Dépôt Institutionnel de l Université libre de Bruxelles / Université libre de Bruxelles Institutional Repository Thèse de doctorat/ PhD Thesis
|
|
|
- Jean-Paul Beausoleil
- il y a 3 ans
- Total affichages :
Transcription
1 Dépôt Institutionnel de l Université libre de Bruxelles / Université libre de Bruxelles Institutional Repository Thèse de doctorat/ PhD Thesis Citation APA: Schiffmann, S. N. (1992). Implications de l'adénosine, la cholecystokinine et leurs récepteurs dans l'organisation fonctionnelle des ganglions de la base (Unpublished doctoral dissertation). Université libre de Bruxelles, Faculté de Médecine Médecine, Bruxelles. Disponible à / Available at permalink : (English version below) Cette thèse de doctorat a été numérisée par l Université libre de Bruxelles. L auteur qui s opposerait à sa mise en ligne dans DI-fusion est invité à prendre contact avec l Université (di-fusion@ulb.ac.be). Dans le cas où une version électronique native de la thèse existe, l Université ne peut garantir que la présente version numérisée soit identique à la version électronique native, ni qu elle soit la version officielle définitive de la thèse. DI-fusion, le Dépôt Institutionnel de l Université libre de Bruxelles, recueille la production scientifique de l Université, mise à disposition en libre accès autant que possible. Les œuvres accessibles dans DI-fusion sont protégées par la législation belge relative aux droits d'auteur et aux droits voisins. Toute personne peut, sans avoir à demander l autorisation de l auteur ou de l ayant-droit, à des fins d usage privé ou à des fins d illustration de l enseignement ou de recherche scientifique, dans la mesure justifiée par le but non lucratif poursuivi, lire, télécharger ou reproduire sur papier ou sur tout autre support, les articles ou des fragments d autres œuvres, disponibles dans DI-fusion, pour autant que : - Le nom des auteurs, le titre et la référence bibliographique complète soient cités; - L identifiant unique attribué aux métadonnées dans DI-fusion (permalink) soit indiqué; - Le contenu ne soit pas modifié. L œuvre ne peut être stockée dans une autre base de données dans le but d y donner accès ; l identifiant unique (permalink) indiqué ci-dessus doit toujours être utilisé pour donner accès à l œuvre. Toute autre utilisation non mentionnée ci-dessus nécessite l autorisation de l auteur de l œuvre ou de l ayant droit English Version This Ph.D. thesis has been digitized by Université libre de Bruxelles. The author who would disagree on its online availability in DI-fusion is invited to contact the University (di-fusion@ulb.ac.be). If a native electronic version of the thesis exists, the University can guarantee neither that the present digitized version is identical to the native electronic version, nor that it is the definitive official version of the thesis. DI-fusion is the Institutional Repository of Université libre de Bruxelles; it collects the research output of the University, available on open access as much as possible. The works included in DI-fusion are protected by the Belgian legislation relating to authors rights and neighbouring rights. Any user may, without prior permission from the authors or copyright owners, for private usage or for educational or scientific research purposes, to the extent justified by the non-profit activity, read, download or reproduce on paper or on any other media, the articles or fragments of other works, available in DI-fusion, provided: - The authors, title and full bibliographic details are credited in any copy; - The unique identifier (permalink) for the original metadata page in DI-fusion is indicated; - The content is not changed in any way. It is not permitted to store the work in another database in order to provide access to it; the unique identifier (permalink) indicated above must always be used to provide access to the work. Any other use not mentioned above requires the authors or copyright owners permission.

2 Université Libre de Bruxelles Faculté de Médecine Laboratoire de Neuropathologie et de Recherche sur les Peptides du Système Nerveux Prof. J.-J. Vanderhaeghen IMPLICATIONS PE L ADENOSINE, LA CHOLECYSTOKININE ET LEURS DANS L ORGANISATION FONCTIONNELLE. =. 5 ; ç.... DES GANGLIONS DE LA BASE Serge SCHIFFMANN Volume 1 Mémoire présenté en vue de l obtention du titre d Agrégé de l Enseignement Supérieur (1992)
3 % f UNIVERSITE LIBRE DE BRUXELLES FACULTE DE MEDECINE Epreuve publique pour l'obtention du titre légal d'agrégé de l'enseignement supérieur M. Serge SCHIFFMANN Docteur en Médecine, Chirurgie et Ac «aents défendra publiquement le mercredi 29 avril 1992 à 18 heures, une thèse d'agrégation intitulée : "IMPLICATIONS DE L'ADENOSINE, LA CHOLECYSTOKININE ET LEURS RECEPTEURS DANS L'ORGANISATION FONCTIONNELLE DES GANGLIONS DE LA BASE" M. SCHIFFMANN fera une leçon sur le sujet suivant : "PHYSIOPATHOLOGIE DES MOUVEMENTS ANORMAUX" Auditoire Bordet (local F2.304) route de Lennik Bruxelles
