytosine (C) de l autre brin. De sorte que si on désigne les nucléotides par les lettres A, C, G et T en fonction des bases azotées qu ils contiennent, on peut lire sur cette figure le texte suivant : AGAGTCGTCTCGAGTCA......TCTCAGCAGAGCTCAGT
60 Écrire à la suite les lettres A, C, G et T dans l ordre où on rencontre les nucléotides sur l un des brins de l ADN de l extrémité 5 à l extrémité 3 aboutit à un texte. Le texte ainsi transcrit contient toute l information nécessaire pour la synthèse d une protéine). C est pourquoi on appelle ce brin d ADN le brin «sens». L autre brin est le brin complémentaire ou brin «antisens». L ensemble de l information génétique transmise par chacun des parents à un enfant (génome haploïde) est contenue ainsi en 3 x 10 9 nucléotides. Ces 3 x 10 9 nucléotides d information génétique constituent des gènes = séquences codantes = exons = cadres ouvert de lecture (Open Reading Frames = ORF). Ces gènes sont copiés dans le noyau en trna ou rrna ou mrna = Transcription. Seuls les mrna sont traduit dans le cytoplasme en protéines = Traduction.

61 Gène
62 Pour permettre la biosynthèse d une protéine, il doit y avoir dans la structure de l ADN, une séquence de nucléotides qui constitue le gène de cette protéine. Il existe des séquences régulatrices = éléments régulateurs ou promoteur: généralement proximal en amont du gène Le promoteur peut être en aval ou distal du gène et ne devient proximal que lors de la transcription et ceci via des remaniements structuraux de la chromatine). Le promoteur est une séquence non-transcrite et non-traduite. Une région génique ou locus peut contenir une suite variable d exons (ORFs transcrites et traduites) et d introns ( sequences non-codantes) L ensemble des mécanismes qui à partir de la séquence du gène (ADN) conduisent à la production d un acide ribonucléique (ARN) est désigné sous le terme de Transcription ARNm proteine == traduction Transcription & traduction == expression génétique.
63 Brin sens & Brin antisens Tout le DNA n est pas transcrit seuls les gènes = exons = ORFs Sont transcrits. En plus même si l ADN est double brin seul un brin est transcrit ou copié en mrna. Le mrna est complémentaire & antiparallèle au brin d ADN transcrit (nommé: Brin antisens) Le mrna est identique à l autre brin non transcrit (nommé: Brin sens). 5.AGCTTAACGCGTA. 3 3.TCGAATTGCGCAT. 5 Transcription 5.AGCUUAACGCGUA. 3 Brin DNA sens= non transcrit=informatif. Brin de DNA antisens=transcrit. mrna

64 1.A. La transcription
65 Les nucléotides qui précèdent l exon contiennent de nombreuses séquences reconnues par des protéines nucléaires qui permettent le début (initiation) & la régulation de la transcription. C est le promoteur On distingue en particulier : vers (20 à 30 nucléotides en amont de l exon 1 en direction de l extrémité 5 du brin sens) une séquence TATATA appelée «boîte TATA», qui est spécifiquement reconnue par la protéine TFIID, cofacteur de la RNA-polymérase II ; vers -80 une séquence GGCCCATCCAT à la fois «boîte CAT ou CAAT» et «boîte GC», sur laquelle viennent se fixer d autres protéines de la transcription (CTF et Sp-1) ; de nombreuses autres séquences sont des sites de fixation des protéines régulatrices de la transcription. Par exemple, une séquence TRE (TGACTCA) au début de la troisième ligne de la sequence, sur laquelle vont se fixer des protéines de la famille AP-1 pour activer la transcription.

66 Cis- et trans-régulateurs
67 Les séquences de nucléotides du promoteur=(facteurs cisrégulateurs), sont reconnues par une classe de protéines spécifiques=(facteurs trans-régulateurs) dont la structure permet une liaison avec l ADN (DNA binding proteins). La structure secondaire et tertiaire des DNA binding proteins comprend des domaines spécifiques permettant la reconnaissance des séquences régulatrices du gène : - domaines en doigts de zinc/ structures riches en leucine en forme de «fermeture Eclair» (Leu zipper)/ homéodomaines, bhlh = région basique, hélice a, boucle, hélice a/ - domaines riches en glutamine/ domaines riches en proline/ - hélices a acides (Asp, Glu). Les séquences qui reconnaissent les DNA binding proteins (facteurs cis-régulateurs=promoteur) ont des effets sur la vitesse de la transcription: activateurs = (séquences enhancer) ou inhibiteurs = (sé-quences silencer). Leur liaison avec les protéines dépend le plus souvent de circonstances physiologiques qui induisent ou répriment l expression du gène.

68 Initiation de la transcription La RNA polymérase II est l enzyme de la transcription des gènes exprimés sous forme de protéines. Elle est présente dans tous les noyaux cellulaires. Sa masse moléculaire est de 500 kilo daltons pour 10 sous-unités. La transcription qu elle catalyse nécessite des ribonucléosides triphosphates comme substrats (ATP, CTP, GTP et UTP), plusieurs cofacteurs protéiniques (transcription factors) L énergie de la réaction est fournie par l hydrolyse des liaisons riches en énergie des nucléosides triphosphates (42 kj/mol).
69 Initiation de la transcription L initiation de la transcription sur un promoteur humain implique plusieurs facteurs de transcription, connus sous les noms de TFIIA, TFIIB, TFIID, TFIIE, TFIIF, TFIIH et TFIIJ en plus de la RNA polymérase II elle-même. Au centre de ce mécanisme initial, il y a l association de TBP, la sous-unité reconnaissant l ADN du facteur TFIID, avec la boîte TATA du promoteur. L adjonction successive de TFIIB et de RAP30, petite sous-unité de TFIIF, sont ensuite nécessaires pour la fixation de la polymérase et déterminent à la fois le brin transcrit et le sens de la transcription. A ce complexe ADN-TFIID-TFIIB-RAP30-polymérase II, s ajoutent encore successivement la grande sous-unité de TFIIF (RAP74), TFIIE et TFIIH. Alors la transcription peut commencer si les ribonucléotides substrats sont présents.
70 Le complexe d initiation de la transcription recouvre une séquence d environ 100 nucléotides (100pb avant Nt: +1) du brin antisens de l ADN, en provoquant l ouverture et le déroulement partiel de la double hélice à cet endroit. La transcription commence à environ 22 nucléotides de la boîte TATA en direction opposée à celle des éléments GC-CAAT et se poursuit en remontant le brin antisens =(transcrit), en direction de son extrémité 5. La formation de ce complexe est activée ou inhibée par les facteurs trans-régulateurs liés aux séquences cisrégulatrices du même promoteur.

71

72 Elongation de la transcription
73 Chaque nucléoside triphosphate est choisi spécifiquement pour être complémentaire de la base du brin antisens qui va lui faire face. La liaison riche en énergie entre le premier phosphate estérifiant le carbone 5 du ribose et les deux autres phosphates est hydrolysée, libérant un pyrophosphate qui sera hydrolysé ensuite par une pyrophosphatase. Le phosphate restant est lié par une liaison ester au carbone 3 OH libre du dernier nucléotide du transcrit en cours de synthèse. La RNA polymérase construit un ARN hybridé avec le brin antisens de l ADN, dont la séquence primaire est la copie du brin sens mais composée de ribonucléotides au lieu des désoxyribonucléotides et d uracile à la place des thymines.
74
75
76
77
78 Exon
79 Les exons sont des fragments de séquence primaire d un gène qui seront recopiés dans la structure primaire de l ARN messager, après l épissage. Un exon code le plus souvent pour un domaine fonctionnel de la protéine ou une partie d un tel domaine. Au cours de l évolution le gène peut être modifié par la substitution, l insertion ou la délétion d un exon entier (conversion de gènes) ce qui modifie, apporte ou retire à la protéine un domaine fonctionnel en entier.
80 Exon 1
81 Après la fixation de la RNA-polymérase sur la boîte TATA par l intermédiaire du facteur TFIID, la transcription va commencer 23 nucléotides au delà de cette boîte. A ce point, la séquence du brin sens est 5 AGGCACAGA...3, celle du brin antisens est donc 3 TCCGTGTCT...5 (complémentaire et antiparallèle). En choisissant comme substrats les ribonucléotides complémentaires de ce brin antisens, la RNA-polymérase va composer 5 AGGCACAGA...3 qui sera le début de la séquence de l ARN transcrit. Lorsque la polymérase rencontre un A sur le brin antisens, elle incorpore un nucléotide à Uracile dans le mrna : 5 UGGCUCUGU...3. La séquence de l ARN transcrit recopie donc celle du brin sens de l ADN. La transcription se poursuit alors tout au long du brin antisens en incorporant les nucléotides dans l ARN transcrit.

82 Fin de la transcription
83
84
85 Vers la fin du gène la RNA-polymérase rencontre une séquence 3 -TTATTT-5 qu elle reconnaît comme un signal de fin d activité. La transcription s arrête en effet peu après ce signal qui est transcrit en AAUAAA. La RNA-polymérase quitte l ADN et libère le transcrit d ARN qui contient l information génétique recopiée pour permettre l expression du gène sous forme de protéine.

86 Modifications du transcrit
87 Chapeau = Capping: une enzyme ajoute un nucléotide à Guanine sur les phosphates du Carbone 5 du premier nucléotide du transcrit et transfère des radicaux méthyles sur les premiers nucléotides. Ces modifications font un début commun à tous les transcrits primaires, précurseurs des RNA messagers. Queue poly-a : une enzyme coupe le transcrit environ 10 à 20 nucléotides au delà de la séquence AAUAAA et synthétise sur le Carbone 3 libre du dernier nucléotide du transcrit restant une longue chaîne de 500 à 2000 nucléotides à Adénine polymérisés. Édition : Des enzymes peuvent «éditer» (au sens anglais du terme) la structure primaire du transcrit en modifiant certaines bases (exemple : C en U par une cytosine désaminase spécifique Excision - épissage : les parties non codantes de la structure primaire du transcrit sont coupées et les parties codantes ré assemblées
88 Les ARN messagers sont détruits par des ribonucléases dans le cytoplasme. Leur hydrolyse libère des nucléotides qui seront réemployés à la synthèse de nouveaux acides nucléiques. Pour protéger les ARN messagers de ce catabolisme, une structure particulière dissimule l extrémité 5 terminale de ces ARN aux exonucléases spécifiques de cette partie de l ARN. Cette structure est appelée chapeau du messager (cap). Au minimum, il y a fixation d un GMP sur le deuxième phosphate du nucléoside triphosphate qui se trouve à l extrémité 5 du transcrit. Ce GMP est méthylé sur son azote n 7. Si le nucléotide initial comprend une adénine celle-ci peut être méthylée sur l azote n 6. Les riboses des nucléotides initiaux sont quelquefois méthylés sur l oxygène de la fonction alcool en 2.
89 Le transcrit primaire contient des nucléotides. Il a reçu un chapeau (cap), GMP méthylé qui se fixe sur le phosphate b de l ATP en 5 du transcrit primaire. Il reçoit une queue poly-a synthétisée en 3 après la fin du transcrit. les introns excisés seront reconnus par : un site donneur GU en 5 de chaque intron, un site receveur AG en 3 de chaque intron. Après cette maturation le transcrit primaire devenu ARN messager sera transporté vers le cytoplasme pour la traduction.

90 Excision - épissage
91 Les introns, parties non codantes des gènes des eucaryotes, sont coupés (excision) de la structure primaire des ARN au cours de la maturation des messagers. Les exons, parties codantes, sont ensuite liés entre eux bout à bout (épissage), pour établir la séquence primaire de l ARN messager. L excision et l épissage représentent donc l action d enzymes et de ribozymes qui catalysent la coupure de l ARN (endoribonucléase) et la fermeture de la brèche (ARN ligase). L épissage peut être alternatif, c est à dire peut conduire à plusieurs structures d ARNm : les ARNm alternatifs ont chacun en propre certains exons ainsi que des exons en commun.

92 Formation du lasso
93 L excision des introns et l épissage des exons est l étape la plus importante de la maturation du transcrit dans le noyau cellulaire. Elle fait appel à des facteurs spécifiques : ribonucléoprotéines contenant des petits ARN (snr-np), ribozymes (ARN ayant des propriétés catalytiques), protéines codées par les introns eux-mêmes (maturases)... La structure secondaire du transcrit met en contact trois séquences de l intron : la séquence du début de l intron (extrémité 5 ou site donneur), la séquence de la fin de l intron (extrémité 3 ou site accepteur) et une séquence riche en pyrimidines (C ou U) contenant un nucléotide à adénine (A du branchement). Le site donneur commence habituellement par un nucléotide à guanine dont le phosphate est détaché de l exon précédent pour être transféré sur le carbone 2 de l A du branchement. Le dernier nucléotide du site accepteur, aussi une guanine, est détaché de l exon suivant, dont le premier nucléotide est lié par son phosphate 5 au carbone 3 du dernier nucléotide de l exon précédent. L intron, libéré sous forme de lasso, est détruit par des nucléases. Le transcrit perd successivement tous ses introns et les exons épissés constituent la séquence codante du messager.
94
95

96 RNA Messager
97 L ARN messager comprend plusieurs séquences : une séquence 5 non-codante, qui ne sera pas traduite, qui correspond à l exon 1 et à une partie de l exon 2 dans le gène de l apoa-ii ; un codon AUG qui fixe le tarn de la méthionine initiale ; des séquences de codons pour incorporer les acides aminés du signal-peptide et du propeptide; la séquence des codons dont les acides aminés formeront la séquence primaire de la protéine ; le codon de terminaison (ici, UGA) ; une séquence 3 non-traduite qui s achève par la queue poly-a. Sur la séquence 5 non-traduite, va se construire le complexe d initiation de la traduction où les ribosomes vont s assembler successivement pour commencer la traduction.
98

99
100

101 La traduction
102 Lors de la traduction: la structure transcrite sur l ARNm, s exprime par la synthèse de protéines dont la séquence traduit en acides aminés l information portée par la structure primaire de l ADN. La traduction est faite dans le cytoplasme des cellules : soit pour libérer des protéines cytoplasmiques, soit pour conduire ces protéines dans les membranes ou les organites de la cellule (reticulum endoplasmique, appareil de Golgi, membrane plasmique, lysosomes, mitochondries, noyau, etc...), soit pour excréter ces protéines à travers les membranes vers l extérieur de la cellule.

103 Codon
104 La liaison de l ARN de transfert (ARNt) avec l ARN messager (ARNm) porteur de l information, se fait par complémentarité entre 3 nucléotides de chacun de ces deux ARN. Les 3 nucléotides de l ARNm constituent un codon et les 3 nucléotides de l ARNt un anticodon. Au cours de la traduction l anticodon et le codon se lient de manière antiparallèle, et l acide aminé porté par l ARNt est incorporé à la protéine en cours de synthèse. La séquence primaire de l ARNm est donc traduite par groupes de 3 nucléotides (codons). Un nucléotide de l ARNm au cours de la traduction peut se trouver en position 1, 2 ou 3 dans un codon si le nombre de nucléotides qui le séparent du codon d initiation AUG est ou n est pas un multiple de 3. Cette position porte le nom de cadre de lecture.

105 Code génétique
106 La séquence codante est une suite de codons dont chacun permet d incorporer spécifiquement un acide aminé dans la synthèse d une protéine. Le code génétique est le même pour tous les êtres vivants de la biosphère (universel). Il existe quelques variations (codons propres à la biosynthèse des protéines dans les mitochondries). Le code génétique comporte 61 codons pour signifier les 20 acides aminés qui participent à la synthèse des protéines : chaque acide aminé peut être codé par plusieurs codons (de un à six) qui diffèrent en général par leur troisième nucléotide. On dit que le code est dégénéré.

107 Ribosome eucaryote
108 Les acides ribonucléiques ribosomiaux et les protéines ont des sites de fixation pour la séquence du messager, Les ARN de transfert portent l acide aminé à incorporer (site A) et le peptide en cours de synthèse (site P), un site catalytique pour former les liaisons peptidiques, des sites de fixation pour les cofacteurs protéiques de l initiation (eif2, eif3), de l élongation et de la terminaison et des sites de régulation (protéine S6).
109 Polyribosomes
110 L initiation est un phénomène permanent à l extrémité 5 d un ARN messager et les ribosomes se succèdent sur le même messager à raison d un tous les 100 nucléotides environ, formant un polyribosome. Les polyribosomes présentent un aspect différent selon la régulation des différentes étapes de la traduction. Plus l initiation est active plus les ribosomes sont nombreux sur le messager. Si l élongation est lente, les ribosomes vont mettre plus de temps à lire la séquence codante. Une activation de la terminaison dissocie tous les ribosomes du messager. De nombreux antibiotiques sont capables d interférer avec chacune des étapes de la synthèse des protéines (streptomycine, cycloheximide, puromycine).

111 ARN de transfert
112 Les ARN de transfert constituent le lien chimique nécessaire entre la structure du codon, reconnu par l anticodon, et l acide aminé spécifique porté par l ARNt. C est en quelque sorte le dictionnaire de la traduction. L anticodon est une séquence de trois nucléotides située à l extrémité de la boucle inférieure de l ARNt, complémentaire et antiparallèle de la séquence du codon de l acide aminé correspondant. Chaque acide aminé est lié spécifiquement (code génétique) par une aminoacyl-trna-synthétase à l extrémité 3 de l ARNt dont l anticodon lui correspond (ARNt chargé). Les ribosomes lient les ARNt chargés sur le site A de l élongation si leur anticodon s apparie avec le codon du messager à cet endroit. L élongation transfère alors le peptide sur l acide aminé nouveau, lui-même porté par l ARN de transfert.