4 I «r 1 Université Libre de Bruxelles - Campus ERASME-Anderlecht Laboratoire de Neuropathologie et de Recherche sur les Peptides du Système Nerveux Laboratoire d Anatomie Pathologique et de Microscopie Electronique Directeur: Professeur Jean-Jacques Vanderhaeghen Monsieur le Professeur J-L Vanherweghem, Doyen de la Faculté de Médecine Bruxelles, le 11 Mars 1992 Monsieur le Doyen, Je vous prie de trouver ci-dessous mon choix concernant la consultation de ma thèse déposée en vue de l obtention du titre d Agrégé de l enseignement supérieur. 3.a) Consultation Autorisée Je reste à votre disposition pour toute information complémentaire et vous prie de croire. Monsieur le Doyen, à mes sentiments dévoués et respectueux. Serge N. SCHIFFMANN 808 Route de Lennik C.P Bruxelles - Tél: 02/ Fax: 02/
5 Université Libre de Bruxelles Faculté de Médecine Laboratoire de Neuropathologie et de Recherche sur les Peptides du Système Nerveux Prof. J.-J. Vanderhaeghen IMPLICATIONS DE L ADENOSINE, LA CHOLECYSTOKININE ET LEURS RECEPTEURS DANS L ORGANISATION FONCTIONNELLE^^^^^^^^^^^^^^^^^ DES GANGLIONS DE LA BASE Serge SCHIFFMANN Volume 1 Mémoire présenté en vue de l obtention du titre d Agrégé de l Enseignement Supérieur (1992)
6 A Michèle A ma Famille A Boris
7 Remerciements Je tiens à exprimer ma profonde reconnaissance au professeur Jean-Jacques Vanderhaeghen pour m avoir accueilli dans son laboratoire, pour avoir su me transmettre son enthousiasme sans faille pour la recherche et pour avoir guidé mes travaux dans les méandres des connexions des "basal ganglia". Je remercierai également tous les chercheurs passés ou présents du laboratoire qui par leurs avis et encouragements ont contribué à la progression harmonieuse de ma formation et à l aboutissement de ce mémoire. Je citerai tout d abord Serge Goldman qui a guidé mes premiers pas au laboratoire après avoir attisé mon intérêt en neurologie clinique, et ensuite Serge Przedborski, Pierre Mailleux, Françoise Lotstra, Erik Teugels, Olivier Van Reeth, Jean-Marie Vanderwinden, Rafik Karmali, Eric Wespes et Olivier Jacobs. Mon ouverture au monde de la biologie moléculaire n aurait été possible sans les avis et conseils du professeur Gilbert Vassart et de Frédérik Libert et Marc Parmentier de ri.r.i.b.h.n. Qu ils en soient ici cordialement remercier. La confiance que m ont accordé les professeurs Gilbert Vassart et Jacques Dumont tout au long de la collaboration de nos deux laboratoires suscite aussi ma reconnaissance. Mes remerciements vont également à Martin Schalling et au professeur Tomas Hôkfelt du laboratoire d Histologie et de Neurobiologie du Karolinska Institut de Stockholm, pour m avoir initier aux techniques d hybridation in situ et pour leur accueil lors de mon séjour dans leur laboratoire. Les longues discussions et les longues marches à la Nouvelle Orléans sont autant de moments dont je voudrais remercier Gabriel Von Euler du Karolinska Institut de Stockholm. Je tiens enfin à remercier chaleureusement Pascale Halleux et Roberte Menu pour la qualité de leur aide technique, Jean-Louis Conreur pour celle, incomparable, de son travail iconographique et Michèle De Bardemaeker pour son aide dans la dactylographie de ce mémoire ainsi que tous les autres membres du laboratoire dont la sympathie et l aide occasionnelle m ont toujours été d un grand support.
8 TABLE DES MATIERES 1. I. INTRODUCTION GENERALE 7. IL ORGANISATION STRUCTURELLE, CHIMIQUE ET FONCTIONNELLE DES GANGLIONS DE LA BASE. Rappels bibliographiques. 8. II. 1. Définition 8. II.2 Neuroanatomie 8. II.2.a Le circuit principal 9. Noyau afférent: le striatum 11. Noyaux efférents: le globus pallidus interne et la substance noire pars reticulata 12. II.2.b Extensions ventrales des noyaux du circuit principal 14. II.2.C Organisation en boucles parallèles 16. II.2.d Relais intrinsèques et voies modulatrices: la voie nigro-striatale et la boucle pallido-sous-thalamo-pallidale 16. La voie nigro-striatale 16. La boucle pallido-sous-thalamo-pallidale 16. II.3 Histologie 18. II.4 Neurochimie 19. II.4.a Le striatum 21. II.4.b La voie cortico-striatale 21. II.4.C La voie nigro-striatale (substance noire pars compacta) 22. II.4.d Le globus pallidus et la substance noire pars reticulata 22. II.4.e Le noyau sous-thalamique 22. II.5 Compartimentalisation du striatum 25. II.6 Rôle des ganglions de la base dans la physiologie du mouvement et du comportement. Applications en pathologie. 28. III. PREMIERE PARTIE: LOCALISATION ET FONCTION DES RECEPTEURS DE L ADENOSINE. IDENTIFICATION DES GENES ENCODANT LES RECEPTEURS Al ET A2 30. III. 1 Introduction et Objectifs 33. 1II.2 Résultats et Discussion a Identification et caractérisation de RDC7 et RDC8 comme récepteurs Al et A2 de l adénosine (articles 1, 2, 3) b Localisation du récepteur A2a de l adénosine dans les ganglions de la base et identification phénotypique des neurones qui l expriment (articles 1, 4, 5, 6) C Rôle(s) du(es) récepteur(s) A2 dans la régulation de l expression génétique dans le striatum (article 6) d Modification d expression du récepteur A2a de l adénosine dans le striatum au cours du développement (article 7) III.3 Conclusions et Perspectives
9 46. IV. DEUXIEME PARTIE; LOCALISATION ET FONCTION DE LA CHOLECYSTOKININE ET SES RECEPTEURS 48. IV. 1 Introduction et Objectifs 51. IV.2 Résultats et Discussion 51. IV.2.a Localisation immunohistochimique de la cholécystokinine dans les ganglions de la base chez l homme (article 8) 53. IV.2.b Localisation de TARN messager encodant la cholécystokinine dans les ganglions de la base et leurs structures afférentes avec référence particulière au mésencéphale ventral (articles 9, 10, 11) 56. IV.2.C Régulation dopaminergique de l expression de l ARN messager encodant la cholécystokinine dans le striatum (article 12) 57. IV.2.d Ontogenèse des récepteurs de la cholécystokinine dans le striatum humain (article 13) 58. IV.3 Conclusions et perspectives 60. V. BIBLIOGRAPHIE 74. VI. THESES ANNEXES
10 -1- I. INTRODUCTION GENERALE
11 -2- Le système des ganglions de la base est impliqué dans la pathogénie de multiples affections neurologiques voire également psychiatriques. L association de divers tableaux cliniques caractérisés par des anomalies des mouvements avec des lésions dégénératives au sein de ce système est connue depuis le début du siècle. L atrophie des noyaux caudé et putamen, caractéristique de la maladie de Huntington, est en effet décrite dans les années 1920 (Lewy, 1921; Dunlap, 1927). Longtemps, les connaissances dans ce domaine ont progressé par les corrélations entre les lésions anatomo-pathologiques et la sémiologie neurologique. La découverte par Horniciewicz (1963), d une déplétion en dopamine dans les ganglions de la base de patients ayant souffert de la maladie de Parkinson ouvre la voie à une approche différente de la physiopathologie des affections de ces noyaux. Cette approche neurochimique a été accélérée par la découverte dans le système nerveux, aux côtés des neurotransmetteurs "classiques" comme l acide gamma-aminobutyrique (GABA), le glutamate, la dopamine et l acétylcholine, d un grand nombre de molécules y compris peptidiques (Vanderhaeghen et al., 1975) auxquelles on a attribué des fonctions neuromédiatrices. Le développement de techniques neurobiochimiques performantes ainsi que les apports récents de la biologie moléculaire ont permis de démontrer l existence de mécanismes d adaptation ou de plasticité fonctionnelle induits par des déafférentiations expérimentales lésionnelles ou pharmacologiques. Dans l étude de ce concept, le meilleur modèle expérimental est précisément celui qui mime la maladie de Parkinson, à savoir la destruction sélective de la voie dopaminergique nigro-striée (pour revue voir Zigmond, 1990). De nombreuses études ont en effet montré qu outre les modifications de synthèse et de libération de dopamine dans les neurones dopaminergiques épargnés par la lésion, cette déafférentiation induit des modifications d expression de neurotransmetteurs, de neuropeptides et de récepteurs dans les neurones striataux post-synaptiques (pour revue voir Graybiel, 1990). Le rôle déterminant de la voie dopaminergique nigro-striée dans le contrôle du système des ganglions de la base est confirmé par l utilisation prépondérante de drogues agissant au niveau de la synapse dopaminergique dans le traitement des affections neuro-psychiatriques de ce système.
12 -3- L adénosine et la cholécystokinine présentent la caractéristique, comme la dopamine, d être des neuromédiateurs abondants dans le système des ganglions de la base, et d être directement impliquées dans le contrôle de l activité motrice (Green et al., 1982, Zetler, 1985; Durcan and Morgan, 1989; Ferre et al., 1991). De plus, par rapport à d autres éléments neurobiochimiques présents dans ce système, l adénosine et la cholécystokinine ont été relativement moins étudiées. Notre objectif dans ce travail est de préciser les relations anatomiques et fonctionnelles de ces deux neuromédiateurs avec le système dopaminergique dans les ganglions de la base. Sur base de ces observations, nous avons posé des hypothèses quant à leur intervention dans la pathogénie de certaines affections des noyaux de la base et proposé de nouvelles solutions thérapeutiques. L adénosine est une molécule ubiquitaire participant au métabolisme cellulaire. La localisation ubiquitaire de l adénosine nécessite la caractérisation anatomique et pharmacologique de ses récepteurs afin de comprendre les fonctions neuromodulatrices de cette molécule. Ces récepteurs sont par ailleurs la cible de molécules psychotropes de consommation courante telle que la caféine. Notre premier objectif a été d identifier et de caractériser deux récepteurs clonés à ri.r.i.b.h.n. mais non identifiés et donc appelés récepteurs "orphelins" (Libert et al., 1989). Ce travail a mené à leur identification comme récepteurs Al et A2 de l adénosine. Ce résultat ouvre la voie d une nouvelle pharmacologie des récepteurs de l adénosine car tant au niveau du système nerveux central qu au niveau périphérique (cardiaque, pulmonaire,...) le développement d agonistes et d antagonistes Al et A2 sélectifs s avère indispensable pour envisager leur utilisation clinique. Cette caractérisation a permis ensuite de préciser les relations neuroanatomo-chimiques du récepteur A2a de l adénosine avec la dopamine et ses récepteurs dans les ganglions de la base. Elle a permis également d étudier la régulation de l expression génétique de divers neuromédiateurs dans le striatum par les récepteurs A2 de l adénosine et les interactions de cette régulation avec le contrôle dopaminergique.