113 Sérine trna synthétase
114 Cystéine trna synthétase
115 Les aminoacide-trna-synthétases sont les enzymes qui chargent les acides aminés libres du cytoplasme sur les ARN de transfert correspondants. Le coenzyme ATP est hydrolysé en AMP et en pyrophosphate pour fournir l énergie nécessaire. La liaison ester constituée entre l acide aminé et son ARNt est riche en énergie et sera hydrolysée au cours de l étape d élongation de la traduction. Les aminoacide-trna-synthétases ont une double spécificité très étroite pour les deux substrats : l acide aminé reconnu (ici, la cystéine) et les ARNt dont les anticodons sont complémentaires des codons correspondant à cet acide aminé dans le code génétique. L exactitude de la traduction repose entièrement sur cette double spécificité. La reconnaissance de l ARNt se fait par l anticodon dans certains cas (l enzyme ne tenant pas toujours compte de la première base de l anticodon, ce qui explique la dégénérescence du code génétique) ou bien encore par d autres séquences de l ARNt communes aux ARNt synonymes. Certaines aminoacide-trnasynthétases sont régulées par phosphorylation.

116 Initiation de la traduction
117 L initiation de la traduction est l étape limitante de la traduction. Les sous-unités des ribosomes sont dissociées dans le cytoplasme. Une cascade d évènements vont former un complexe d initiation. Le facteur eif2 (eucaryotic initiation factor 2) est porteur d un GDP, coenzyme qu il vient d hydrolyser à l étape précédente. En présence du facteur eif2b, un nouveau GTP est substitué à ce GDP. Le facteur eif2 ainsi activé, peut alors lier l ARNt chargé d une méthionine dont l anticodon est complémentaire du codon d initiation (AUG) du messager. En présence du cofacteur eif4c, la petite sous-unité va fixer le facteur eif3 et le facteur eif2 activé qui porte l ARNt chargé de la méthionine initiale. L énergie de la formation de ce complexe a été fournie par l hydrolyse de la liaison riche en énergie du GTP porté par le facteur eif2.
118 La séquence 5 non traduite de l ARN messager est reconnue par les cofacteurs eif4a, eif4b et eif4f qui s y fixent. Grâce à l hydrolyse d un ATP pour fournir l énergie, le messager est alors transféré sur la petite sous-unité, en regard du site P, de façon à hybrider les nucléotides du codon d initiation avec ceux de l anticodon de l ARNt de la méthionine initiale. En présence du dernier cofacteur eif5, le complexe va se lier à une grande sous-unité pour constituer un ribosome fonctionnel. Les cofacteurs d initiation sont libérés et la traduction commence. Le cofacteur eif2 toujours porteur de son GDP, se libère pour recommencer un nouveau complexe.
119 Cadre de lecture
120 Le positionnement du messager par rapport à l ARN de transfert de la Méthionine initiale détermine le cadre de lecture de la traduction du messager. L ARN de transfert de la méthionine occupe un des deux sites de fixation des trna sur le ribosome : site P (ou peptidique, car il contiendra le peptide en cours de synthèse). Le codon AUG du messager, en s hybridant dans le site P avec l anticodon CAU de l ARN de transfert de la méthionine, place le messager de telle sorte que le codon suivant (N1N2N3) apparaisse dans l autre site de fixation : site A (ou acide aminé, car il servira à la fixation des nouveaux acides aminés incorporés). Le nucléotide N1 sera toujours le premier de chaque codon.

121 Facteurs d initiation
122 La formation du complexe d initiation fait appel a de nombreux facteurs qui participent à la structure, à l activation ou à la régulation de ce complexe. Parmi ces facteurs on reconnaît les deux sous-unités des ribosomes (Unité L ou 60S et unité S ou 40S), le Met-tARN initiateur, l ARN messager et les coenzymes donneurs d énergie ATP et GTP. Les cofacteurs protéiniques de cette étape sont classifiés sur ce Tableau. Certains participent à l activation du Met-tARN initiateur ou à sa liaison avec la sous-unité S, d autres à la reconnaissance de l extrémité 5 de l ARN messager ou à sa liaison avec le complexe d initiation, d autres enfin à la liaison finale avec la sousunité L. Certains de ces facteurs protéiniques et une au moins des chaînes protéiniques de la petite sous-unité sont régulées par phosphorylation ou déphosphorylation sur des Ser ou des Thr.
123 Elongation

124 Incorporation d un acide aminé Un ARNt chargé vient se fixer sur le site A libre. Le codon du messager au fond du site A va se lier complémentairement avec l anticodon de l ARNt du nouvel acide aminé ce qui va permettre l incorporation de cet acide aminé dans le peptide en cours de synthèse.

125 Transfert du peptide Le ribosome catalyse alors le transfert du peptide situé sur l ARNt du site P sur la fonction amine de l acide aminé de l ARNt du site A. Il utilise pour cela, l énergie de l hydrolyse de la liaison ester riche en énergie entre le peptide et l ARNt du site P.
 du site A vers le site P, sans qu")
126 Translocation des codons L ARNt libre du site peptidique quitte le ribosome. Le messager, l ARNt restant et le peptide en cours de synthèse sont alors déplacés (translocation) du site A vers le site P, sans qu il y ait fusion de l hybride entre le codon et l anticodon
127 Terminaison de la traduction Lorsque le site A se trouve en regard d un codon nonsens annonçant la fin de la traduction, le complexe va se dissocier du messager en présence d un dernier cofacteur erf. Les deux sous-unités du ribosome se dissocient, la protéine synthétisée est libérée, ainsi que le dernier ARNt.
128
129 Facteurs d élongation ou de terminaison L élongation ou la terminaison de la protéine naissante font appel a de nombreux facteurs qui participent aux étapes successives de la traduction. Parmi ces facteurs on reconnaît les ribosomes, les aminoacyltarn activés, l ARN messager et le coenzyme donneur d énergie GTP. Les cofacteurs protéiniques de ces étapes sont classifiés sur Tableau. Certains participent à l activation des aminoacyl-tarn ou à leur liaison au site A du ribosome, d autres à la translocation du tarn porteur du peptide naissant du site A vers le site P. Le dernier facteur en-gendre la dissociation du ribosome et la libération de la protéine synthétisée.
130
131
132
133
134 Signal-peptide Lorsqu une protéine est synthétisée, sa structure secondaire ou tertiaire se construit au fur et à mesure de la traduction et elle est libérée dans le cytoplasme. Lorsque cette protéine est destinée à être incorporée dans les membranes ou les organites de la cellule, la partie codante de l ARN messager commence par une séquence de quelques acides aminés qui sert d adresse pour l incorporation de cette protéine dans la membrane. Ces peptides orientent la destinée de la protéine : incorporation dans les membranes (reticulum endoplasmique, appareil de Golgi, membrane plasmique, lysosomes...), entrée dans les mitochondries, excrétion hors de la cellule via l appareil de Golgi, etc... Le signal-peptide est un peptide d adressage situé à l extrémité NH 2 - terminale des protéines à excréter. Parce que sa fonction est de pénétrer dans la membrane du reticulum endoplasmique, sa structure est riche en acides aminés hydrophobes (Phe, Leu, Ile, Met, Val).
135
136 Polyribosomes liés Les polyribosomes qui se forment sur un messager comportant un signal peptide ont une évolution légèrement différente. La traduction s arrête peu après la synthèse du signal-peptide, dès que celui-ci apparaît hors de la structure du ribosome et se lie avec la SRP (SRP = Signal Recognition Particle), qui inhibe l élongation. Pour que l élongation puisse se poursuivre, il faut que la SRP soit reconnue spécifiquement par un récepteur de la membrane du reticulum endoplasmique. Dès que cette liaison est établie, le ribosome est lié par la ribophorine, toujours dans la membrane du reticulum, près d une protéine qui ouvre un pore à travers cette membrane. Le SRP écarté, l élongation va reprendre.
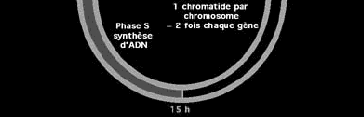
137 Le peptide en cours de synthèse est alors dirigé à travers cette membrane pour se développer dans la lumière du reticulum endoplasmique. A la face interne de la membrane une endopeptidase spécifique va couper le signal peptide et la synthèse se poursuivra jusqu à la terminaison. La protéine sera enfin incorporée dans une membrane ou exportée, à travers l appareil de Golgi, vers l extérieur de la cellule. L adressage des protéines vers les mitochondries fait appel à un autre type de peptide d adressage qui conduit les peptides à traverser la membrane mitochondriale.
138
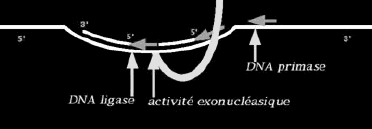
139 modifications chimiques se produisent après l incorporation des acides aminés dans la structure primaire de la protéine (traduction); on les appelle: modifications post-traductionnelles. On distingue des modifications cotraductionnelles qui se produisent alors que la traduction se poursuit encore et que le peptide naissant est encore attaché au ribosome qui l a construit, des modifications post-traductionnelles proprement dites qui ont lieu dans la cellule, dans les organites ou hors de la cellule. On appelle protéine mature la forme chimique définitive que la protéine montrera au moment où elle remplira sa fonction dans l organisme.
140
141 Addition de sucres: Glycosylation
142
143 La réplication Au cours de la vie de la cellule (cycle cellulaire, d une division mitotique à la suivante), l ADN doit être dédoublé pour que chaque cellule fille reçoive un génome complet dans son noyau. Cette synthèse se produit à la phase S (au milieu du cycle cellulaire) grâce à l activité de la DNA-polymérase a. D autres DNA-polymérases participent à la réparation de l ADN lésé ou à la réplication de l ADN mitochondrial.

144
145 Le cycle cellulaire La vie de la cellule se déroule entre deux mitoses. Chez les Mammifères, cette période dure en moyenne 30 heures bien qu il y ait des cellules dont la vie soit très courte ou au contraire très longue. Durant ces trente heures la cellule traverse quatre phases : la phase G1 où le génome étant diploïde, chaque gène est représenté en deux exemplaires. La chromatine est accessible aux RNA-polymérases qui transcrivent les gènes en messagers, qui seront à leur tour traduits. Phase de préparation à la synthèse d ADN. vers la moitié du cycle commence la réplication de l ADN : les DNApolymérases vont mettre environ 8 heures (phase S) pour recopier en double l ADN de chaque chromosome. Durant cette phase la transcription est inhibée. Phase de synthèse d ADN. puis la cellule entre en phase G2 où chaque gène est représenté en quatre exemplaires. La chromatine est à nouveau accessible aux RNA-polymérases qui recommencent à transcrire. Phase de réparation des mutations. enfin survient la mitose, qui donne naissance à deux cellules filles. Chacune recevra une des copies identiques de l ADN de chaque chromosome et chaque gène y sera représenté en deux exemplaires.
146
147 DNA polymérase Les DNA-polymérases sont des enzymes du noyau cellulaire qui agissent en phase S du cycle pour doubler systématiquement l ensemble du génome diploïde. Elles utilisent des désoxyribonucléotides triphosphates (datp, dctp, dgtp et dttp) et des amorces de RNA synthétisées par une DNA primase (RNA polymérase capable de synthétiser un court brin de RNA complémentaire d un brin de DNA). Elles synthétisent l ADN nouveau par fragments qui sont liés entre eux par une DNA-ligase. Elles ont aussi une activité exonucléasique, qui leur permet en particulier d hydrolyser l ARN des amorces.
148 Réplication Les DNA-polymérases commencent leur synthèse en de nombreux points d initiation de la réplication. La synthèse de DNA commence sur des amorces de RNA constituées par la DNA-primase. La réplication se poursuit dans une direction : dans ce sens l un des deux brins de l ADN (brin «avancé») est parcouru par l enzyme dans le sens ce qui permet la synthèse d un autre brin dans le sens Les DNA-ligases assurent ensuite la liaison entre les différents fragments de l ADN nouveau. La synthèse de l autre brin (brin «retardé») est plus complexe parce que l enzyme parcourt ce brin de La DNA primase synthétise des amorces de quelques nucléotides en avant de la zone de réplication, et la DNA polymérase construit à la suite de petits fragments d ADN dans le sens La DNA polymérase hydrolyse en avançant l amorce de RNA du fragment précédent (activité exonucléasique). Les petits fragments seront ensuite reliés entre eux par la DNA-ligase.
149
150 LA RÉPLICATION DE L ADN CHEZ LES PROCARYOTES Caractéristiques générales. réplication dite semi-conservatrice. Ceci signifie que sur les deux brins d ADN, on a toujours un brin d ADN qui provient d un des deux brins de l ADN parental et un brin nouvellement formé. A chaque réplication, les deux brins d ADN parental se séparent, chacun de ces deux brins sert de matrice pour la synthèse d un brin complémentaire. Les éléments nécessaires à la réplication de l ADN. La réplication de l ADN nécessite: - Tout d abord, une matrice d ADN constituée par un brin parental. - La présence de nucléotides propres à l ADN, c est-à-dire contenant du 2 - désoxyribose, des bases A, T, G et C et sous forme de nucléosides triphosphates: datp, dttp, dctp et dgtp (on écrit souvent pour les dénommer dntp). - La présence de nombreux enzymes ( P dna A, Pdna B= hélicase, P SSB =stabilisateur de sb, Primases, polymérases, exonucléases, ligases & P Tus=terminaison ). - La présence de certains ions (cations bivalents: Mg 2+, ce cation est indispensable pour la réplication de l ADN).
151 Les mécanismes de l initiation de la réplication L unité d ADN où se produit la réplication est appelée le réplicon. Il a une origine où est initiée la réplication et une terminaison où est arrêtée la réplication. L ADN bactérien constitue à lui seul un réplicon. A partir d un point d initiation, la réplication peut progresser soit de manière unidirectionnelle, soit de manière bidirectionnelle. Chez les procaryotes, à partir d une origine de la réplication (ou oeil de réplication), la réplication progresse dans les deux sens: bidirectionnel Chez E. coli, l ADN bicaténaire a une origine unique de la réplication appelée OriC. Le locus OriC est une séquence de 250 pb: tandem de trois séquences presque identiques de 13 nucléotides (séquences de type GATCTNTTNTTTT).
152 Au locus OriC, quatre sites de liaison comportant un motif de paires de bases lient Des copies multiples d une protéine appelée dnaa ou protéine d initiation de la réplication, codée par le gène dnaa. Ces liaisons nécessitent de l ATPet sont indispensable à la transformation localisée de l ADN double brin en ADN simple brin. Ces complexes vont ensuite fixer 2 ADN hélicases, ou protéine dnab, une hélicase pour chacun des brins d ADN codée par le gène dnab. L hélicase déroule l ADN (en présence d ATP et d une autre protéine appelée dnac) et définit la fourche de réplication.
153 Les brins séparés de l ADN sont stabilisés sous forme simple brin grâce à la fixation de protéines appelées SSB (pour «single strand binding»). Ces protéines SSB empêchent les deux brins d ADN de se réapparier. le complexe primosome entre l hélicase et une enzyme primase (ou protéine dnag) permet la Synthèse des amorce d ARN (9-12 nucléotides). L ADN polymérase III s insère au niveau de la fourche de réplication et utilise l amorce d ARN pour la synthèse de l ADN. L ADN polymérase III possède deux sites actifs capables de synthétiser les nouveaux brins d ADN au niveau de la fourche de réplication.
154 La réplication n est pas identique entre les deux brins d ADN. Pour l un des brins: la réplication s effectue de manière continue, brin avancé, sens 5 à 3 Pour l autre brin elle est discontinue, brin retardé, orienté de manière anti-parallèle 3 à 5 synthèse discontinue des fragments d OKAZAKI. le brin matrice du brin retardé forme une boucle autour d un des deux sites actifs de l ADN polymérase III. intervention de la primase et de l ADN polymérase III pour la synthèse du brin retardé d ADN dans le sens 5 à3 Après synthèse la boucle est défaite et une nouvelle est reformée au niveau de l ouverture de la fourche de réplication.
155 L ADN polymérase III utilise l ADN comme matrice (sens 5 à3 ), l ARN comme amorce & comporte 7 sous-unités catalytiques. les amorce d ARN sont synthétisées par une ARN polymérase ou primase. l ADN polymérase III allonge cette amorce d ARN L hydrolyse et le remplacement des amorces d ARN, intervention de la RNAse H, l ADN polymérase I et une DNA-ligase. Les ADN polymérases I et III impliquées dans la réplication de l ADN présentent plusieurs fonctions enzymatiques: une fonction polymérasique: ajoute un nucléotide à l extrémité 3 OH d un acide nucléique, une fonction exonucléasique 5 à3 (activité de réparation de l ADN) & une fonction exonucléasique 3 à5 (l enzyme vérifie le dernier nucléotide mis en place). Terminaison de la réplication. IL existe au moins 6 sites de terminaison ou sites (ter) de terminaison dans le génome bactérien. Une protéine spécifique appelée Tus peut se lier aux sites de terminaison. Cette liaison arrête la protéine dnab (hélicase).
156 LA RÉPLICATION DE L ADN CHEZ LES EUCARYOTES Caractéristiques générales. La réplication de l ADN chez les eucaryotes est tout à fait comparable à la réplication de l ADN chez les procaryotes. Elle est généralement bidirectionnelle, elle est discontinue entre les deux brins d ADN. Des amorces d ARN sont nécessaires. Cette réplication est également complémentaire, antiparallèle et dans le sens 5 à 3. Caractéristiques particulières. La réplication se fait en de nombreux points d initiation. Elle fait intervenir un nombre d ADN polymérases plus important que chez les procaryotes. De nombreuses protéines interviennent comme facteurs de réplication. Les changements subis par les nucléosomes au cours de la réplication de l ADN eucaryote & la réplication de l ADN des extrémités chromosomiques (ou télomères) ont un rôle primordial dans la régulation du cycle & de la division cellulaire.
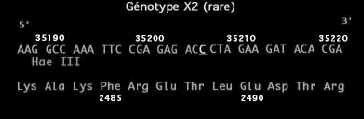
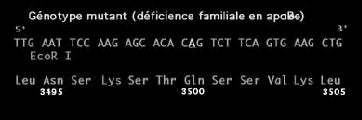
157
158
159
160
161 Mutations génétiques & systèmes de réparations LES TYPES DE MUTATION DE L ADN. Les mutations sont définies comme un changement transmissible dans le matériel génétique. Ces mutations peuvent concerner les cellules germinales ou les cellules somatiques. Elles peuvent être spontanées ou consécutives à des altérations des bases puriques ou pyrimidiques. Elles peuvent être provoquées par des dysfonctionnements au cours de la réplication ou de la réparation de l ADN. On peut classer les mutations selon l étendue de la lésion de l ADN: les macrolésions et les microlésions.
162 Les macrolésions de l ADN. Dans ce groupe, on peut ranger: - Les délétions (amputations de matériel génétique d amplitude variable). - Les duplications. - Les amplifications (multiplication de séquences. - Les fusions de gènes. - Les inversions (changement d orientation tête-bêche d un segment variable d ADN). - Les insertions de séquences d ADN.
163 Les microlésions. Dans ce cadre, on range les mutations ponctuelles de l ADN. délétion, insertion ou substitution, d un nucléotide par un autre. La substitution d une paire de bases par une autre peut s agir d une transition ou d une transversion; La transition: correspond au remplacement d une base purique par une autre base purique ou une base pyrimidique par une autre base pyrimidique. La transversion: correspond au remplacement d une base purique est par une pyrimidine ou d une pyrimidine par une purine.