13 -4- Le récepteur A2a de l adénosine est exprimé sélectivement par une sous-population striatale à l origine d une voie de régulation qui est hyperactive dans les syndromes hypokinétiques et hypoactive dans les pathologies hyperkinétiques. Par l activation de ce récepteur, l adénosine exerce un contrôle positif sur l expression génétique de ces neurones. Ces résultats permettent d envisager le développement de nouveaux antagonistes sélectifs du récepteur A2a comme compléments thérapeutiques dans les affections caractérisées par le parkinsonisme. Notre second objectif est de préciser la distribution de la cholécystokinine et ses relations avec le système dopaminergique dans les noyaux de la base chez l animal et l homme ainsi que d investiguer l éventualité d une expression de cholécystokinine par les neurones striataux après modifications expérimentales de la neurotransmission dopaminergique. Chez l homme, l abondance des fibres à cholécystokinine dans le striatum ventral et les autres régions des ganglions de la base en relation avec le système limbique évoque sa participation aux mécanismes d initiation des mouvements en réponse aux stimuli émotionnels. L expression de cholécystokinine dans les voies cortico-, thalamo-, et nigro-striatales et dans d autres structures cérébrales projetant vers le striatum indique que ce peptide peut avoir de multiples fonctions tant dans les systèmes moteur que cognitif et limbique. De plus, une déplétion expérimentale en dopamine induit l expression de cholécystokinine par certains neurones striataux, ce qui constitue un nouvel exemple de plasticité fonctionnelle. Enfin, l expression plus importante de cholécystokinine par les neurones dopaminergiques des voies nigro-striée et méso-limbique chez les patients souffrant de schizophrénie constitue un élément nouveau dans l étude neurochimique de cette affection qui malgré l intérêt qu il suscite, nécessite prudence et confirmation. Notre travail repose sur l utilisation des méthodes suivantes. L approche morphologique est abordée par les techniques immunohistochimiques et l hybridation in situ qui ont une résolution anatomique et cellulaire et par la méthode de radioliaison de ligands spécifiques
14 -5- sur coupes de tissus qui ne permet qu une résolution anatomique. Les caractérisations biochimique et pharmacologique sont abordées par les techniques de Northern blotting d ARN et celles de radioliaison sur préparations membranaires. Des méthodes de neurologie expérimentale ont permis de reproduire des modèles animaux qui ont été analysés à l aide des diverses méthodes reprises ci-dessus. Enfin, l existence de différences d espèces a justifié, autant que possible, leur application au matériel cérébral humain. Le détail de ces techniques est repris dans les différentes publications dont ce mémoire est la synthèse. Un rappel de l organisation structurelle, chimique et fonctionnelle des ganglions de la base sera suivi de la présentation et de la discussion de nos résultats dans deux chapitres successifs : Première partie: Localisation et fonction des récepteurs de l adénosine. Identification des gènes encodant les récepteurs Al et A2. Deuxième partie: Localisation et fonction de la cholécystokinine J

15 -6- Ce mémoire représente la synthèse des articles suivants: 1. SCHIFFMANN, S.N., LIBERT, F., VASSART, G., DUMONT, J.E. and VANDERHAEGHEN, J-J. : A cloned G protein-coupled protein with a distribution restricted to striatal medium-sized neurons. Possible relationship with DI dopamine receptor. Brain Res., 519, , MAENHAUT,C., VAN SANDE, J., LIBERT, F., ABRAMOVICZ, M., PARMENTIER, M., VANDERHAEGHEN, J.-J., DUMONT, J.E., VASSART, G. and SCHIFFMANN, S.N.: RDC8 codes for an A2 receptor with physiological constitutive activity. Biochem. Biophys. Res. Comm. 173(3), , LIBERT, F., SCHIFFMANN, S.N., LEFORT, A., PARMENTIER, M., GERARD, C., DUMONT, J.E., VANDERHAEGHEN, J.-J. and VASSART, G.: The orphan receptor cdna RDC7 encodes an Al adenosine receptor. EMBO J., 10: , (Note: F. Libert and S.N. Schiffmann contributed equally to this study.) 4. SCHIFFMANN, S.N., LIBERT, F., VASSART, G. and VANDERHAEGHEN, J-J. : Distribution of adenosine A2 receptor mrna in the human brain. Neurosci. Lett., 130: , SCHIFFMANN, S.N., JACOBS, O.P. and VANDERHAEGHEN, J.-J.: Striatal Restricted Adenosine A2 Receptor (RDC8) Is Expressed by Enkephalin but Not by Substance P Neurons: An In Situ Hybridization Histochemistry Study. J. Neurochem., 57: , SCHIFFMANN S.N. and VANDERHAEGHEN J.-J.: Adenosine A2 receptors regulate the gene expression of striatopallidal and striatonigral neurons. Submitted, SCHIFFMANN, S.N. and VANDERHAEGHEN J.