164
165 substitution non-synonyme: incorporation d un acide aminé différent dans la protéine substitution synonyme incorporation du même acide aminé dans la protéine Les substitutions synonymes résultent de la dégénérescence du code génétique. Lorsqu une substitution non-synonyme se produit dans le DNA (génotype) aboutit à l incorporation d un acide aminé fonctionnellement différent dans la structure primaire d une protéine, il en résulte l apparition d un caractère différent (mutation) dans le phénotype de l individu. Toute autre modification entraînant une protéine traduite plus longue ou plus courte, ou encore un décalage de la lecture des codons, se traduit par une séquence primaire différente et donc la plupart du temps une mutation dans le phénotype de l individu.
166
167

168
169 Délétion délétion, c est à dire suppression d un ou de plusieurs nucléotides Les délétions sont d importance variable selon le nombre de nucléotides suprimés: de 1 ou 2 nucléotides, elles décalent le cadre de lecture (codons) de 3 nucléotides, elles aboutissent à la suppression d un acide aminé dans la protéine ex-primée de grande longueur, elles peuvent supprimer l expression d un ou de plusieurs exons, voire d un gène entier.

170
171 Insertion: Insertion, c est à dire addition d un ou de plusieurs nucléotides. Les insertions sont d importance variable selon leur longueur : de 1 ou 2 nucléotides, elles décalent le cadre de lecture (codons) de 3 nucléotides, elles aboutissent à l addition d un acide aminé dans la protéine exprimée de grande longueur, elles peuvent modifier complètement la traduction des exons dans les introns ou dans les exons nontraduits, on rencontre souvent de longues insertions de structure variable qui ont des effet??? sur l expression du gène.

172
173 LES SYSTÈMES DE SAUVEGARDE ET RÉPARATEURS DE L ADN GÉNÉRALITÉS. L ADN est sensible à des agressions soit chimiques soit physiques. Ces agressions peuvent entraîner la perte d une base (purique ou pyrimidique), mais aussi entraîner la rupture d un brin ou des deux brins d ADN. La gravité des lésions que l ADN subit en permanence est liée au fait que cette molécule est unique au sein d une cellule et au caractère transmissible de ces atteintes. Des mécanismes puissants, cependant non infaillibles sont présents dans la cellule pour déceler et corriger les lésions de l ADN.
174 ORIGINE DES ALTÉRATIONS DE L ADN. Les agents physiques. L ADN peut être agressé par des agents physiques comme les rayons X, les rayons gamma, les rayons ultra-violets (UV). Ces agents physiques sont des agents mutagènes (mutagènes= qui provoquent des mutations dans l ADN) les plus anciennement connus. Les rayons UV provoquent la formation de dimères de thymine mais aussi de dimères de thymine-cytosine. La présence de tels dimères entraîne un blocage de la réplication de l ADN et de la transcription. Sans réparation, leur présence serait donc léthale pour la cellule.
175 Les agents chimiques. L ADN peut être agressé par un nombre considérable de dérivés chimiques. Nous citerons plus spécialement: - Les agents alkylants. Ils entraînent l addition de groupes alkyles aux bases (souvent des groupes -CH 3 ). Les agents alkylants sont utilisés comme médicaments anticancéreux. - L acide nitreux (produit dérivé des nitrates ou nitrites présents dans les produits alimentaires ou dans les engrais). Cet agent chimique transforme par exemple, par désamination oxydative, l adénine en hypoxanthine. - Certains composés chimiques ressemblent à des bases puriques ou pyrimidiques. Ils peuvent être incorporés dans l ADN à la place des bases normales (cas du bromouracile qui ressemble à la thymine). - Enfin, certains agents s intercalent entre les deux brins d ADN, ce sont des agents intercalants. Ils perturbent ainsi la réplication. C est le cas du révélateur de l ADN utilisé dans de nombreux laboratoires de biologie moléculaire: le bromure d éthydium ou B.E.T.
176 LES TYPES DE DOMMAGES DE L ADN. Perte d une base. La liaison glycosylique liant dans l ADN les bases avec le désoxyribose est relativement labile dans des conditions physiologiques. A l intérieur d une cellule de mammifère, plusieurs milliers de bases puriques et plusieurs centaines de bases pyrimidiques sont spontanément perdues dans le génome d une cellule haploïde et par jour. La perte d une base purique ou pyrimidique crée un site appelé apurinique ou apyrimidique (ou site AP). Modification d une base. La déamination. Les groupes -NH 2 des bases peuvent être instables. Ils sont convertis en groupes cétoniques =C=O. Ainsi, la cytosine désaminée est transformée en uracile, l adénine en hypoxanthine etc...
177 Les modifications chimiques. Beaucoup d agents chimiques peuvent réagir avec les bases des acides nucléiques. Il peut s agir des espèces oxygénées réactives (peroxyde d hydrogène, radicaux hydroxyles et peroxyles) mais aussi des radiations ionisantes (rayons X, rayons gamma). Ces espèces peuvent conduire à des hydroxylations sur les noyaux des bases (ajout de groupes -OH). L alkylation peut être facilement réalisée sur les bases de l ADN par addition de groupes méthyles ou alkyles par de très nombreux agents alkylants ou même par l action de simples donneurs de groupes méthyles présents dans la cellule (sous forme de S- adénosylméthionine, par exemple). L action des rayons ultra-violets. La lumière UV est absorbée par les bases des acides nucléiques et des modifications chimiques peuvent en découler. Les produits les plus courants fournis par ces réactions photochimiques sont surtout les dimères thymine-thymine (T-T), suivis des dimères thymine-cytosine (T-C) et les dimères cytosine-cytosine. Il s agit de structures stables entraînant une distorsion considérable de la structure de l ADN.
178 Les erreurs de la réplication. Nous avons vu qu au cours de la réplication de l ADN, les ADN polymérases présentent une activité exonucléasique 3 à 5 qui leur permet de vérifier si la dernière base introduite correspond bien aux conditions classiques d appariement. Cette activité d édition diminue considérablement les erreurs possibles. Au cours de la réplication, il a été estimé que l ADN polymérase III introduit un nouveau nucléotide avec un taux d erreur de 1/100 nucléotide incorporé, mais avec la fonction d édition, finalement le taux d erreur passe à 1/1000 nucléotide incorporé. Cette différence considérable correspond à la fidélité de la réplication. Cependant, la probabilité de produire des mauvais appariements ou des petites insertions ou délétions pendant la réplication est loin d être négligeable. Dès lors des mécanismes efficaces de réparation doivent être mis en oeuvre.
179 Formation de ponts entre les brins de l ADN ou entre des protéines et l ADN. Certains agents chimiques (psoralènes par exemple) ou physiques (UV, radiations ionisantes) peuvent conduire à des interactions stables («crosslinks») entre les deux brins d ADN. Des interactions stables peuvent être également observées entre des protéines et les brins d ADN. Les coupures des brins d ADN. Les radiations ionisantes peuvent conduire à des coupures d un brin d ADN ou à des cassures des deux brins d ADN.
180 LES MECANISMES RÉPARATEURS DE L ADN. La possibilité de réversibilité des dommages sur l ADN. C est la possibilité de réparer l ADN la plus simple, mais dans bien des cas elle est impossible à réaliser pour des raisons cinétiques et / ou thermodynamiques. Dans certains cas, des enzymes spécifiques comme les photolyases peuvent transformer réversiblement un ADN lésé avec présence de dimères pyrimidiques en un ADN normal en absorbant de la lumière. Les photolyases sont présentes dans les bactéries, les plantes, les champignons mais elles sont absentes chez les mammifères.
181 Correction des erreurs d appariements. Beaucoup d erreurs d appariements (ou «mismatches») sont dues à des erreurs au cours de la réplication de l ADN. Cependant, des erreurs d appariements peuvent être également produites à la suite d une désamination de la 5 -méthyl cytosine pour produire de la thymine qui ne s apparie pas correctement avec G. Chez E. coli, la présence d un mauvais appariement localisé est décelée par une protéine spécifique appelée protéine MutS. La protéine MutS reconnaît des erreurs d appariements mais aussi des insertions ou des délétions de quelques nucléotides. Une protéine supplémentaire, la protéine MutL stabilise le complexe formé entre MutS et la portion d ADN double brin qui présente le mauvais appariement. Ce complexe va activer une protéine MutH qui va couper le brin nouvellement synthétisé. Il est remarquable que ce système de coupure différencie le brin parental et le brin nouvellement synthétisé par methylation.
182 En effet, dans l ADN d E. coli, les séquences GATC sont normalement méthylées. Dans le brin nouvellement synthétisé, la méthylation n est pas immédiatement réalisée. La protéine MutH va couper le brin nouvellement synthétisé à l opposé de la séquence GATC méthylée du brin parental. Une coopération avec la protéine uvrd (hélicase II) est nécessaire. Le brin nouvellement formé est alors partiellement dégradé et resynthétisé correctement par l ADN polymérase III. La soudure finale est assurée par l ADN ligase. Les organismes eucaryotes possèdent de nombreux systèmes de réparation des erreurs d appariements. Certains malades atteints d une forme héréditaire de cancer du colon (syndrome de Lynch) présentent des mutations dans les gènes des enzymes de réparation des erreurs d appariements.
183 L excision-réparation de base. Ce type de réparation est communément utilisé pour éliminer les bases incorrectes (comme l uracile) ou les bases transformées par des agents alkylants. Il comprend trois étapes successives: Tout d abord, une élimination de la base incorrecte par une ADN N-glycosylase avec apparition d un site AP. Puis, une coupure du brin d ADN endommagé en 5 du site AP, créant ainsi un -OH (3 ) adjacent au site AP. Enfin, l extension à partir de cet -OH (3 ) par une ADN polymérase est réalisée avec excision du site AP. La soudure finale est assurée par une ADN-ligase.
184 chez les mammifères et l Homme, les doublets nucléotidiques CG peuvent être méthylés au niveau de la cytosine et que cette méthylation était en relation directe avec l inactivation des gènes correspondants. La désamination des cytosines méthylées aboutit à la formation de la thymine. Une ADN glycosidase particulière reconnaît spécifiquement le mésappriement TG et élimine T. Cependant, l efficacité de ce système est loin d être parfaite. Souvent, les mutations ponctuelles de l ADN concernent ces cytosines méthylées, on parle de points chauds de mutation.
185
186
187
188 L excision-réparation de nucléotide. Bien que l excision-réparation de base joue un rôle très important comme processus de réparation de l ADN, elle ne peut pas suffire à corriger toutes les erreurs dans l ADN. En particulier, la disponibilité des ADN N-glycosylases ne peut pas être étendue à toutes les altérations possibles des bases de l ADN. Un autre mécanisme plus flexible de réparation est impliqué. Ce mécanisme est appelé excision-réparation de nucléotide. Il est disponible dans tous les organismes vivants et il implique les étapes suivantes: Tout d abord, la reconnaissance du dommage sur l ADN. Puis une liaison d un complexe multi-protéique avec le site endommagé. Une double excision du brin endommagé suit avec coupure plusieurs nucléotides en 5 et en 3 de celui-ci. La lacune présente est comblée par une ADN polymérase. Une soudure finale est ensuite assurée par une ADN-ligase.
189 Chez E. coli, trois protéines, les produits des gènes uvra, uvrb et uvrc sont responsables de la reconnaissance du dommage et de la coupure de l ADN Le modèle actuel implique la formation initiale d un complexe entre deux protéines uvra et uvrb et ceci avec hydrolyse d ATP Ce complexe se lie avec l ADN préférentiellement au niveau de la zone endommagée (dimères de thymine) Une autre protéine uvrc se lie avec uvrb ceci entraîne une coupure en 5 (7 nucléotides avant le dommage) et en 3 (4 nucléotides après le dommage) Intervient, la proteine du gène uvrd, qui est une hélicase II, elle déplace l oligonucléotide endommagé l ADN polymérase I ou l ADN polymérase II comble la lacune résultante La soudure finale est assurée par une ADN-ligase.
190
191 Un tel système de réparation existe également chez les cellules eucaryotes. On connait d ailleurs chez l Homme, des anomalies génétiques du système de réparation par excision-réparation responsables de maladies génétiques graves, la xérodermie pigmentaire (entraînant une sensibilité accrue aux rayons solaires et une augmentation du risque de cancers cutanés), mais aussi le syndrome de Bloom ou la maladie de Fanconi.
192 Les recherches thérapeutiques Le petit nombre de malades et le faible débouché commercial qu'offrent les maladies rares telles que le XP Mais depuis quelques années, les recherches se sont multipliées et elles ont permis de mieux comprendre les mécanismes de la maladie. Evolution des recherches: En 1988: Caractéristiques cliniques génétiques et cellulaires. Développement d'un test anténatal. Activation des oncogènes dans les tumeurs épithéliales isolées de malades atteints du XP En 1990: Une carence en ADN hélicase serait responsable de certaines formes de XP et du syndrome de Cockayne. Clonage du gène et de l'adnc de groupe A du XP En 1996: La thérapie génique envisageable. Une nouvelle technique de correction génétique des cellules a été mise au point. Elle permet, grâce à une construction rétrovirale, de produire des lignées de fibroblastes de peau de malades au phénotype guéri. Cela constitue donc une première étape vers la correction des kératinocytes de malades et peut-être un jour la production in vitro d'un épithélium génétiquement corrigé pouvant servir de greffon. En 1998: Comment l'absence d'interaction entre une hélicase et son régulateur est à l'origine d'une maladie génétique. En 2001: Reconstruction in vitro de peau de patients atteints de XP
193
194 Les réparations post-réplicatives. Si le système de réparation par excision-resynthèse de l ADN est débordé, et surtout si les zones d ADN à réparer coïncident avec la présence de fourches de réplication, il est évident que le brin porteur de la lésion devient impropre à servir de matrice. Dans de telles conditions, le brin parental contiendra un dimère de thymine et l autre brin fils contiendra une lacune, on parle de lacune post-réplicative. C est une lésion grave de l ADN. Pour traiter de telles lésions, un système de réparation approprié va être mis en oeuvre. Il s agit du système des protéines Rec (pour Recombinaison). Ce système de réparation fait donc intervenir une recombinaison. La recombinaison correspond à un échange d ADN ente deux segments homologues. Cette correction fera également intervenir le système de réparation par excision-resynthèse de l ADN. Chez E. coli, la production de la protéine de recombinaison appelée RecA est réduite dans les conditions normales. Ceci est lié à une répression de la synthèse par un répresseur protéique appelé LexA (voir contrôle de la transcription).
195
196
197 Le système S.O.S. Le système S.O.S. a été décrit initialement chez les bactéries. Il concerne au moins une vingtaine de gènes dont les produits protéiques sont impliqués dans les mécanismes de réparation et de recombinaison. Ce système est remarquable parce qu il est inductible, c està-dire mis en oeuvre par l agression physique ou chimique. La protéine de recombinaison RecA joue un rôle central dans un tel mécanisme de sauvegarde. La synthèse de cette protéine est normalement réprimée par une protéine appelée LexA ou répresseur. Si un besoin important de protéine RecA apparaît, la répression est levée grâce à une propriété particulière de la protéine RecA qui est capable de cliver son répresseur.