-J.: Ontogeny of Gene Expression of Adenosine A2 Receptor in the Striatum: Early Localization in the Patch Compartment. J. Comp. Neurol., in press, SCHIFFMANN, S.N., MAILLEUX, P., PRZEDBORSKI, S., HALLEUX, P., LOTSTRA, F. and VANDERHAEGHEN, J-J.: Cholecystokinin distribution in the human striatum and related subcortical structures, Neurochem. Int., 14, , SCHIFFMANN, S.N., and VANDERHAEGHEN J.-J.: Distribution of cells containing mrna encoding cholecystokinin in the rat central nervous System. J. Comp. Neurol., 304, , SCHALLING, M., FRIBERG, K., BIRD, E., SCHIFFMANN, S., MAILLEUX, P., VANDERHAEGHEN, J-J. and HÔKFELT, T. : Presence of cholecystokinin mrna in dopamine cells in the ventral mesencephalon of a human with schizophrenia. Acta Physiol. Scand., 137, , SCHALLING, M., FRIBERG, K., SEROOGY, K., RIEDERER, P., BIRD, E., SCHIFFMANN, S.N., MAILLEUX, P., VANDERHAEGHEN, J.-J., KUGA, S., GOLDSTEIN, M., KITAHAMA, K., LUPPI, P.H., JOUVET, M. and HÔKFELT, T.: Analysis of expression of cholecystokinin in dopamine cells in the ventral mesencephalon of several species and in humans with schizophrenia. Proc. Ntl. Acad. Sel. USA, 87, , SCHIFFMANN S.N. and VANDERHAEGHEN J.-J.: Dopaminergic Régulation of cholecystokinin mrna expression in the rat striatum. An in situ hybridization histochemistry study. Submitted, SCHIFFMANN, S.N., GOLDMAN, S., HEYMAN, P., DE VUYST, E., DE ROY, G. and VANDERHAEGHEN J.-J.: Ontogeny of cholecystokinin receptors in the human striatum. Submitted, 1992.

16 -7- II. ORGANISATION STRUCTURELLE, CHIMIQUE ET FONCTIONNELLE DES GANGLIONS DE LA BASE. Rappels Bibliographiques Neuroplasticité fonctionnelle: Augmentation de l expression d enképhaline dans le striatum ipsilatéral à une lésion de la voie dopaminergique nigro-striée. Image en pseudo-couleur, le rouge correspondant aux régions les plus riches.
17 -8- Les ganglions de la base constituent un système cérébral majeur de contrôle de la motricité. Ils interviennent dans les processus de programmation et d exécution des ces mouvements par le traitement et le filtrage de diverses informations sensorielles ("sensory gating") (cf. paragraphe II.6.). Cette fonction s applique au système moteur mais également au contrôle de comportements plus complexes mettant en oeuvre les systèmes préfrontal cognitif et limbique émotionnel (cf. paragraphes II.2.C. et H.6.). Une lésion ou une perturbation pharmacologique située à un quelconque relais des connexions que forment ces noyaux provoquent des anomalies des mouvements. De longue date, l association de telles anomalies avec certaines dégénérescences sélectives de ces noyaux est connue en pathologie neurologique humaine. L implication de ce système dans la pathogénie de multiples affections neurologiques et psychiatriques motive dès lors les recherches tant cliniques que fondamentales dont il est l objet Définition La définition première des "ganglions de la base" est une définition anatomique. Elle englobe les différentes structures grises, situées à la base du télencéphale antérieur: les noyaux caudé, putamen et accumbens constituant le striatum et le globus pallidus. Cette définition anatomique est complétée par des données fonctionnelles qui justifient d y inclure le tubercule olfactif, le noyau sous-thalamique ou corps de Luys et la substance noire de Sômmering. Sur base anatomique, divers auteurs ont également considéré l amygdale et le noyau basal de Meynert comme parties intégrantes du système des ganglions de la base. Cependant, un consensus justifié par des données fonctionnelles existe actuellement à l encontre de cette proposition et exclut ces deux structures. Les rapports de l amygdale avec certains éléments des ganglions de la base sont néanmoins étroits et seront envisagés plus loin (cf. paragraphe II.2.b.). Par contre, la substance innominée, région dans laquelle l on trouve les neurones cholinergiques du noyau basal de Meynert, abrite aussi les extensions ventrales du striatum et du pallidum (cf. paragraphe II.2.b.). A ce titre, elle doit être partiellement prise en compte dans les définitions tant anatomique que fonctionnelle des ganglions de la base Neuroanatomie a. Le circuit principal Le schéma anatomo-fonctionnel central du système des ganglions de la base s intégre dans la boucle cortico-striato-pallido-thalamo-corticale (fig. II.l), dans laquelle le striatum constitue le premier relais et le pallidum la structure efférente majeure du système (fig. II.l). Sur cette boucle, se greffent deux voies modulatrices importantes: la voie dopaminergique nigro-striatale et la boucle pallido-sous-thalamo-pallidale (cf. paragraphe II.2.d.).
18 -9- Noyau afférent: le striatum Les noyaux caudé et putamen représentent les deux parties d une seule structure appelée le striatum, traversée par les contingents de fibres corticofugales, striatofugales et corticopétales constituées en capsule interne. L existence de ponts de substance grise traversant cette capsule interne et reliant de ce fait le caudé et le putamen confirme cette notion (fig. II.2). Par ailleurs, chez les mammifères inférieurs tels que les rongeurs, la capsule interne est faiblement formée et les fibres corticofugales, striatofugales et corticopétales traversent en fascicules un noyau unique, le caudé-putamen ou striatum, en lui donnant un aspect strié qui lui vaut son nom. Dans une moindre mesure, ces fascicules sont également présents dans le striatum humain (fig. II.2).
19 -10- % Fig. II.2: Striatum humain. Coupe transversale de striatum humain marquée histochimiquement pour la détection de l enzyme acétylcholine estérase. (ACC) noyau accumbens, (C) noyau caudé, (CI) capsule interne, (P) putamen. Le striatum reçoit son innervation majeure du cortex cérébral. L organisation de ces connexions cortico-striatales semble répondre au schéma selon lequel une région corticale projette vers la portion du striatum la plus proche (Kemp and Powell, 1970). Ce schéma doit être cependant complété par des données plus récentes telles que l existence de boucles parallèles et la compartimentalisation du striatum dont il sera question respectivement dans les paragraphes II.2.C. et II.5. L organisation de ces projections permet la conservation de l organisation topographique des cortex d origine. Ainsi, la somatotopie des cortex moteur primaire et somatosensoriel et la tonotopie du cortex auditif se retrouvent dans leurs régions striatales cibles respectives (Alexander et al., 1986; Reale and Imig, 1983). Cette organisation topographique sera préservée tout au long du circuit de connexions ainsi que dans ses voies modulatrices (fig. II.3).
 noyau caudé, (GPe) globus pallidus externe, (GPi) globus pallidus interne, (MC) cortex moteur, (Pu) putamen, (SC) cortex somatosensoriel, (SMA) aire motrice supplémentaire, (SN) substance noire,")
20 -11- Fig. II.3: Organisation somatotopique du circuit moteur contrôlant le membre supérieur. De DeLong and Alexander, (Cd) noyau caudé, (GPe) globus pallidus externe, (GPi) globus pallidus interne, (MC) cortex moteur, (Pu) putamen, (SC) cortex somatosensoriel, (SMA) aire motrice supplémentaire, (SN) substance noire, (STN) noyau sous-thalamique, (VLo) noyau ventro-latéral pars oralis du thalamus. Le striatum reçoit également des afférences thalamiques provenant principalement des noyaux intralaminaires et centraux du thalamus (Herkenham and Pert, 1981). Les neurones efférents du striatum projettent principalement vers les subdivisions interne et externe du globus pallidus et la pars reticulata de la substance noire (Chang et al., 1981; Beckstead and Cruz, 1986; Smith and Parent, 1986b). Les neurones projettant vers ces trois structures appartiennent majoritairement à des sous-populations différentes, y compris en ce qui concerne les neurones innervant le globus pallidus interne et la pars reticulata de la substance noire (Beckstead and Cruz, 1986). Noyaux efférents: le globus pallidus interne et la substance noire pars reticulata Ces deux structures sont les cibles majeures des projections striatales. Le globus pallidus forme le noyau lenticulaire avec le putamen caudal dont il est séparé par la lame médullaire externe (fig. II.4b). Néanmoins et malgré son utilisation fréquente en neuroanatomie, d un point de vue fonctionnel il est abusif de grouper ces deux structures sous un même terme générique. Le globus pallidus est divisé en une partie externe dont il sera question dans le paragraphe II.2.d. (globus pallidus chez les mammifères inférieurs) et une partie interne (noyau entopédonculaire chez les mammifères inférieurs) par la lame médullaire interne. Ces

21 -12- deux subdivisions ont des caractéristiques neurochimiques (cf. paragraphe II.4.) et des connexions différentes. La substance noire, située dans le mésencéphale ventral est constituée d une pars reticulata, ventrale, et d une pars compacta, dorsale. Cette dernière sera également envisagée dans le paragraphe II.2.d. Les caractéristiques morphologiques, physiologiques, neurochimiques et les connexions de la pars reticulata indiquent qu elle représente en fait une extension caudo-médiale du globus pallidus interne (DeLong and Georgopoulos, 1979; Nauta, 1979b). Ce dernier reçoit surtout ses afférences du putamen et la substance noire pars reticulata du caudé (Smith and Parent, 1986b). Les projections du globus pallidus interne et de la substance noire pars reticulata vers le thalamus constituent la voie effectrice principale des ganglions de la base et de fait la voie commune finale de l expression de l activité striatale (fig. II.l). Ils innervent les noyaux thalamiques ventro-latéral, ventral antérieur et médio-dorsal et dans une moindre mesure les noyaux intralaminaires (Nauta, 1979a; 1979b). Le globus pallidus interne et la substance noire pars reticulata innervent par ailleurs également des structures du tronc cérébral telles le noyau pédonculo-pontin et la formation réticulée pontine et enfin, la substance noire pars reticulata projette vers le colliculus supérieur (Nauta, 1979a; Beckstead, 1983). II.2.b. Extensions ventrales des noyaux du circuit principal Des données morphologiques, physiologiques, neurochimiques ainsi que des études des connexions ont permis de démontrer que les limites anatomiques des ganglions de la base s étendent ventralement jusqu à la surface inférieure du cerveau (Heimer et al., 1982; Heimer et al., 1987; Alheid and Heimer, 1988). Ils envahissent des structures telles que la substance innominée ou le tubercule olfactif qui n étaient pas classiquement reconnues comme parties du système. Les appellations de striatum ventral et de pallidum ventral ont donc été introduites. Ces structures ont été l objet d un grand intérêt en raison de leurs relations avec le système limbique. Le noyau accumbens est le constituant principal du striatum ventral et forme un continuum ventral et rostral avec les noyaux caudé et putamen (fig. II.4a). Outre les portions les plus ventrales du putamen caudal, le striatum ventral inclut également de larges territoires du tubercule olfactif. Celui-ci, plus aisément identifiable chez les rongeurs, est relié au noyau accumbens par des ponts de substance grise (fig. II.4c) confirmant leur unicité fonctionnelle.