198
199 Biologie moléculaire du cancer Mécanisme de l oncogénèse INTRODUCTION La survenue dans un organisme d'une tumeur cancéreuse est liée à l'émergence d'un clone cellulaire échappant aux lois qui régissent la prolifération et la cohabitation cellulaire normales. La cellule cancéreuse se caractérise par 2 propriétés fondamentales : 1/ la capacité de se reproduire au delà des limites fixées par le renouvellement naturel du tissu auquel elle appartient et 2/ le pouvoir de coloniser des territoires tissulaires normalement réservés à d'autres catégories cellulaires. Ces 2 propriétés sont communes à l'ensemble des cellules d'une même tumeur ce qui suggère que toute la population est issue d'une seule cellule devenue anormale.
200 MECANISME DE L'ONCOGENESE L'ADN = siège des altérations successives L origine monoclonale de la population de cellules formant une tumeur a pu être vérifiée dans de nombreux cas à l'aide de marqueurs génétiques L'analyse d'une tumeur maligne indique que toutes les cellules expriment la même forme et donc que la masse tumorale correspond à l'expansion d'un clone cellulaire.
201 L'anomalie responsable de la transformation cancéreuse, virale ou non, est transmissible par division cellulaire et consiste en une altération majeure de l'information génétique, c'est à dire une mutation de l'adn. Du reste, la majorité des agents carcinogènes s'avèrent être également des agents mutagènes.
202 Mise en évidence des oncogènes Les "oncogènes" apparaissent comme des homologues de gènes cellulaires physiologiquement impliqués dans le contrôle des processus de maturation, de division et de différenciation cellulaire et dont l'altération ("activation") peut entrainer la transformation d'une cellule normale en cellule maligne. La compréhension des mécanismes d'activation des oncogènes passe par la description des modèles expérimentaux qui ont permis de les étudier: L'intégration virale dans le génome, La transfection d'adn tumoral, Anomalies cytogénétiques caractéristiques de certaines tumeurs, ADN tumoral muté dans les cas de cancers héréditaires, ADN constitutionnel dans des formes familiales de cancers.
203 Filière virale Le chef de file des retrovirus tumorigènes est le Virus du Sarcome de Rous (RSV), virus à ARN, dont l'intégration dans le génome d' une cellule est possible grâce à la retrotranscriptase, (une enzyme qui permet la transcription de l'arn en ADN). Son pouvoir transformant est entièrement contenu dans le gène src, dont l'insertion dans le génome de la cellule hôte peut provoquer la transformation cancéreuse. On en retrouve un équivalent dans le génome de toutes les cellules normales de l'hôte et de nombreuses autres espèces. Cet homologue cellulaire d'un oncogène viral est un gène cellulaire non transformant ou protooncogéne.
204 De nombreux retrovirus tumorigènes possèdant des gènes transformants v-onc homologues de gènes cellulaires normaux c-onc ont été identifiés. L'étude systématique des retrovirus capables de transformer les cellules in vitro a permis d'identifier de nombreux autres oncogènes viraux correspondant chacun à la version un peu modifiée d'un ou plusieurs oncogène(s) cellulaire(s). Leur origine animale ainsi que l'emplacement chromosomique de leur homologue humain sont connus.
205 Oncogenes Retroviraux Les proto oncogènes codent pour des protéines qui jouent un rôle central dans la stimulation de la division cellulaire. Les oncogènes d origine retro virale sont classés en 4 groupes: 1. Facteurs de croissance, (C-sis) & récepteurs de facteurs de croissance (C fms & CerbB); 2. GTP-binding proteins, (C h-ras, C k-ras, C n-ras); 3. Tyrosine specific protein kinases (C abl, C fes, C fgr, C fps, Cros, C src, and C yes); serine/threonine specific protein kinases (C mil, C mos, C raf). 4. Transcription regulators (C fos, C jun, C erb A, C myc, C myb, &Cets). Les protéines produites par les Proto oncogènes, sont alterées par des mutations, infections viral, ou agents carcinogénique. Ces protéines oncogéniques codent pour des protéines modifiées et sur exprimées qui stimulent une division cellulaire anormale.
206 La haute conservation de ces protooncogènes dans toutes les espèces animales étudiées, y compris l'homme, suggère leur intervention dans des fonctions primordiales communes à tous les eucaryotes. lls diffèrent le plus souvent de leur homologue cellulaire par des altérations minimes entraînent des modifications de leur structure ou de leur fonction qui semblent suffisants à leur conférer le pouvoir transformant. L'activation peut être de type qualitatif Ou de type quantitatif.
207 Pouvoir transformant La technique de transfection de fragments d'adn extraits de lignées cellulaires malignes ou de cellules tumorales fraiches à des cellules fibroblastiques en culture (cellules NIH 3T3) permet d'identifier des gènes porteurs d'un pouvoir transformant puisque capables de conférer à ces cellules un phénotype malin. La première mise en évidence de gènes transformants non viraux a été réalisée par transfection de petits fragments d'adn permettant la transformation maligne. Les oncogènes ainsi isolés sont nombreux. Le chef de file en est la famille des gène ras, isolés à partir d'une tumeur spontanée de la vessie chez l'homme et dont l'activation se fait toujours par une mutation ponctuelle sur un seul codon. Cette altération conduit à la production d'une protéine mutée, ne différant de la protéine native que par un acide aminé, modèle de mutation purement qualitative.
208 Filiére cytogénétique L'amélioration des techniques de cytogénétique a permis de reconnaître, parmi les très nombreuses anomalies chromosomiques retrouvées dans différents types de cancers, la spécificité de certains remaniements non aléatoires. Les trois types de lésions morphologiques connues sont: les amplifications géniques, les translocations chromosomiques et les délétions. Amplifications de proto-oncogènes L'amplification correspond à l'accumulation d'un grand nombre de copies d'un même gène. Son existence est soupçonnée en analyse cytogénétique sur la présence de régions chromosomiques se colorant de manière uniforme appelées Homogeneous Staining Régions (HSR) ou se présentant comme des mini chromosomes surnuméraires (chromosomes "double minute"). Les techniques de biologie moléculaire ont permis de montrer que ces anomalies correspondent à une amplification génique, portant sur certains proto-oncogènes.
209 Translocations chromosomiques Certaines translocations provoquent des juxtapositions aberrantes de gènes ou des fusions de 2 gènes. juxtapositions aberrantes une translocation chromosomique interessant le chromosome 8 et le chromosome 14. Il a été démontré que la translocation entraine un échange de matériel entre la région du proto-oncogène c- myc située sur le chromosome 8 et une région codant pour des gènes d'immunoglobulines (Ig). L'activation de c-myc se traduit par son expression anormale en quantité ou en durée dans le cycle cellulaire. La translocation survient à l'occasion d'une erreur de recombinaison génétique au cours des réarrangements des gènes des Ig.
210 fusions de 2 gènes Elle se caractérise par la présence dans 90 % des cellules tumorales d'une translocation entre les chromosomes 9 et 22 au cours de laquelle le protooncogène c-abl du chromosome 9 est fusionné avec une séquence du chromosome 22 toujours retrouvée au niveau du point de cassure et appelée pour cette raison "breakpoint cluster region" (bcr); le produit qui résulte de cette fusion est une protéine anormale qui présente l'intérêt diagnostique de pouvoir être détectée en l'absence d'anomalie.
211 Délétions chromosomiques Elles ont été mises en évidence dans le cadre de l'étude des cancers héréditaires où elles ont conduit à la notion d anti-oncogènes". La constatation de la perte de la tumorigénicité d'une cellule hybride obtenue par fusion entre une cellule maligne et une cellule normale, a conduit à la notion de "gènes suppresseurs" dont l'inactivation pourrait tout aussi bien être responsable de la croissance tumorale que l'activation des oncogènes. Le modèle tumoral choisi pour vérifier leur existence a été le rétinoblastome, une tumeur maligne dans laquelle on distingue 2 formes cliniquement bien distinctes : la forme familiale, héréditaire, où s'observent fréquemment des atteintes occulaires bilatérales et où les cellules tumorales présentent souvent une délétion sur le bras long du chromosome 13, et la forme sporadique, plus fréquente, où les atteintes sont le plus souvent unilatérales.
212 L'hypothèse émise reposait sur les données cliniques et était basée sur la nécessité d'une double mutation sur le même chromosome pour que la prolifération tumorale soit possible. La première mutation sur le gène en cause, constitutionnelle et transmissible dans la forme familiale, est acquise dans la forme sporadique. Le second évènement mutationnel, acquis dans tous les cas au niveau de la cellule somatique, entraine l'homozygotie pour le "gène du rétinoblastome". Plus récemment, cette hypothèse a été confirmée au niveau moléculaire et l'apparition d'un rétinoblastome s'avère liée à l'inactivation des 2 allèles d'un gène récessif normal responsable du contrôle de la croissance normale des cellules rétiniennes. Ce gène, dit Rb, localisé sur le chromosome 13 a été identifié et cloné. Dans les formes héréditaires de la maladie, la perte constitutionnelle du premier allèle est due à une mutation ou à une délétion chromosomique et dans tous les cas la deuxième mutation est représentée par la perte de l'allèle normal restant.
213 Le gène Rb est le chef de file de nombreux autres "anti-oncogènes" ou "gènes suppresseurs", identifiés dans d'autres tumeurs malignes et codant pour des protéines qui sont des éléments clés du "freinage" exercé en permanence par la cellule pour contrôler sa division, sa maturation et sa différenciation normales. Conséquences de l'activation des oncogènes : les protéines oncogéniques Le résultat de l'activation d'un oncogène est la production d'une protéine dite "oncogénique" qui ressemble à la protéine normalement codée par le gène cellulaire correspondant mais qui est anormale. Il peut s'agir d'une altération : - soit qualitative, liée à une mutation portant sur les séquences codantes du gène et entraînant l'apparition d'une protéine mutée, - soit quantitative, liée à une mutation portant sur les séquences régulatrices non codantes du gène et entraînant la production de quantités anormales d'une protéine. Le résultat de l inactivation (délétion) d'un anti-oncogène gène su presseur de tumeur est la perte d une protéine qui déclenche l apoptose des cellules mutantes qui peuvent donc échapper aux systèmes de réparation et de control de la division cellulaire.
214 Les protéines codées par les oncogènes peuvent ainsi être regroupées en quatre grandes catégories fonctionnelles : Les protéines analogues de facteurs de croissance cellulaires. La protéine codée par l'oncogène sis par exemple, représente un analogue muté du PDGF (Platelet Derived Growth Factor) et son pouvoir transformant dépend de la présence du récepteur correspondant à la surface des cellules. Les protéines analogues de récepteurs de facteurs de croissance. La protéine codée par l'oncogène erb B par exemple, représente un analogue muté du récepteur cellulaire pour l' EGF (Epidermal Growth Factor). La fixation du facteur de croissance correspondant au récepteur entraîne la transmission d'un signal de division cellulaire par l'intermédiaire d'une activité protéine kinase. La protéine oncogénique pourrait possèder la même capacité fonctionnelle mais répondre différemment aux systèmes de régulation, ce qui aurait pour effet le détournement des processus de division cellulaire au profit de la prolifération tumorale.
215 Les protéines impliquées dans une activité enzymatique. Les mieux connues dans cette catégorie sont les protéines kinases membranaires dont le chef de file est la protéine oncogénique pp60 src, codée par le gène src du RSV. Elles exercent un rôle régulateur important dans le fonctionnement de nombreuses protéines cellulaires, comme les protéines du cytosquelette ou les protéines impliquées dans le métabolisme des prostaglandines. Les protéines à activité nucléaire. Les protéines transformantes codées par les gènes c-myc sont présentes dans le noyau. Elles agissent vraisemblablement au début du cycle cellulaire, peu avant la duplication, par liaison directe à l'adn à un stade très terminal de la cascade.
216 Les protéines des antioncogènes La protéine oncogénique p 105-Rb, codée par le gène "suppresseur" Rb appartient au système de contrôle de la prolifération cellulaire. Elle représente par exemple le point d'impact de l'activité de certains oncogènes comme le E1A, provenant des adenovirus. La protéine codée par cet oncogène d'origine virale (adénovirus) agit par formation d'un complexe avec p 105-Rb qui entraine l'inactivation de celle-ci. La protéine p53 a été identifiée comme la protéine cellulaire qui se couple à l'antigène T, produit transformant du virus simien SV 40. La protéine p53 peut être considérée comme la «gardienne du génôme ". A la suite d'une lésion affectant l'adn (rayonnement UV, exposition à un mutagène), elle agit comme un agent décisionnel qui provoquerait soit l'arrêt de la division cellulaire soit l'apoptose. L'ensemble de ces phénomènes contribue à la stabilité de l'intégrité génétique.
217 Par contre, les cellules tumorales présentant une mutation de p53 ne sont plus capables d'assurer ce maintien, la cellule ne recevant plus de signal d'arrêt de division. Son génome devient moins stable et susceptible d'accumuler des mutations diverses permettant l'émergence de clones cellulaires de malignité accrue. La p53 normale agit comme le produit d'un gène suppresseur récessif dans les modèles animaux. Du reste, le gène en cause est situé sur le chromosome 17, qui est le siège fréquent de délétions chromosomiques au cours de pathologies néoplasiques diverses.
218 Les connaissances actuelles sur la biologie du cancer conduisent donc à considérer la prolifération maligne comme résultant d'une succession d'évènements le plus souvent rares et aléatoires dont l'accumulation permet à une cellule d'échapper au contrôle génétique, humoral et immunologique exercé en permanence par l'organisme, et de se diviser de façon autonome, clonale et anarchique, acquérant au fur et à mesure de sa progression de nouvelles capacités de prolifération, d'extension, et de résistance aux tentatives de régulation endogènes et thérapeutiques. La multiplicité et l'intrication de ces mécanismes constitue la principale cause d'échec des traitements, qu'ils soient non spécifiques (cytotoxicité directe) ou plus spécifiques (stimulation des mécanismes de défense naturels de l'organisme).
219 le cancer est une maladie de l'adn et plusieurs mutations sont nécessaires au niveau du génome d'une cellule normale pour entraîner sa transformation en cellule maligne. Le cancer peut donc être considéré comme une maladie génétique à l'échelon de la cellule somatique. A l'échelon de la cellule germinale, La prédisposition génétique au cancer, dépend à la fois de facteurs liés à l'environnement, de facteurs génétique et des facteurs liés au simple hasard. La mise en évidence d'une prédisposition génétique au cancer a des applications pratiques qui peuvent en découler, en matière de prévention et de dépistage.
220 Retinoblastome Origine Moléculaire & Genetique Perte de fonction du gène RB gène suppresseur de tumeurs mutation Germinale du gène suppresseur de tumeurs «RB» Substitution: C T chromosome 13 inactivation de son expression Retinoblastome héréditaire ou sporadique, Cancer infantile cause des tumeurs multiples impliquant les deux yeux, Ou une tumeur isolé au niveau d un oeil
221 Neurofibromatoses Origine Moléculaire & Genetique Perte de fonction du gène NF1 gène suppresseur de tumeurs locus du gène: sur le chromosome 17 Le gène NF1 produit: la Neurofibromine (une protéine activatrice de la GTPase) Maladie génétique autosomique & dominante Cause des neurofibromes multiples & une prédisposition au tumeurs du système nerveux
222 Interaction de la protéine P53 avec la double hélice d ADN gène P53: localisé sur chromosome 17 Code pour une protéine de 53 kd «DNA-binding nuclear protein» «Transcription factor» Facteur clef de la régulation du cycle cellulaire Perte de fonction de P53 Cause des types multiples de cancer
223 Thérapie Conventionnel du Cancer Traitements disponibles du cancer : Radiothérapie, chimiothérapie, immunothérapie & Chirurgie, Traitements disponibles sont non spécifique aux cellules cancéreuses & causent donc des effets secondaires importants; La thérapie conventionnel du cancer fait face à ces limites dans les cas de progressions incurable des tumeurs & le cancer généralisé. Médicaments anticancers Médicaments alkylants (Endoxan): perturbent la réplication & empechent les deux brins d ADN de s écarter.
224 Thérapie Génique du Cancer. Utilisation des gènes suppresseurs des tumeurs qui existent Naturellement dans la cellule ou des gènes d antitopoisomerases, Insertion de ces gènes dans des vecteurs de DNA, portant des promoteurs tissus spécifique, La construction génétique recombinante est introduite dans des cellules cancéreuses, Permet une expression spécifique de protéines anti cancéreuses dans des cellules cibles, Sans effets sur les cellules non carcino génique.
225 Gene delivery systems Retroviral vectors delivery : stratégie de choix pour les essaies cliniques de thérapie génique: Vecteurs retro viraux recombinants & non virulent Ces vecteurs portent des gènes anti tumoraux Produisent un ARN génomique viral & recombinant «Packaging» en virions Utilisation de la Reverse transcriptase virale afin de produire de l ADN double brin «ds DNA» dans les cellules cibles La dsdna recombinante intègre la chromatine des cellules cibles Les cellules cancéreuses transfectés ou transduites expriment le produit du gène anti tumoral délivré par le vecteur viral recombinant Moyen efficace de transfert de gènes Les systèmes Adenoviraux, sont les virus recombinants de transfert les plus utilisés lors des essais cliniques
226 Gene delivery systems Il existe des méthodes physique pour transférer un DNA dans les cellules : Lipofection «Ligand DNA conjugates» «Adenovirus-ligand DNA conjugates» injection directe de l ADN Complexes Liposome-DNA.
227 Essays de Thérapie géniques du cancer & des maladies génétiques, infectieuses et métaboliques Premier transferts de gènes ont été approuvés en 1989 «marker gene», Premier protocole fédéral de thérapie génique a été approuvé en 1990 pour le traitement de l (ADA ): Adenosine DeAminase deficiency (Une déficience héréditaire, recessive qui cause la «Severe Combined Immunodeficiency Syndrome SCID». Actuellement, plusieurs protocoles actifs sont en cours concernant la thérapie génique des tumeurs et du cancer ainsi que la thérapie génique de maladies génétiques, métaboliques & infectieuse. Exemples de protocoles actifs de thérapie génique: Mélanomes, Neuroblastomes, tumeurs du cerveau, du sein, Colorectal, Ovarien & Renale. SCID, Gaucher s disease, Cystic fibrosis, Hypercholesterolemia AIDS vaccines
228 NeoR/Til gene marking des cellules thérapeutiques Nouvelle approche, pour le traitement du cancer lors des essais cliniques implique: Tumor Infiltrating Lymphocytes TILs. Des lymphocytes modifiées génétiquement afin de produire une protéine anti tumorale «TNF» (Tumor Necrosis Factor). Ces lymphocytes modifiées sont re introduites dans l organisme du patient cancéreux, elles migrent vers la tumeur, infiltrent les cellules cancéreuses, produisent le «TNF» & facilitent le rétrécissement de la tumeur. TILs peuvent produire et transférer des cytokines (interleukin 2), qui stimulent le système immunitaire pour attaquer les cellules cancéreuses. Le transfert des TILs, dans les patients soufrant d un cancer avancé est actuellement approuvé en tant que protocole de traitement par génie génétique.
229 Catégories de thérapies géniques utilisées pour lutter contre le cancer. Introduction de gènes suppresseurs de tumeurs dans les cellules tumorales Déclencher ou augmenter la réaction immunitaire dans les cellules cancéreuses: Introduction de gènes de cytrokines dans les cellules tumorales Réintroduction des cellules génétiquement modifiée chez le patient Ainsi la production de Cytokine dans les cellules cancéreuses active leur immunité Augmenter l antigenecité des cellules tumorales: Introduire le gène de HLA-B7 dans les mélanomes malignes (via les vecteurs retroviraux, ou via le complexe DNA/Liposome) déclencher une forte réponse immunitaire allogénique au site tumoral.
230 Introduction of gènes suicide directement dans les tumeurs: Le gène de la thymidine kinase du virus herpes simplex «HSV-TK gene». Cette kinase virale rend les cellules susceptible à être tuées par «Ganciclovir». Transfert Préférentiel du gène HSV-TK à la masse tumorale éradication Complète de la masse tumorale par l effet bystander Protéger le système hematopoietique du patient des effets toxique de la chimiothérapie à haute dose: Introduction de gènes de résistance à la chimiothérapie : Le gène «MDR-1» code pour une protéine (P glycoprotéine), utilisé une pompe (a multi drug efflux pump) qui enlève les agents chimiothérapeutique des cellules.
231 La recherche continue afin de découvrire de nouveaux gènes & protéines naturels anticancer dans la cellule Gènes Anti topoisomerases : Bloque le déroulement de l ADN chromosomale Inhibition de la transcription & de l expression génétique Mort des cellules cancéreuses. Naïma Alami-Ouahabi and al: Clonage du gène de tshmg Expression de la protéine Recombinante & sa caractérisation Facteur nucléaire à activité Anti topoisomerase I : tshmg.
232 Biologie moléculaire des Virus, Viroïdes & Prions Biologie moléculaire des Virus Agent infectieux à Structure simple; Une particule = Virion Virion: Acide. Nucléique ADN ou ARN (retrovirus) Une capside externe, protéique &protectrice (Ou juste une nucléocapside ou nucléoïde ou core) Une enveloppe: protège dans certains cas la nucleocapside Pouvoir pathogène vira, Nécessitée cellule hôte, Lutte antiviral Antiviraux: -Aciclovir (Zovirax) inhibe la DNA polymerase du virus Herpex (analogue structural de deoxyguanosine = pseudonucléoside de synthèse) -AZT: Azidothymidine inhibe la reverse transcriptase du HIV (analogue structural de la thymine). -Interférons: protéines de 17 à 25 kdaltons, synthétisés par les leukocytes Lymphocytes T & fibroblastes en réponse aux infections virales. (Interféron empêche la réplication virale & hydrolyse l ARNm virale.
233 Virus de l hépatite Types: A, B, C, D, E; différents structures, modes de contamination & pouvoir pathogène HBV Virus à ADN, génome < 5 kb, Génome double brin partiel: Brin long (-) antisens = 3,2 kb Brin court (+) sens =longeur variable Génome circulaire, complémentarité partielle (200nt) à l extrémité 5 des deux brins. Génome particulier: Gènes chevauchants permettent à un petit génome 3,2 kb de coder pour plusieurs protéines Une sequence à 3 cadres ouvert de lecture (ORF): transcrit 3 RNA & traduit 3 protéines
234
235 Gènes codant & protéines de HBV Gènes codant protéines codée P DNA polymerase C (core) HBc prot de nucléocapside Pré C Hbe derive de HBc Pré S1 grande prot de l enveloppe Pré S2 moyenne prot de l envelop S courte prot de l enveloppe prot Majeur prots: S1, S2 & S ont activité antigénique de surface HBS (test ELISA) Région C & S = 1er cadre de leture Région X = 2ème cadre de lecture Région P = 3ème cadre de lecture
236
237
238 Réplication de HBV Réplication sans intégration dans le DNA de la cellule hôte. Virus à ADN Le transcrit RNA 3,5 kb traduit protéines: polymerase, HBc &Hbe Le même RNA 3,5 kb donne : RNA pregénomique qui donne DNA genomique Par reverse transcriptase.
239
240 Détection de HBV: Analyse du sang et détection de l Ag HBs avec l Ac anti HBs Détection de l ADN virale par PCR «Polymerase Chain Reaction» Prevention de HBV: Vaccins: Protéine Ag = HBs Péptide synthétique = oligopeptide antigénique de HBs Vaccins recombinant séparés par Génie génétique: Construction génétique: Vecteur, promoteur& gène S Introduction dans les levures, production de HBs non glycosylée Vaccin recombinant + immunogénique Construction génétique: Vecteur, promoteur sequences pré S2 & S Cellules dérivées d ovaires de hamster chinois (CHO) Produit HBs courte & moyenne glycosylé = augmente immunogénécité Essais de vaccins recombinant vivant = modifier génétiquement (non virulent).
241 Virus HCV Variante détectée grâce à la biologie moléculaire par séquençage Virus à ARN monobrin = 10 kb RNA génomique brin codant (+) Gènes codent pour protéines structurales nucléocapside, l enveloppe & les prots trans membranaires les gènes qui codent pour la réplicase Patient présence Ac Anti HCV = détéction tardive Détection précoce de l ARN virale Rt PCR
242
243 Virus HIV 1 Virus à RNA = Rétrovirus, RNA donne DNA par la Reverse transcriptase DNA viral intègre DNA de la cellule hôte C T4 = T CD4+
244
245
246
247
248 Gènes codant du HIV 1 3 gènes de structure: Gag (prot: interieur de env, majeur de capside & nucléocapside) pol (région code pour retrotranscriptase, integrase & protéase) Env (Glycoprotéine de l enveloppe) 6 gènes régulateurs: nef ( Negative regulatory factor = latence) Tat (Transcription trans activator) Rev (Regulatior of expression of virion protéines) Vif ( virion infectivity factor = augmente pouvoir infectieux) Vpr & Vpu ou Vpx
249
250
251 Reconnaissance & affinité pour les protéines de surface la c hôte Injection du RNA viral dans c hôte RNA viral donne DNA viral par reverse transcriptase Intégration de HIV dans DNA de C Hôte par les bouts LTR Latence virale séronégatif Expression DNA virale Dimérisation du RNA viral Sortie des particules virales par bourgeonnement Poursuite de la maturation des particules virales Période de Primoinvasion sujet séropositif HIV augmente apoptose T4, augmentation de DNAase donc de fragmentation de l ADN HIV diminue l immunité de l organisme AIDS = SIDA Test ELISA (Enzyme Linked Immuno essay) detecte présence d Ac anti HIV Test de confirmation: Western blot Tests tardifs Tests precoces : Rt PCR détection RNA viral, PCR détection DNA viral
252 Ciblage Moléculaire HIV Contrer la reconnaissance gp 120 / CD4+ (Analogues moléculaires structuraux de CD4+) Contrer l action de la reverse transcriptase (Analogues structuraux de nucléotides avec perte du 3 OH Donc blocage de polymérisation donc de transformation de RNA viral en DNA viral) AZT = Azidothymine DDC = Dideoxycytidine DDI = Dideoxyinosine Inhiber l action des Protéases (maturation des particules virales) par des inhibiteur: sequinavir, Ritonavir Bithérapie ( AZT + anti protéases) ou Trithérapie
253
254 Viroïdes Structure + Simple que le virus Une molécule de RNA non protégée par protéines Structure en bâtonnet permet de résister aux attaques par nucléases 2/3 de complémentarité de séquence de l ARN Résponsable de maladie chez les plantes Homme??
255 BIOLOGIE MOLECULAIRE DES PRIONS PRIONS: agents infectieux «Small proteinaceous infectious particles» Absence d acides nucléiques, Petite protéine, Atteinte du système nerveux, Encéphalopathie spongiforme, Cerveau à aspect spongieux. Chez animal: Scarpie = tremblante de mouton & encéphalopathie bovine Chez l homme: Kuru (cannibalisme) Creutzfeld-Jacob (syndrome, 1/ /an), mutations 200 «Glu-----Lys», 178 «Asp----Asn», répétition d 1 octapeptide. Gerstmann-straüssler-sheinker, mutations: 102 «Pro----Leu» & 117 «Ala----Val»
256 Mise en évidence dans le génome mammifère d un gène qui code pour une protéine: Pr P c = Prion endogène exprimée normalement dans le cerveau, Gène sur Chromosome 20, avec 2 exons dont un seul (ex: 2) est codant, Élimination à l extrémité N-terminale de la protéine d 1 seq signal (22aa), Protéine glycosylée PrPc cellulaire Liée à la surface externe des cellules (neurones) par un ancrage GPI = Glycosyl-phophatdyl-inositol Entierement dégradable par protéinase K
257 Protéine Prion infectieux: PrPsc qui dérive de PrPc, PrPsc 33-35: ancrage GPI mais localisation intracellulaire, Très résistantes aux attaques protéolytiques. Protéolyse partielle coupe 67 aa & laisse une protéine glycosylée infcectieuse de 141 aa : PrP Transmission de maladie: 10% à 15% hérédité Contamination: Hormones de croissance (cadavres) Greffes de cornée Instruments neurochirurgicaux Consommation d abats ou de viandes contaminéés
258 Hypothèse Prusiner: Conversion de PrPc ( structure en hélice α )en PrPsc (structure en Feuillet β) provoquée par: PrPsc. Formation d un hétéro dimère entre PrPc & PrPsc. Transformation de PrPc en PrPsc. Accumulation de PrPsc dans les lysosymes qui éclatent & les enzymes Lysosomales endommagent les cellules nerveuses. Les prions libérés attaquent d autres cellules.
259 OUTILS & TECHNIQUES DE BIOLOGIE MOLECULAIRE OUTILS DE BIOLOGIE MOLECULAIRE ENZYMES: VECTEURS: MATRICE: SONDES: Enzymes de restriction Ligases Nucléases polymerases/ Taq polymerase Séquanases Phosphatases/kinases Plasmidiques Viraux hybrides ADN, ARN, Protéines ADN, ARN, Oligonucléotides, Biopuces
260 Enzymes de restriction: Endonucléases d origine bactérienne, coupent l ADN à des séquences de reconnaissance, Nomenclature: Exemples: Eco RI Extraite de Escherichia coli RYB site reconnu: G / AATTC Sma I Extraite de Serratia marcescens site reconnu: CCC / GGG Pst I Extraite de Providencia stuartii site reconnu: CTGCA / G
261 Type de coupure: «Cohesive ends» = cos ends Blunt ends = coupure droite
262
263
264
265
266 TECHNIQUES DE BIOLOGIE MOLECULAIRE CLONAGE EXTRACTION ADN, ARN DIGESTION DE L ADN PAR LES ENZYMES DE RESRTICTION SEPARATION DE L ADN & L ARN SUR GEL D AGAROSE SEPARATION DES PROTEINES SUR GEL DE POLYACRYLAMIDE VISUALISATION DE L ADN: COLORATION AU BROMURE D ETHIDIUM TRANSFERT DE L ADN, L ARN & LES PROTEINES SUR MEMBRANES DE NITROCELLULOSE «SOUTHERN, NORTHERN & WESTERN BLOTS» MARQUAGE DES SONDES, HYBRIDATION & AUTORADIOGRAPHIE SEQUENÇAGE D ADN. PCR ORGANISMES TRANSGENIQUES EMPREINTES D ADN RFLPS HYBRIDATION IN SITU.
267
268
269
270
271
272

273
274 LA PCR EN TEMPS RÉEL techniques moléculaires de génotypage en temps réel
275
276
277 La Polymerase Chain Reaction PCR : un outil crucial en diagnostic moléculaire permet une détection sensible et spécifique des cibles d acides nucléiques.
278 Le suivie en temps réel de la PCR: accélération et une simplification considérable des procédures de laboratoire quantification, l amplification et la détection de mutations au niveau des séquences cibles.

279 Le système Light Cycler RD Le système Light Cycler Le LC* est un appareil fondé sur le principe de PCR quantitative en temps réel. Il permet donc la détection et la quantification de marqueurs fluorescents dont l émission est directement proportionnelle à la quantité d amplicons générés pendant la réaction de PCR. Les mesures sont effectuées en phase exponentielle, plutôt qu en point final Les systèmes de détection : -les agents se liant à l ADN double brin ( exp : SYBR Green I qui est un fluorophore capable de se lier au sillon mineur de l ADN double brin )et, -les sondes fluorescentes
280 Au début de l amplification,le mélange réactionnel contient de l ADN dénaturé,les amorces et le fluorophore non lié. Après le couplage des amorces,le fluorophore se lie au double brin,ce qui se traduit par une augmentation de la fluorescence. Pendant l étape d élongation, le nombre de molécules de fluorophores lié à l ADN synthétisé augmente, se traduisant par une augmentation de la fluorescence.
281 Les sondes fluorescentes : deux sondes ( Hybridization probes ) : Méthode de détection est basée sur le principe FRET (Fluorescence Resonance Energy Transfer): deux sondes oligonucléotidiques linéaires complémentaires à une séquence cible 1ère sonde, dite sonde d ancrage, bloquée à son extrémité 3 afin de prévenir son extension durant l étape d élongation et transporte en 3 un fluorochrome donneur ( fluorescéine ) qui émet une lumière verte à 530 nm lorsque excité par une source de lumière une deuxième sonde d hybridation marquée en 5 avec le LC Red 640 (fluorochrome accepteur ).
282 En solution, les deux sondes sont libres et séparées. Lorsque,lors de l étape d hybridation,les deux sondes se fixent à leur séquences respectives, elles se trouvent très proches l une de l autre, cette proximité permet le transfert énergétique de la Fluorescéine verte par le FRET au fluorochrome accepteur rouge... Et provoque son émission rouge détectée par le spectrofluorimètre. Pendant l étape d élongation, les sondes retournent de façon indépendante en solution ce qui supprime la fluorescence rouge.
283 Les courbes de fusion : Chaque produit d ADN double brin synthétisé à une température de fusion Tm spécifique définie comme étant la température à laquelle 50 % de l ADN est sous forme double brin et 50% sous forme simple brin. Tm dépend essentiellement de la longueur du fragment d ADN double brin et de la teneur en GC de ce fragment (et aussi du degré d homologie entre les deux brins d ADN Analyse des courbes de fusion utilisant le SYBR Green I : 1/ le SYBR Green I est fluorescent tant qu il est lié à l ADN double brin, 2/ quand la température augmente l ADN double brin se dissocie, ce qui entraîne la libération du SYBR Green dans le milieu et une diminution progressive de la fluorescence, 3/ lorsque 50% de l ADN double brin sont dissociés, la fluorescence chute brutalement, c est cette température qui correspond à la température de fusion du produit synthétisé. La température de fusion est obtenue, en traçant la dérivée première négative de la fluorescence en fonction de la température : elle correspond au maximum de la courbe

284
285 Analyse des mutations par le LightCycler : Par analyse des courbes de fusion en utilisant les sondes d hybridation(oligonucléotides): Les deux sondes sont normalement complémentaires à une région spécifique au niveau de la séquence amplifiée. Si le gène amplifié présente une mutation ponctuelle au niveau de cette région, on peut avoir un ou plusieurs mésappariements (mismatches). En effet la Tm ne dépend pas uniquement de la longueur et du contenu en GC, mais aussi du degré d homologie entre les deux brins d ADN. Les sondes d hybridation liés à l ADN dans un appariement parfait nécessitent une Tm supérieure pour les séparer que ceux liées à l ADN contenant des mésappariements qui le déstabilisent. Le mismatch réduit donc la Tm de l oligonucléotide,cette diminution peut être détectée grâce à l analyse des courbes de fusion.
286 On remarque que la cible non mutée (sauvage ) présente une Tm supérieure à celle des séquences mutées
287 FISH

288 FISH FDA-approved kit for HER-2 testing by FISH and follow the scoring and interpretation recommended by the manufacturer. In this procedure, the chromosome 17 centromere is marked with a green florescent signal and HER-2 gene with an orange florescent signal. A tumor is designated as positive for gene amplification if gene-tochromosome (17) ratio is >2.0
289 Microarrays: Biopuces