 et caudal (b) marquées histochimiquement pour la détection de l acétylcholine estérase, (c) Coupe transversale de télencéphale de rat")
22 -13- TO Fig. II.4: Extensions ventrales du striatum et du pallidum. Coupes transversales au niveau du striatum humain rostral (a) et caudal (b) marquées histochimiquement pour la détection de l acétylcholine estérase, (c) Coupe transversale de télencéphale de rat comprenant le striatum rostral et marquée par hybridation in situ pour la détection de TARN messager encodant le précurseur de l enképhaline. (ACC) noyau accumbens, (AMY) amygdale, (C) noyau caudé, (GPe) globus pallidus externe, (GPi) globus pallidus interne, (P) putamen, (SI) substance innominée, (TO) tubercule olfactif Les neurones constitutifs de ces deux structures appartiennent aux mêmes classes que les neurones du striatum dorsal (cf. paragraphe II.3.) et possèdent également les mêmes caractéristiques neurochimiques (cf. paragraphe H.4.). Le striatum ventral reçoit lui aussi ses afférences du cortex cérébral mais à la différence du striatum dorsal il est innervé par les régions corticales appartenant au système limbique comme la formation hippocampique, le cortex entorhinal, le cortex olfactif, le cortex cingulaire antérieur et le noyau basolatéral amygdalien (Groenewegen et al., 1987). Le striatum ventral projette vers les extensions pallidales ventrales ou pallidum ventral (Groenewegen and Russchen, 1984). Ce dernier est situé sous la commissure blanche antérieure, il peut s étendre jusqu au tubercule olfactif qu il envahit et l espace perforé antérieur où ses constituants s entremêlent avec des éléments du striatum ventral (Heimer et al., 1982; Haber and Nauta, 1983; Alheid and Heimer, 1988). Plus caudalement, il occupe une partie importante de la substance innominée (fig. II.4b). La neurochimie du pallidum ventral (cf. paragraphe II.4.) indique que
23 -14- les deux subdivisions du globus pallidus contribuent à l extension pallidale ventrale qui est incomplètement segmentée (Haber and Nauta, 1983; Haber and Watson, 1985). Le pallidum ventral projette essentiellement vers le noyau médiodorsal du thalamus (Groenewegen, 1988) qui innerve l aire cingulaire antérieure (Alexander et al., 1986). Comme on le voit, le circuit impliquant les extensions ventrales du striatum et du pallidum est en tout point parallèle au circuit principal décrit ci-dessus (Heimer et al., 1982; Alexander et al., 1986). Les régions de la substance innommée qu occupent les extensions ventrales du striatum et du pallidum abritent également les extensions du noyau amygdalien ou "extended amygdala". En effet, des éléments des noyaux central et médian de l amygdale forment un continuum avec le noyau du lit de la strie terminale (fig. II.4b) via la substance innominée souslenticulaire et envahissant également l accumbens médial (Alheid and Heimer, 1988). La substance innominée est donc constituée des neurones cholinergiques du noyau basal de Meynert, des extensions ventrales du striatum et du pallidum et des extensions du noyau amygdalien. Il peut être difficile de discriminer les constituants des striatum et pallidum ventraux de ceux de 1 "extended amygdala" (cf. paragraphe IV.2.a) (Alheid and Heimer, 1988). IL2.C. Organisation en boucles parallèles Longtemps le système des ganglions de la base a été considéré comme un système d intégration et de convergence des informations corticales multiples qu il reçoit. Cette intégration résultant en une régulation par "feed-back" des diverses fonctions corticales. Cette notion doit être revue à la lumière des données anatomiques et physiologiques émanant principalement du groupe de DeLong et Alexander (Alexander et al., 1986; Alexander and Crutcher, 1990; Delong, 1990). En effet, cette équipe a montré qu il existait plusieurs boucles parallèles calquées sur l organisation cortico-striato-pallido-thalamo-corticale du circuit principal. Chacune de ces boucles engagent des régions spécifiques du cortex cérébral, du striatum, du pallidum, de la substance noire et du thalamus. Cinq de ces boucles ont été caractérisées par cette équipe sans toutefois que les fonctions de chacune d entre elles n aient pu être définies. Elles sont reprises en détail dans la figure II.5.