290 La biopuce à ADN est une plaque de verre ou de silicium sur laquelle sont gravées un grand nombre de microcuvettes. Sur chacune d entre elles, on accroche une séquence de bases d ADN, qui joue le rôle de sonde et qui est caractéristique d un gène, d une mutation ou d une maladie. On prélève alors de l ADN sur le patient et on le verse dans les microcuvettes. L ADN-sonde se liera avec l ADN du prélèvement si et seulement si les séquences sont complémentaires. On lave ensuite la biopuce. Pour permettre la lecture du résultat, on a préalablement accroché à l extrémité de l ADN du prélèvement une molécule fluorescente. Ainsi, dans les microcuvettes où il y a eu appariement, l ADN du prélèvement est resté accroché à l ADN-sonde et est visible à la lumière ultraviolette grâce à la molécule fluorescente. Dans les microcuvettes où il n y a pas eu d appariement, l ADN du prélèvement a été enlevé par le lavage et il n y a pas de signal lumineux.
CHAPITRE 3 LA SYNTHESE DES PROTEINES
 CHAITRE 3 LA SYNTHESE DES ROTEINES On sait qu un gène détient dans sa séquence nucléotidique, l information permettant la synthèse d un polypeptide. Ce dernier caractérisé par sa séquence d acides aminés
CHAITRE 3 LA SYNTHESE DES ROTEINES On sait qu un gène détient dans sa séquence nucléotidique, l information permettant la synthèse d un polypeptide. Ce dernier caractérisé par sa séquence d acides aminés
VI- Expression du génome
 VI- Expression du génome VI-1.- EXPRESSION DU GÉNOME- PRINCIPES GÉNÉRAUX DOGME CENTRAL Les gènes et l information génétique sont conservés sous forme d acides nucléiques La perpétuation à l identique de
VI- Expression du génome VI-1.- EXPRESSION DU GÉNOME- PRINCIPES GÉNÉRAUX DOGME CENTRAL Les gènes et l information génétique sont conservés sous forme d acides nucléiques La perpétuation à l identique de
TD de Biochimie 4 : Coloration.
 TD de Biochimie 4 : Coloration. Synthèse de l expérience 2 Les questions posées durant l expérience 2 Exposé sur les méthodes de coloration des molécules : Générique Spécifique Autres Questions Pourquoi
TD de Biochimie 4 : Coloration. Synthèse de l expérience 2 Les questions posées durant l expérience 2 Exposé sur les méthodes de coloration des molécules : Générique Spécifique Autres Questions Pourquoi
Chapitre 7 : Structure de la cellule Le noyau cellulaire
 UE2 : Structure générale de la cellule Chapitre 7 : Structure de la cellule Le noyau cellulaire Professeur Michel SEVE Année universitaire 2010/2011 Université Joseph Fourier de Grenoble - Tous droits
UE2 : Structure générale de la cellule Chapitre 7 : Structure de la cellule Le noyau cellulaire Professeur Michel SEVE Année universitaire 2010/2011 Université Joseph Fourier de Grenoble - Tous droits
ULBI 101 Biologie Cellulaire L1. Le Système Membranaire Interne
 ULBI 101 Biologie Cellulaire L1 Le Système Membranaire Interne De la nécessité d un SMI Le volume augmente comme le cube de la dimension linéaire, alors que la surface n'est augmentée que du carré Une
ULBI 101 Biologie Cellulaire L1 Le Système Membranaire Interne De la nécessité d un SMI Le volume augmente comme le cube de la dimension linéaire, alors que la surface n'est augmentée que du carré Une
Contrôle de l'expression génétique : Les régulations post-transcriptionnelles
 Contrôle de l'expression génétique : Les régulations post-transcriptionnelles http://perso.univ-rennes1.fr/serge.hardy/ utilisateur : biochimie mot de passe : 2007 L'ARNm, simple intermédiaire entre le
Contrôle de l'expression génétique : Les régulations post-transcriptionnelles http://perso.univ-rennes1.fr/serge.hardy/ utilisateur : biochimie mot de passe : 2007 L'ARNm, simple intermédiaire entre le
Séquence 2. L expression du patrimoine génétique. Sommaire
 Séquence 2 L expression du patrimoine génétique Sommaire 1. La synthèse des protéines 2. Phénotypes, génotypes et environnement Synthèse de la séquence 2 Exercices de la séquence 2 Glossaire des séquences
Séquence 2 L expression du patrimoine génétique Sommaire 1. La synthèse des protéines 2. Phénotypes, génotypes et environnement Synthèse de la séquence 2 Exercices de la séquence 2 Glossaire des séquences
Bases moléculaires des mutations Marc Jeanpierre
 Bases moléculaires des mutations Marc Jeanpierre Chaque enfant qui naît hérite de 10 à 30 nouvelles mutations ponctuelles. L essentiel des ces mutations sont heureusement des variations neutres de séquence
Bases moléculaires des mutations Marc Jeanpierre Chaque enfant qui naît hérite de 10 à 30 nouvelles mutations ponctuelles. L essentiel des ces mutations sont heureusement des variations neutres de séquence
Univers Vivant Révision. Notions STE
 Univers Vivant Révision Notions STE Chap. 13) L Écologie 1) a) Qu est-ce que l empreinte écologique? L empreinte écologique correspond à la surface terrestre et aquatique totale nécessaire à un individu,
Univers Vivant Révision Notions STE Chap. 13) L Écologie 1) a) Qu est-ce que l empreinte écologique? L empreinte écologique correspond à la surface terrestre et aquatique totale nécessaire à un individu,
Respiration Mitochondriale
 Université Pierre et Marie Curie Respiration Mitochondriale Objectifs au cours de Révisions Biochimie PCEM2 Révisions Biochimie Métabolique 2004-2005 Pr. A. Raisonnier (alain.raisonnier@upmc.fr) Mise à
Université Pierre et Marie Curie Respiration Mitochondriale Objectifs au cours de Révisions Biochimie PCEM2 Révisions Biochimie Métabolique 2004-2005 Pr. A. Raisonnier (alain.raisonnier@upmc.fr) Mise à
Les OGM. 5 décembre 2008. Nicole Mounier
 Les OGM 5 décembre 2008 Nicole Mounier Université Claude Bernard Lyon 1 CGMC, bâtiment Gregor Mendel 43, boulevard du 11 Novembre 1918 69622 Villeurbanne Cedex OGM Organismes Génétiquement Modifiés Transfert
Les OGM 5 décembre 2008 Nicole Mounier Université Claude Bernard Lyon 1 CGMC, bâtiment Gregor Mendel 43, boulevard du 11 Novembre 1918 69622 Villeurbanne Cedex OGM Organismes Génétiquement Modifiés Transfert
Séquence 1. Reproduction conforme de la cellule et réplication de l ADN Variabilité génétique et mutation de l ADN
 Séquence 1 Reproduction conforme de la cellule et réplication de l ADN Variabilité génétique et mutation de l ADN Sommaire 1. Reproduction conforme de la cellule et réplication de l ADN 2. Variabilité
Séquence 1 Reproduction conforme de la cellule et réplication de l ADN Variabilité génétique et mutation de l ADN Sommaire 1. Reproduction conforme de la cellule et réplication de l ADN 2. Variabilité
Biomarqueurs en Cancérologie
 Biomarqueurs en Cancérologie Définition, détermination, usage Biomarqueurs et Cancer: définition Anomalie(s) quantitative(s) ou qualitative(s) Indicative(s) ou caractéristique(s) d un cancer ou de certaines
Biomarqueurs en Cancérologie Définition, détermination, usage Biomarqueurs et Cancer: définition Anomalie(s) quantitative(s) ou qualitative(s) Indicative(s) ou caractéristique(s) d un cancer ou de certaines
Tableau 1. Liste (non exhaustive) des protéines se localisant dans les P-Bodies
 des protéines se localisant dans les P-Bodies") NOM FONCTION EFFET DE L ABSENCE OU DE REFERENCES LA SUREXPRESSION SUR LES P- BODIES XRN1 exonucléase 5 3 Absence : augmente la taille et le Bashkirov et al. 1997 Shet et Parker 2003 Cougot nombre des P-bodies
NOM FONCTION EFFET DE L ABSENCE OU DE REFERENCES LA SUREXPRESSION SUR LES P- BODIES XRN1 exonucléase 5 3 Absence : augmente la taille et le Bashkirov et al. 1997 Shet et Parker 2003 Cougot nombre des P-bodies
Travaux dirigés de Microbiologie Master I Sciences des Génomes et des Organismes Janvier 2015
 Andrew Tolonen atolonen@genoscope.cns.fr Travaux dirigés de Microbiologie Master I Sciences des Génomes et des Organismes Janvier 2015 A- Généralités I- La vie sur terre telle que nous la connaissons ne
Andrew Tolonen atolonen@genoscope.cns.fr Travaux dirigés de Microbiologie Master I Sciences des Génomes et des Organismes Janvier 2015 A- Généralités I- La vie sur terre telle que nous la connaissons ne
Module 5 La maturation de l ARN et le contrôle post-transcriptionnel chez les eucaryotes
 Module 5 La maturation de l ARN et le contrôle post-transcriptionnel chez les eucaryotes Où trouver l'information complémentaire? MCB -11, GVII-5, 22, 23. La maturation des ARNm chez les eucaryotes Les
Module 5 La maturation de l ARN et le contrôle post-transcriptionnel chez les eucaryotes Où trouver l'information complémentaire? MCB -11, GVII-5, 22, 23. La maturation des ARNm chez les eucaryotes Les
Cellules procaryotes Service histologie Pr.k.mebarek
 Cellules procaryotes Service histologie Pr.k.mebarek I) Les cellules procaryotes II) Les cellules eucaryotes o 1) Caractéristiques générales des cellules eucaryotes o 2) Organisation des cellules eucaryotes
Cellules procaryotes Service histologie Pr.k.mebarek I) Les cellules procaryotes II) Les cellules eucaryotes o 1) Caractéristiques générales des cellules eucaryotes o 2) Organisation des cellules eucaryotes
Dr E. CHEVRET UE2.1 2013-2014. Aperçu général sur l architecture et les fonctions cellulaires
 Aperçu général sur l architecture et les fonctions cellulaires I. Introduction II. Les microscopes 1. Le microscope optique 2. Le microscope à fluorescence 3. Le microscope confocal 4. Le microscope électronique
Aperçu général sur l architecture et les fonctions cellulaires I. Introduction II. Les microscopes 1. Le microscope optique 2. Le microscope à fluorescence 3. Le microscope confocal 4. Le microscope électronique
Contrôle de l'expression génétique :
 Contrôle de l'expression génétique : Les régulations post-transcriptionnelles L'ARNm, simple intermédiaire entre le génome et les protéines? gène protéine L'ARNm, simple intermédiaire entre le génome et
Contrôle de l'expression génétique : Les régulations post-transcriptionnelles L'ARNm, simple intermédiaire entre le génome et les protéines? gène protéine L'ARNm, simple intermédiaire entre le génome et
INTRODUCTION À L'ENZYMOLOGIE
 INTRODUCTION À L'ENZYMOLOGIE Les enzymes sont des macromolécules spécialisées qui - catalysent les réactions biologiques - transforment différentes formes d'énergie. Les enzymes diffèrent des catalyseurs
INTRODUCTION À L'ENZYMOLOGIE Les enzymes sont des macromolécules spécialisées qui - catalysent les réactions biologiques - transforment différentes formes d'énergie. Les enzymes diffèrent des catalyseurs
Partie Observer : Ondes et matière CHAP 04-ACT/DOC Analyse spectrale : Spectroscopies IR et RMN
 Partie Observer : Ondes et matière CHAP 04-ACT/DOC Analyse spectrale : Spectroscopies IR et RMN Objectifs : Exploiter un spectre infrarouge pour déterminer des groupes caractéristiques Relier un spectre
Partie Observer : Ondes et matière CHAP 04-ACT/DOC Analyse spectrale : Spectroscopies IR et RMN Objectifs : Exploiter un spectre infrarouge pour déterminer des groupes caractéristiques Relier un spectre
Les outils de génétique moléculaire Les techniques liées aux acides nucléiques
 Les outils de génétique moléculaire Les techniques liées aux acides nucléiques Sommaire Preparation des acides nucléiques Extraction / purification Les enzymes agissant sur les acides nucléiques Les enzymes
Les outils de génétique moléculaire Les techniques liées aux acides nucléiques Sommaire Preparation des acides nucléiques Extraction / purification Les enzymes agissant sur les acides nucléiques Les enzymes
4 : MÉTHODES D ANALYSE UTILISÉES EN ÉCOLOGIE MICROBIENNE
 4 : MÉTHODES D ANALYSE UTILISÉES EN ÉCOLOGIE MICROBIENNE L écologie microbienne (ou étude des micro-organismes de l environnement) étudie : les relations entre les différentes populations de micro-organismes
4 : MÉTHODES D ANALYSE UTILISÉES EN ÉCOLOGIE MICROBIENNE L écologie microbienne (ou étude des micro-organismes de l environnement) étudie : les relations entre les différentes populations de micro-organismes
MAB Solut. vos projets. MABLife Génopole Campus 1 5 rue Henri Desbruères 91030 Evry Cedex. www.mabsolut.com. intervient à chaque étape de
 Mabsolut-DEF-HI:Mise en page 1 17/11/11 17:45 Page1 le département prestataire de services de MABLife de la conception à la validation MAB Solut intervient à chaque étape de vos projets Création d anticorps
Mabsolut-DEF-HI:Mise en page 1 17/11/11 17:45 Page1 le département prestataire de services de MABLife de la conception à la validation MAB Solut intervient à chaque étape de vos projets Création d anticorps
IMMUNOLOGIE. La spécificité des immunoglobulines et des récepteurs T. Informations scientifiques
 IMMUNOLOGIE La spécificité des immunoglobulines et des récepteurs T Informations scientifiques L infection par le VIH entraîne des réactions immunitaires de l organisme qui se traduisent par la production
IMMUNOLOGIE La spécificité des immunoglobulines et des récepteurs T Informations scientifiques L infection par le VIH entraîne des réactions immunitaires de l organisme qui se traduisent par la production
ACIDES BASES. Chap.5 SPIESS
 ACIDES BASES «Je ne crois pas que l on me conteste que l acide n ait des pointes Il ne faut que le goûter pour tomber dans ce sentiment car il fait des picotements sur la langue.» Notion d activité et
ACIDES BASES «Je ne crois pas que l on me conteste que l acide n ait des pointes Il ne faut que le goûter pour tomber dans ce sentiment car il fait des picotements sur la langue.» Notion d activité et
Compétence 3-1 S EXPRIMER A L ECRIT Fiche professeur
 Compétence 3-1 S EXPRIMER A L ECRIT Fiche professeur Nature de l activité : Réaliser 3 types de productions écrites (réécriture de notes, production d une synthèse de documents, production d une argumentation)
Compétence 3-1 S EXPRIMER A L ECRIT Fiche professeur Nature de l activité : Réaliser 3 types de productions écrites (réécriture de notes, production d une synthèse de documents, production d une argumentation)
CHAPITRE 3 LES TASQ COMME PLATEFORME POUR LA CATALYSE PSEUDO-ENZYMATIQUE
 L. Stefan Chapitre 3 CHAPITRE 3 LES TASQ COMME PLATEFORME POUR LA CATALYSE PSEUDO-ENZYMATIQUE Nous avons démontré dans le chapitre précédent que les TASQ hydrosolubles développés au cours de cette thèse
L. Stefan Chapitre 3 CHAPITRE 3 LES TASQ COMME PLATEFORME POUR LA CATALYSE PSEUDO-ENZYMATIQUE Nous avons démontré dans le chapitre précédent que les TASQ hydrosolubles développés au cours de cette thèse
Hépatite chronique B Moyens thérapeutiques
 Hépatite chronique B Moyens thérapeutiques Dr Olfa BAHRI Laboratoire de Virologie Clinique Institut Pasteur de Tunis INTRODUCTION Plus de 300. 10 6 porteurs chroniques de VHB dans le monde Hépatite chronique
Hépatite chronique B Moyens thérapeutiques Dr Olfa BAHRI Laboratoire de Virologie Clinique Institut Pasteur de Tunis INTRODUCTION Plus de 300. 10 6 porteurs chroniques de VHB dans le monde Hépatite chronique
ANTIVIRAUX ET INTERFERONS LES ANTIVIRAUX
 ANTIVIRAUX ET INTERFERONS LES ANTIVIRAUX La thérapeutique antivirale progresse avec lenteur car elle s'attaque à des micro-organismes ne se multipliant qu'à l'intérieur des cellules vivantes dont ils détournent
ANTIVIRAUX ET INTERFERONS LES ANTIVIRAUX La thérapeutique antivirale progresse avec lenteur car elle s'attaque à des micro-organismes ne se multipliant qu'à l'intérieur des cellules vivantes dont ils détournent
Compléments - Chapitre 5 Spectroscopie
 ompléments - hapitre 5 Spectroscopie Spectroscopie par résonance magnétique nucléaire (RMN 13 ) Tandis que la spectroscopie RMN 1 H fournit des données sur la disposition des atomes d'hydrogène dans une
ompléments - hapitre 5 Spectroscopie Spectroscopie par résonance magnétique nucléaire (RMN 13 ) Tandis que la spectroscopie RMN 1 H fournit des données sur la disposition des atomes d'hydrogène dans une
Annales de Biologie Cellulaire QCM (niveau SVT 1 er année)
") Annales de Biologie Cellulaire QCM (niveau SVT 1 er année) Equipe pédagogique Université Bordeaux-1 Didier Morin, Michel Moenner, Sophie North, Gérard Tramu et IJsbrand Kramer Contact : i.kramer@iecb.u-bordeaux.fr
Annales de Biologie Cellulaire QCM (niveau SVT 1 er année) Equipe pédagogique Université Bordeaux-1 Didier Morin, Michel Moenner, Sophie North, Gérard Tramu et IJsbrand Kramer Contact : i.kramer@iecb.u-bordeaux.fr
Luca : à la recherche du plus proche ancêtre commun universel Patrick Forterre, Simonetta Gribaldo, Céline Brochier
 MEDECINE/SCIENCES 2005 ; 21 :???-??? > L application des méthodes de la biologie moléculaire à l étude des premières étapes de l évolution a montré que le monde vivant pouvait être divisé en trois domaines
MEDECINE/SCIENCES 2005 ; 21 :???-??? > L application des méthodes de la biologie moléculaire à l étude des premières étapes de l évolution a montré que le monde vivant pouvait être divisé en trois domaines
Comprendre l Univers grâce aux messages de la lumière
 Seconde / P4 Comprendre l Univers grâce aux messages de la lumière 1/ EXPLORATION DE L UNIVERS Dans notre environnement quotidien, les dimensions, les distances sont à l échelle humaine : quelques mètres,
Seconde / P4 Comprendre l Univers grâce aux messages de la lumière 1/ EXPLORATION DE L UNIVERS Dans notre environnement quotidien, les dimensions, les distances sont à l échelle humaine : quelques mètres,
Les débuts de la génétique
 HPITRE 9 DES DÉBTS DE L ÉNÉTIQE X ENJEX TELS DES BIOTEHNOLOIES 1 Les débuts de la génétique est avec les travaux de regor Mendel vers la fin du XIX e siècle que furent posées les bases de la génétique.
HPITRE 9 DES DÉBTS DE L ÉNÉTIQE X ENJEX TELS DES BIOTEHNOLOIES 1 Les débuts de la génétique est avec les travaux de regor Mendel vers la fin du XIX e siècle que furent posées les bases de la génétique.
EXERCICES : MECANISMES DE L IMMUNITE : pages 406 407 408 409 410
 EXERCICES : MECANISMES DE L IMMUNITE : pages 406 407 408 409 410 EXERCICE 1 PAGE 406 : EXPERIENCES A INTERPRETER Question : rôles respectifs du thymus et de la moelle osseuse dans la production des lymphocytes.
EXERCICES : MECANISMES DE L IMMUNITE : pages 406 407 408 409 410 EXERCICE 1 PAGE 406 : EXPERIENCES A INTERPRETER Question : rôles respectifs du thymus et de la moelle osseuse dans la production des lymphocytes.
Université de Montréal. Développement d outils pour l analyse de données de ChIP-seq et l identification des facteurs de transcription
 Université de Montréal Développement d outils pour l analyse de données de ChIP-seq et l identification des facteurs de transcription par Eloi Mercier Département de bioinformatique Faculté de médecine
Université de Montréal Développement d outils pour l analyse de données de ChIP-seq et l identification des facteurs de transcription par Eloi Mercier Département de bioinformatique Faculté de médecine
Utilisation des substrats énergétiques
 Utilisation des substrats énergétiques Collège des Enseignants de Nutrition Date de création du document 2010-2011 Table des matières I Les organes et les substrats... 3 I.1 Les substrats énergétiques...
Utilisation des substrats énergétiques Collège des Enseignants de Nutrition Date de création du document 2010-2011 Table des matières I Les organes et les substrats... 3 I.1 Les substrats énergétiques...
- pellicule de fruits qui a un rôle de prévention contre l'évaporation, le développement de moisissures et l'infection par des parasites
 LES LIPIDES Quelles Sont les Idées Clés? Les lipides sont les huiles et les graisses de la vie courante. Ils sont insolubles dans l eau. Pour les synthétiser, une réaction : l Estérification. Pour les
LES LIPIDES Quelles Sont les Idées Clés? Les lipides sont les huiles et les graisses de la vie courante. Ils sont insolubles dans l eau. Pour les synthétiser, une réaction : l Estérification. Pour les
Université d Evry-Val d Essonne Ecole Doctorale des Génomes Aux Organismes. Thèse
 Université d Evry-Val d Essonne Ecole Doctorale des Génomes Aux Organismes Thèse Présentée pour obtenir le grade de Docteur en sciences de l université d Evry-Val d Essonne Spécialité Bioinformatique par
Université d Evry-Val d Essonne Ecole Doctorale des Génomes Aux Organismes Thèse Présentée pour obtenir le grade de Docteur en sciences de l université d Evry-Val d Essonne Spécialité Bioinformatique par
I. La levure Saccharomyces cerevisiae: mode de vie
 LES LEVURES UE «levures» -5 avril: généralités (MN Simon) -6 avril: analyse génétique (MN Simon) -6 avril: Cycle cellulaire I: la réplication (E. bailly) -7 avril: Cycle cellulaire II: la mitose (E. Bailly)
LES LEVURES UE «levures» -5 avril: généralités (MN Simon) -6 avril: analyse génétique (MN Simon) -6 avril: Cycle cellulaire I: la réplication (E. bailly) -7 avril: Cycle cellulaire II: la mitose (E. Bailly)
2 C est quoi la chimie?
 PARTIE 1 AVANT LA CHIMIE VERTE... 2 C est quoi la chimie? L inconnu étant source d angoisse, nous allons essayer de définir les grands domaines de la chimie pour mieux la connaître, l appréhender et donc
PARTIE 1 AVANT LA CHIMIE VERTE... 2 C est quoi la chimie? L inconnu étant source d angoisse, nous allons essayer de définir les grands domaines de la chimie pour mieux la connaître, l appréhender et donc
Formavie 2010. 2 Différentes versions du format PDB...3. 3 Les champs dans les fichiers PDB...4. 4 Le champ «ATOM»...5. 6 Limites du format PDB...
 Formavie 2010 Les fichiers PDB Les fichiers PDB contiennent les informations qui vont permettre à des logiciels de visualisation moléculaire (ex : RasTop ou Jmol) d afficher les molécules. Un fichier au
Formavie 2010 Les fichiers PDB Les fichiers PDB contiennent les informations qui vont permettre à des logiciels de visualisation moléculaire (ex : RasTop ou Jmol) d afficher les molécules. Un fichier au
UE : GENE 2208. Responsable : Enseignant : ECUE 1. Enseignant : ECUE 2. Dr COULIBALY Foungotin Hamidou
 UE : GENE 2208 Responsable : Enseignant : ECUE 1 Enseignant : ECUE 2 Dr COULIBALY Foungotin Hamidou Spécialités : Génétique Humaine Biologie de la Procréation Cytogénétique Spermiologie Essais Cliniques
UE : GENE 2208 Responsable : Enseignant : ECUE 1 Enseignant : ECUE 2 Dr COULIBALY Foungotin Hamidou Spécialités : Génétique Humaine Biologie de la Procréation Cytogénétique Spermiologie Essais Cliniques
LA MITOSE CUEEP - USTL DÉPARTEMENT SCIENCES BAHIJA DELATTRE
 Biologie LA MITOSE CUEEP - USTL DÉPARTEMENT SCIENCES BAHIJA DELATTRE Février 2006 I. L'INTRODUCTION Chaque cellule d'un organisme supérieur provient de la multiplication d'une cellule préexistante (cellule
Biologie LA MITOSE CUEEP - USTL DÉPARTEMENT SCIENCES BAHIJA DELATTRE Février 2006 I. L'INTRODUCTION Chaque cellule d'un organisme supérieur provient de la multiplication d'une cellule préexistante (cellule
Chapitre II La régulation de la glycémie
 Chapitre II La régulation de la glycémie Glycémie : concentration de glucose dans le sang valeur proche de 1g/L Hypoglycémie : perte de connaissance, troubles de la vue, voire coma. Hyperglycémie chronique
Chapitre II La régulation de la glycémie Glycémie : concentration de glucose dans le sang valeur proche de 1g/L Hypoglycémie : perte de connaissance, troubles de la vue, voire coma. Hyperglycémie chronique
Structure quantique cohérente et incohérente de l eau liquide
 Structure quantique cohérente et incohérente de l eau liquide Prof. Marc HENRY Chimie Moléculaire du Solide Institut Le Bel, 4, Rue Blaise Pascal 67070 Strasbourg Cedex, France Tél: 03.68.85.15.00 e-mail:
Structure quantique cohérente et incohérente de l eau liquide Prof. Marc HENRY Chimie Moléculaire du Solide Institut Le Bel, 4, Rue Blaise Pascal 67070 Strasbourg Cedex, France Tél: 03.68.85.15.00 e-mail:
Production d une protéine recombinante
 99 Production d une protéine recombinante Lic. B. PIRSON Lic. J-M. SERONT ISICHt - Mons Production de la protéine recombinante GFP (Green Fluorescent Protein d Aequoria victoria) par une bactérie ( E.
99 Production d une protéine recombinante Lic. B. PIRSON Lic. J-M. SERONT ISICHt - Mons Production de la protéine recombinante GFP (Green Fluorescent Protein d Aequoria victoria) par une bactérie ( E.
Stéphane Priet. Research Group Viral Replicases: Structure, Function & Drug Design. Architecture et Fonction des Macromolécules Biologiques (AFMB)
") Stéphane Priet Research Group Viral Replicases: Structure, Function & Drug Design Architecture et Fonction des Macromolécules Biologiques (AFMB) CNRS UMR 7257- Aix-Marseille Université stephane.priet@afmb.univ-mrs.fr
Stéphane Priet Research Group Viral Replicases: Structure, Function & Drug Design Architecture et Fonction des Macromolécules Biologiques (AFMB) CNRS UMR 7257- Aix-Marseille Université stephane.priet@afmb.univ-mrs.fr
COUSIN Fabien KERGOURLAY Gilles. 19 octobre 2007. de l hôte par les. Master 2 MFA Responsable : UE Incidence des paramètres environnementaux
 COUSIN Fabien KERGOURLAY Gilles 19 octobre 2007 Inhibition des défenses de l hôte par les bactéries pathogènes Master 2 MFA Responsable : UE Incidence des paramètres environnementaux Gwennola ERMEL I Détection
COUSIN Fabien KERGOURLAY Gilles 19 octobre 2007 Inhibition des défenses de l hôte par les bactéries pathogènes Master 2 MFA Responsable : UE Incidence des paramètres environnementaux Gwennola ERMEL I Détection
Molécules et Liaison chimique
 Molécules et liaison chimique Molécules et Liaison chimique La liaison dans La liaison dans Le point de vue classique: l approche l de deux atomes d hydrogd hydrogènes R -0,9-1 0 0,5 1 1,5,5 3 3,5 4 R
Molécules et liaison chimique Molécules et Liaison chimique La liaison dans La liaison dans Le point de vue classique: l approche l de deux atomes d hydrogd hydrogènes R -0,9-1 0 0,5 1 1,5,5 3 3,5 4 R
L universalité et la variabilité de l ADN
 L universalité et la variabilité de l DN Unité 4 3 μm hromosomes observés en microscopie électronique à balayage. Les chromosomes, présents dans le noyau, sont constitués d acide désoxyribonucléique (DN).
L universalité et la variabilité de l DN Unité 4 3 μm hromosomes observés en microscopie électronique à balayage. Les chromosomes, présents dans le noyau, sont constitués d acide désoxyribonucléique (DN).
De la physico-chimie à la radiobiologie: nouveaux acquis (I)
") De la physico-chimie à la radiobiologie: nouveaux acquis (I) Collaboration: - Laboratoire de Radiotoxicologie et Oncologie (L. Sabatier) CEA, DSV - Laboratoire de Génotoxicité et Modulation de l Expression
De la physico-chimie à la radiobiologie: nouveaux acquis (I) Collaboration: - Laboratoire de Radiotoxicologie et Oncologie (L. Sabatier) CEA, DSV - Laboratoire de Génotoxicité et Modulation de l Expression
a. Fusion et énergie de liaison des noyaux b. La barrière Coulombienne c. Effet tunnel & pic de Gamov
 V. Les réactions r thermonucléaires 1. Principes a. Fusion et énergie de liaison des noyaux b. La barrière Coulombienne c. Effet tunnel & pic de Gamov 2. Taux de réactions r thermonucléaires a. Les sections
V. Les réactions r thermonucléaires 1. Principes a. Fusion et énergie de liaison des noyaux b. La barrière Coulombienne c. Effet tunnel & pic de Gamov 2. Taux de réactions r thermonucléaires a. Les sections
La reconnaissance moléculaire: la base du design rationnel Modélisation moléculaire: Introduction Hiver 2006
 La reconnaissance moléculaire: la base du design rationnel En 1890 Emil Fisher a proposé le modèle "serrure et clé" pour expliquer la façon de fonctionner des systèmes biologiques. Un substrat rentre et
La reconnaissance moléculaire: la base du design rationnel En 1890 Emil Fisher a proposé le modèle "serrure et clé" pour expliquer la façon de fonctionner des systèmes biologiques. Un substrat rentre et
Transport des gaz dans le sang
 UE3-2 - Physiologie Physiologie Respiratoire Chapitre 9 : Transport des gaz dans le sang Docteur Sandrine LAUNOIS-ROLLINAT Année universitaire 2010/2011 Université Joseph Fourier de Grenoble - Tous droits
UE3-2 - Physiologie Physiologie Respiratoire Chapitre 9 : Transport des gaz dans le sang Docteur Sandrine LAUNOIS-ROLLINAT Année universitaire 2010/2011 Université Joseph Fourier de Grenoble - Tous droits
IlliS. Le ribosome bactérien : structure et fonctions SYNTHÈSE
 IlliS SYNTHÈSE médecine! sciences 1989 ; 5: 662-69 Le ribosome bactérien : structure et fonctions Le ribosome est une particule ribonucléoprotéique, complexe jouant le rôle d'«unité centrale» de la synthèse
IlliS SYNTHÈSE médecine! sciences 1989 ; 5: 662-69 Le ribosome bactérien : structure et fonctions Le ribosome est une particule ribonucléoprotéique, complexe jouant le rôle d'«unité centrale» de la synthèse
Insulinothérapie et diabète de type 1
 Insulinothérapie et diabète de type 1 Introduction: la molécule d insuline L instauration de l insulinothérapie Dispositif d administration de l insuline Les propriétés de l insuline Insuline et schémas
Insulinothérapie et diabète de type 1 Introduction: la molécule d insuline L instauration de l insulinothérapie Dispositif d administration de l insuline Les propriétés de l insuline Insuline et schémas
3: Clonage d un gène dans un plasmide
 3: Clonage d un gène dans un plasmide Le clonage moléculaire est une des bases du génie génétique. Il consiste à insérer un fragment d'adn (dénommé insert) dans un vecteur approprié comme un plasmide par
3: Clonage d un gène dans un plasmide Le clonage moléculaire est une des bases du génie génétique. Il consiste à insérer un fragment d'adn (dénommé insert) dans un vecteur approprié comme un plasmide par
1. Principes de biochimie générale. A. Bioénergétique et dynamique. a) Intro: Les mitochondries passent leur temps à fabriquer de l énergie.
 Intro: Les mitochondries passent leur temps à fabriquer de l énergie.") RESPIRATION MITOCHONDRIALE 1. Principes de biochimie générale. A. Bioénergétique et dynamique. a) Intro: Les mitochondries passent leur temps à fabriquer de l énergie. b) Rappels: L énergie chimique sert
RESPIRATION MITOCHONDRIALE 1. Principes de biochimie générale. A. Bioénergétique et dynamique. a) Intro: Les mitochondries passent leur temps à fabriquer de l énergie. b) Rappels: L énergie chimique sert
Le rôle de l endocytose dans les processus pathologiques
 UE7 Cours n 9 C. LAMAZE 24.11.11 Elise GODEAU (partie1) Guillaume MERGENTHALER (partie2) Le rôle de l endocytose dans les processus pathologiques SOMMAIRE : I. L endocytose à récepteurs : la voie des clathrines
UE7 Cours n 9 C. LAMAZE 24.11.11 Elise GODEAU (partie1) Guillaume MERGENTHALER (partie2) Le rôle de l endocytose dans les processus pathologiques SOMMAIRE : I. L endocytose à récepteurs : la voie des clathrines
Partie 1. Addition nucléophile suivie d élimination (A N + E) 1.1. Réactivité électrophile des acides carboxyliques et groupes dérivés
 1.1. Réactivité électrophile des acides carboxyliques et groupes dérivés") Molécules et matériaux organiques Partie 1. Addition nucléophile suivie d élimination (A N + E) 1.1. Réactivité électrophile des acides carboxyliques et groupes dérivés bjectifs du chapitre Notions à connaître
Molécules et matériaux organiques Partie 1. Addition nucléophile suivie d élimination (A N + E) 1.1. Réactivité électrophile des acides carboxyliques et groupes dérivés bjectifs du chapitre Notions à connaître
Antiviraux ciblant les polymérases virales
 Antiviraux ciblant les polymérases virales Barbara Selisko Barbara.Selisko@afmb.univ-mrs.fr Research Group Viral Replicases: Structure, Function & Drug Design Head: Bruno Canard Architecture et Fonction
Antiviraux ciblant les polymérases virales Barbara Selisko Barbara.Selisko@afmb.univ-mrs.fr Research Group Viral Replicases: Structure, Function & Drug Design Head: Bruno Canard Architecture et Fonction
Journée SITG, Genève 15 octobre 2013. Nicolas Lachance-Bernard M.ATDR Doctorant, Laboratoire de systèmes d information géographique
 Monitorint spatio-temporel intégré de la mobilité urbaine Monitoring spatio-temporel de l ADN urbain Une réponse aux défis, problèmes, enjeux et risques des milieux urbains Nicolas Lachance-Bernard M.ATDR
Monitorint spatio-temporel intégré de la mobilité urbaine Monitoring spatio-temporel de l ADN urbain Une réponse aux défis, problèmes, enjeux et risques des milieux urbains Nicolas Lachance-Bernard M.ATDR
Pemetrexed, pionnier de la chimiothérapie histoguidée. Dr Olivier CASTELNAU Institut Arnault TZANCK ST Laurent du Var
 Pemetrexed, pionnier de la chimiothérapie histoguidée Dr Olivier CASTELNAU Institut Arnault TZANCK ST Laurent du Var SFPO Octobre 2009 EPIDEMIOLOGIE Incidence : 1.35 M par an dans le monde (12,4%) 28 000
Pemetrexed, pionnier de la chimiothérapie histoguidée Dr Olivier CASTELNAU Institut Arnault TZANCK ST Laurent du Var SFPO Octobre 2009 EPIDEMIOLOGIE Incidence : 1.35 M par an dans le monde (12,4%) 28 000
Information génétique
 chapitre 3 Information génétique et division cellulaire L étude de la division cellulaire est abordée pour découvrir comment est transmise et conservée l information génétique portée par les chromosomes.
chapitre 3 Information génétique et division cellulaire L étude de la division cellulaire est abordée pour découvrir comment est transmise et conservée l information génétique portée par les chromosomes.
Transformations nucléaires
 Transformations nucléaires Stabilité et instabilité des noyaux : Le noyau d un atome associé à un élément est représenté par le symbole A : nombre de masse = nombre de nucléons (protons + neutrons) Z :
Transformations nucléaires Stabilité et instabilité des noyaux : Le noyau d un atome associé à un élément est représenté par le symbole A : nombre de masse = nombre de nucléons (protons + neutrons) Z :
5.5.5 Exemple d un essai immunologique
 5.5.5 Exemple d un essai immunologique Test de grossesse Test en forme de bâtonnet destiné à mettre en évidence l'hormone spécifique de la grossesse, la gonadotrophine chorionique humaine (hcg), une glycoprotéine.
5.5.5 Exemple d un essai immunologique Test de grossesse Test en forme de bâtonnet destiné à mettre en évidence l'hormone spécifique de la grossesse, la gonadotrophine chorionique humaine (hcg), une glycoprotéine.
Conférence technique internationale de la FAO
 Décembre 2009 ABDC-10/7.2 F Conférence technique internationale de la FAO Biotechnologies agricoles dans les pays en développement: choix et perspectives pour les cultures, les forêts, l élevage, les pêches
Décembre 2009 ABDC-10/7.2 F Conférence technique internationale de la FAO Biotechnologies agricoles dans les pays en développement: choix et perspectives pour les cultures, les forêts, l élevage, les pêches
AGREGATION DE BIOCHIMIE GENIE BIOLOGIQUE
 AGREGATION DE BIOCHIMIE GENIE BIOLOGIQUE CONCOURS EXTERNE Session 2005 TRAVAUX PRATIQUES DE BIOCHIMIE PHYSIOLOGIE ALCOOL ET FOIE L éthanol, psychotrope puissant, est absorbé passivement dans l intestin
AGREGATION DE BIOCHIMIE GENIE BIOLOGIQUE CONCOURS EXTERNE Session 2005 TRAVAUX PRATIQUES DE BIOCHIMIE PHYSIOLOGIE ALCOOL ET FOIE L éthanol, psychotrope puissant, est absorbé passivement dans l intestin
Transport des gaz dans le sang
 UE3-2 - Physiologie Physiologie Respiratoire Chapitre 9 : Transport des gaz dans le sang Docteur Sandrine LAUNOIS-ROLLINAT Année universitaire 2011/2012 Université Joseph Fourier de Grenoble - Tous droits
UE3-2 - Physiologie Physiologie Respiratoire Chapitre 9 : Transport des gaz dans le sang Docteur Sandrine LAUNOIS-ROLLINAT Année universitaire 2011/2012 Université Joseph Fourier de Grenoble - Tous droits
Chapitre 2 - Complexité des relations entre génotype et phénotype
 Chapitre 2 - Complexité des relations entre génotype et phénotype Chaque chromosome est en double exemplaire Donc chaque gène (situé sur son locus) est en double exemplaires : et peut être sous différente
Chapitre 2 - Complexité des relations entre génotype et phénotype Chaque chromosome est en double exemplaire Donc chaque gène (situé sur son locus) est en double exemplaires : et peut être sous différente
Séquence 4. La nature du vivant. Sommaire. 1. L unité structurale et chimique du vivant. 2. L ADN, support de l information génétique
 Séquence 4 La nature du vivant Sommaire 1. L unité structurale et chimique du vivant 2. L ADN, support de l information génétique 3. Synthèse de la séquence 4. Exercices Devoir autocorrectif n 2 Séquence
Séquence 4 La nature du vivant Sommaire 1. L unité structurale et chimique du vivant 2. L ADN, support de l information génétique 3. Synthèse de la séquence 4. Exercices Devoir autocorrectif n 2 Séquence
1.2 Coordinence. Notion de liaison de coordinence : Cas de NH 3. et NH 4+ , 3 liaisons covalentes + 1 liaison de coordinence.
 Règle de l octet : tendance qu on les atomes à s entourer de 8 électrons dans l édifice moléculaire. Ce n est pas une règle générale. Composés respectant la règle de l octet Composés ne respectant pas
Règle de l octet : tendance qu on les atomes à s entourer de 8 électrons dans l édifice moléculaire. Ce n est pas une règle générale. Composés respectant la règle de l octet Composés ne respectant pas
Energie Nucléaire. Principes, Applications & Enjeux. 6 ème - 2014/2015
 Energie Nucléaire Principes, Applications & Enjeux 6 ème - 2014/2015 Quelques constats Le belge consomme 3 fois plus d énergie que le terrien moyen; (0,56% de la consommation mondiale pour 0,17% de la
Energie Nucléaire Principes, Applications & Enjeux 6 ème - 2014/2015 Quelques constats Le belge consomme 3 fois plus d énergie que le terrien moyen; (0,56% de la consommation mondiale pour 0,17% de la
CATALOGUE DES PRESTATIONS DE LA
 1/23 La plate-forme Biopuces et Séquençage de Strasbourg est équipée des technologies Affymetrix et Agilent pour l étude du transcriptome et du génome sur puces à ADN. SOMMAIRE ANALYSE TRANSCRIPTIONNELLE...
1/23 La plate-forme Biopuces et Séquençage de Strasbourg est équipée des technologies Affymetrix et Agilent pour l étude du transcriptome et du génome sur puces à ADN. SOMMAIRE ANALYSE TRANSCRIPTIONNELLE...
Chapitre 6. Réactions nucléaires. 6.1 Généralités. 6.1.1 Définitions. 6.1.2 Lois de conservation
 Chapitre 6 Réactions nucléaires 6.1 Généralités 6.1.1 Définitions Un atome est constitué d électrons et d un noyau, lui-même constitué de nucléons (protons et neutrons). Le nombre de masse, noté, est le
Chapitre 6 Réactions nucléaires 6.1 Généralités 6.1.1 Définitions Un atome est constitué d électrons et d un noyau, lui-même constitué de nucléons (protons et neutrons). Le nombre de masse, noté, est le
8/10/10. Les réactions nucléaires
 Les réactions nucléaires En 1900, à Montréal, Rutherford observa un effet curieux, lors de mesures de l'intensité du rayonnement d'une source de thorium [...]. L'intensité n'était pas la même selon que
Les réactions nucléaires En 1900, à Montréal, Rutherford observa un effet curieux, lors de mesures de l'intensité du rayonnement d'une source de thorium [...]. L'intensité n'était pas la même selon que
CHAPITRE 2 : Structure électronique des molécules
 CHAPITRE 2 : Structure électronique des molécules I. La liaison covalente 1) Formation d une liaison covalente Les molécules sont des assemblages d atomes liés par des liaisons chimiques résultant d interactions
CHAPITRE 2 : Structure électronique des molécules I. La liaison covalente 1) Formation d une liaison covalente Les molécules sont des assemblages d atomes liés par des liaisons chimiques résultant d interactions
Génomique Comparative et intégrative
 Génomique Comparative et intégrative Introduction : Le big data : on peut traiter des données massives à présent, l'objectif à présent est d'éviter les transferts de données trop longs. On a tout à portée
Génomique Comparative et intégrative Introduction : Le big data : on peut traiter des données massives à présent, l'objectif à présent est d'éviter les transferts de données trop longs. On a tout à portée
ANALYSE SPECTRALE. monochromateur
 ht ANALYSE SPECTRALE Une espèce chimique est susceptible d interagir avec un rayonnement électromagnétique. L étude de l intensité du rayonnement (absorbé ou réémis) en fonction des longueurs d ode s appelle
ht ANALYSE SPECTRALE Une espèce chimique est susceptible d interagir avec un rayonnement électromagnétique. L étude de l intensité du rayonnement (absorbé ou réémis) en fonction des longueurs d ode s appelle
Vieillissement moléculaire et cellulaire
 Vieillissement moléculaire et cellulaire Yves Courtois Lorsque, en 1961, Léonard HAYFLICK, aux USA, re - mit en cause le concept général de l'immortalité des cellules, il apportait à la recherche en gérontologie
Vieillissement moléculaire et cellulaire Yves Courtois Lorsque, en 1961, Léonard HAYFLICK, aux USA, re - mit en cause le concept général de l'immortalité des cellules, il apportait à la recherche en gérontologie
MYRIAD. l ADN isolé n est à présent plus brevetable!
 MYRIAD La Cour Suprême des Etats-Unis revient sur plus de 30 ans de pratique : l ADN isolé n est à présent plus brevetable! Mauvaise passe pour les inventions en biotechnologies sur le territoire américain.
MYRIAD La Cour Suprême des Etats-Unis revient sur plus de 30 ans de pratique : l ADN isolé n est à présent plus brevetable! Mauvaise passe pour les inventions en biotechnologies sur le territoire américain.
grande simple microscope microscope inventé années 1825. biologie = cellule) et (logos de plus en Anglais. Utilise un La microscopie, 1665,
 et (logos de plus en Anglais. Utilise un La microscopie, 1665,") Cours de Biologie Cellulaire Présentés par Mr CHELLI A. FSNV 2012/ /2013 CHAPITRE I : INTRODUCTION A LA BIOLOGIE CELLULAIRE A- Introduction et définitionn de la biologie cellulaire : Il était difficile
Cours de Biologie Cellulaire Présentés par Mr CHELLI A. FSNV 2012/ /2013 CHAPITRE I : INTRODUCTION A LA BIOLOGIE CELLULAIRE A- Introduction et définitionn de la biologie cellulaire : Il était difficile
Atelier : L énergie nucléaire en Astrophysique
 Atelier : L énergie nucléaire en Astrophysique Elisabeth Vangioni Institut d Astrophysique de Paris Fleurance, 8 Août 2005 Une calculatrice, une règle et du papier quadrillé sont nécessaires au bon fonctionnement
Atelier : L énergie nucléaire en Astrophysique Elisabeth Vangioni Institut d Astrophysique de Paris Fleurance, 8 Août 2005 Une calculatrice, une règle et du papier quadrillé sont nécessaires au bon fonctionnement
Détection des duplications en tandem au niveau nucléique à l'aide de la théorie des flots
 Université Toulouse 3 Paul Sabatier(UT3 Paul Sabatier) Informatique Spécialité Bioinformatique Eric AUDEMARD lundi 28 novembre 2011 Détection des duplications en tandem au niveau nucléique à l'aide de
Université Toulouse 3 Paul Sabatier(UT3 Paul Sabatier) Informatique Spécialité Bioinformatique Eric AUDEMARD lundi 28 novembre 2011 Détection des duplications en tandem au niveau nucléique à l'aide de
ÉCOLES NORMALES SUPÉRIEURES ÉCOLE NATIONALE DES PONTS ET CHAUSSÉES CONCOURS D ADMISSION SESSION 2013 FILIÈRE BCPST COMPOSITION DE BIOLOGIE
 ÉCOLES NORMALES SUPÉRIEURES ÉCOLE NATIONALE DES PONTS ET CHAUSSÉES CONCOURS D ADMISSION SESSION 2013 FILIÈRE BCPST COMPOSITION DE BIOLOGIE Épreuve commune aux ENS de Cachan, Lyon, Paris et de l ENPC Durée
ÉCOLES NORMALES SUPÉRIEURES ÉCOLE NATIONALE DES PONTS ET CHAUSSÉES CONCOURS D ADMISSION SESSION 2013 FILIÈRE BCPST COMPOSITION DE BIOLOGIE Épreuve commune aux ENS de Cachan, Lyon, Paris et de l ENPC Durée
Leslie REGAD ; Gaëlle LELANDAIS. leslie.regad@univ- paris- diderot.fr ; gaelle.lelandais@univ- paris- diderot.fr
 Programmation avec R Polycopié d exercices Leslie REGAD ; Gaëlle LELANDAIS leslie.regad@univ- paris- diderot.fr ; gaelle.lelandais@univ- paris- diderot.fr Ce polycopié rassemble différents exercices vous
Programmation avec R Polycopié d exercices Leslie REGAD ; Gaëlle LELANDAIS leslie.regad@univ- paris- diderot.fr ; gaelle.lelandais@univ- paris- diderot.fr Ce polycopié rassemble différents exercices vous
ARRÊTÉ du. Projet d arrêté fixant le programme d'enseignement de santé et social en classe de seconde générale et technologique
 RÉPUBLIQUE FRANÇAISE Ministère de l Éducation nationale NOR : MENE1007267A ARRÊTÉ du Projet d arrêté fixant le programme d'enseignement de santé et social en classe de générale et technologique Le Ministre
RÉPUBLIQUE FRANÇAISE Ministère de l Éducation nationale NOR : MENE1007267A ARRÊTÉ du Projet d arrêté fixant le programme d'enseignement de santé et social en classe de générale et technologique Le Ministre
Résonance Magnétique Nucléaire : RMN
 21 Résonance Magnétique Nucléaire : RMN Salle de TP de Génie Analytique Ce document résume les principaux aspects de la RMN nécessaires à la réalisation des TP de Génie Analytique de 2ème année d IUT de
21 Résonance Magnétique Nucléaire : RMN Salle de TP de Génie Analytique Ce document résume les principaux aspects de la RMN nécessaires à la réalisation des TP de Génie Analytique de 2ème année d IUT de
TRAITEMENTS ANTI-VIRAUX
 UE de l Agent Infectieux à l Hôte Mars 2012 TRAITEMENTS ANTI-VIRAUX Dr Isabelle GARRIGUE UMR CNRS MFP Microbiologie Fondamentale et Pathogénicité isabelle.garrigue@chu-bordeaux.fr La lutte antivirale Mesures
UE de l Agent Infectieux à l Hôte Mars 2012 TRAITEMENTS ANTI-VIRAUX Dr Isabelle GARRIGUE UMR CNRS MFP Microbiologie Fondamentale et Pathogénicité isabelle.garrigue@chu-bordeaux.fr La lutte antivirale Mesures
PARTIE I Compte pour 75 %
 PARTIE I Compte pour 75 % Instructions : Noircissez la lettre correspondant à la bonne réponse sur la feuille de réponse fournie. 1. Dans le diagramme, quelles structures font partie du système nerveux
PARTIE I Compte pour 75 % Instructions : Noircissez la lettre correspondant à la bonne réponse sur la feuille de réponse fournie. 1. Dans le diagramme, quelles structures font partie du système nerveux
Les cytokines et leurs récepteurs. Laurence Guglielmi laurence.guglielmi@univ-montp1.frli
 Les cytokines et leurs récepteurs Laurence Guglielmi laurence.guglielmi@univ-montp1.frli l i@ i 1 Les cytokines et leurs récepteurs 2 mécanismes principaux d interactions cellulaires : - contact membranaire
Les cytokines et leurs récepteurs Laurence Guglielmi laurence.guglielmi@univ-montp1.frli l i@ i 1 Les cytokines et leurs récepteurs 2 mécanismes principaux d interactions cellulaires : - contact membranaire
Mise en place d une solution automatique de stockage et de visualisation de données de capture des interactions chromatiniennes à l échelle génomique
 Rapport de stage de deuxième année de DUT Génie Biologique option Bioinformatique Mise en place d une solution automatique de stockage et de visualisation de données de capture des interactions chromatiniennes
Rapport de stage de deuxième année de DUT Génie Biologique option Bioinformatique Mise en place d une solution automatique de stockage et de visualisation de données de capture des interactions chromatiniennes
Recommandations pour la surveillance de la température de l eau chaude sanitaire
 Délégation territoriale du Val-d Oise Recommandations pour la surveillance de la température de l eau chaude sanitaire Maîtriser la température de l eau chaude sanitaire L exploitant d un réseau d eau
Délégation territoriale du Val-d Oise Recommandations pour la surveillance de la température de l eau chaude sanitaire Maîtriser la température de l eau chaude sanitaire L exploitant d un réseau d eau
TP N 3 La composition chimique du vivant
 Thème 1 : La Terre dans l'univers, la vie et l'évolution du vivant : une planète habitée Chapitre II : La nature du vivant TP N 3 La composition chimique du vivant Les conditions qui règnent sur terre
Thème 1 : La Terre dans l'univers, la vie et l'évolution du vivant : une planète habitée Chapitre II : La nature du vivant TP N 3 La composition chimique du vivant Les conditions qui règnent sur terre
UE6 - Cycle de vie du médicament : Conception rationnelle
 UE6 - Cycle de vie du médicament : Conception rationnelle Dr. Raphaël Terreux Faculté de Pharmacie (ISPB) Département pédagogique des Sciences Physico-Chimiques et Pharmacie Galénique 8 avenue Rockefeller,
UE6 - Cycle de vie du médicament : Conception rationnelle Dr. Raphaël Terreux Faculté de Pharmacie (ISPB) Département pédagogique des Sciences Physico-Chimiques et Pharmacie Galénique 8 avenue Rockefeller,
Physique Chimie. Utiliser les langages scientifiques à l écrit et à l oral pour interpréter les formules chimiques
 C est Niveau la représentation 4 ème 2. Document du professeur 1/6 Physique Chimie LES ATOMES POUR COMPRENDRE LA TRANSFORMATION CHIMIQUE Programme Cette séance expérimentale illustre la partie de programme
C est Niveau la représentation 4 ème 2. Document du professeur 1/6 Physique Chimie LES ATOMES POUR COMPRENDRE LA TRANSFORMATION CHIMIQUE Programme Cette séance expérimentale illustre la partie de programme
------- SESSION 2013 ÉPREUVE À OPTION. (durée : 4 heures coefficient : 6 note éliminatoire 4 sur 20) CHIMIE
 CHIMIE") CNCURS SUR ÉPREUVES UVERT AUX CANDIDATS TITULAIRES D UN DIPLÔME U TITRE CNFÉRANT LE GRADE DE MASTER U D'UN DIPLÔME U TITRE HMLGUÉ U ENREGISTRÉ AU RÉPERTIRE NATINAL DES CERTIFICATINS PRFESSINNELLES AU NIVEAU
CNCURS SUR ÉPREUVES UVERT AUX CANDIDATS TITULAIRES D UN DIPLÔME U TITRE CNFÉRANT LE GRADE DE MASTER U D'UN DIPLÔME U TITRE HMLGUÉ U ENREGISTRÉ AU RÉPERTIRE NATINAL DES CERTIFICATINS PRFESSINNELLES AU NIVEAU
MABioVis. Bio-informatique et la
 MABioVis Modèles et Algorithmes pour la Bio-informatique et la Visualisation Visite ENS Cachan 5 janvier 2011 MABioVis G GUY MELANÇON (PR UFR Maths Info / EPI GRAVITE) (là, maintenant) - MABioVis DAVID
MABioVis Modèles et Algorithmes pour la Bio-informatique et la Visualisation Visite ENS Cachan 5 janvier 2011 MABioVis G GUY MELANÇON (PR UFR Maths Info / EPI GRAVITE) (là, maintenant) - MABioVis DAVID