24 -15- DORSOLATERAL LATERAL ANTERIOR MOTOR OCULOMOTOR PREFRONTAL ORBITOFRONTAL CINQULATE Fig. II.5: Organisation en boucles parallèles du circuit cortico-strioato-pallido-thalamocorticale des ganglions de la base. De Alexander et al., (ACA) aire cingulaire antérieure, (APA) aire prémotrice arcuate, (Caud) corps (b) et tête (h) du noyau caudé, (DLC) cortex préfrontal dorsolatéral, (EC) cortex entorhinal, (FEF) champs orbitaires frontaux, (GPi) globus pallidus interne, (HC) cortex hippocampal, (ITG) gyrus temporal inférieur, (LOF) cortex orbitofrontal latéral, (MC) cortex moteur, (MDpl) noyau dorsomédian pars paralamellaris, (MDmc) noyau dorsomédian pars magnocellularis, (MDpc) noyau dorsomédian pars parvocellularis, (PPC) cortex pariétal postérieur, (Pu) putamen, (SC) cortex somatosensoriel, (SMA) aire motrice supplémentaire, (SNr) substance noire pars reticulata, (STG) gyrus temporal supérieur, (VAmc) noyau ventral antérieur pars magnocellularis, (VApc) noyau ventral antérieur pars parvocellularis, (VLm) noyau ventro-latéral pars medialis, (VLo) noyau ventro-latéral pars oralis, (VP) pallidum ventral, (VS) striatum ventral, (c) caudal, (d) dorsal), (1) latéral, (m) médian, (r) rostral, (v) ventral. La boucle fonctionnellement et cliniquement la mieux connue est la boucle motrice (fig. 11.5) impliquant les aires corticales motrice primaire, motrice supplémentaire et somatosensorielle. La boucle oculomotrice constitue une seconde boucle motrice. La boucle cingulaire antérieure ou boucle limbique est constituée d aires corticales limbiques et des extensions ventrales du striatum et du pallidum (cf. paragraphe II.2.b). Les deux dernières boucles prennent leur origine dans des aires corticales associatives et sont appelées circuits d association. La notion d intégration, de convergence ou "funnelling" qui prévalait auparavant reste néanmoins partiellement valable. En effet, chacune de ces boucles prend son origine dans plusieurs aires corticales alors qu m fine une seule de ces régions corticales reçoit les afférences thalamiques spécifiques de la boucle (fig. II.5). Au sein de chaque boucle parallèle, les régions spécifiques des ganglions de la base intègrent donc les informations provenant de plusieurs aires corticales et régulent l activité de l une d entre elles.

25 d. Relais intrinsèques et voies modulatrices: la voie nigro-striatale et la boucle pallido-sous-thalamo-pallidale La voie nigro-striatale Le mésencéphale ventral abrite les neurones dopaminergiques qui innervent les diverses composantes du striatum. Ils sont localisés dans des structures contiguës appelées selon la classification de Dalhstrôm et Fuxe (1964) A8 ou aire rétrorubrale, A9 ou substance noire pars compacta et AlO ou aire tegmentale ventrale. La substance noire pars compacta forme une fine couche compacte de cellules apposées à la partie dorsale de la pars reticulata. Chez les primates, la couleur noire de ce noyau est liée au contenu en mélanine de ces neurones. La substance noire pars compacta ou A9 et l aire rétrorubrale ou A8 innervent le striatum dorsal (Gerfen et al., 1987; Jimenez-Castellanos and Graybiel, 1987), différentes populations neuronales projettant vers les différents compartiments striataux (Gerfen et al., 1987; Jimenez-Castellanos and Graybiel, 1987) (cf. paragraphe II.5.). L aire tegmentale ventrale ou AlO projette principalement vers les diverses composantes du striatum ventral (Gerfen et al., 1987; Jimenez-Castellanos and Graybiel, 1987) et intervient de ce fait dans la boucle cingulaire antérieure ou boucle limbique. La boucle pallido-sous-thalamo-pallidale Le globus pallidus externe et le noyau sous-thalamique ou corps de Luys sont les deux relais intrinsèques impliqués dans cette boucle. Le globus pallidus externe projette essentiellement vers le noyau sous-thalamique dont il est la principale structure afférente (Carpenter et al., 1981; Smith et al., 1990a). Par ailleurs le noyau sous-thalamique reçoit également des afférences corticales provenant principalement des aires motrices primaire et supplémentaire. Comme dans les autres connexions du système, l organisation topographique de ces projections est préservée. Ce noyau sous-thalamique innerve les noyaux efférents du système des ganglions de la base que sont le globus pallidus interne et la substance noire pars reticulata (Parent and Smith, 1987; Parent et al., 1989; Smith et al., 1990b). Une projection minoritaire est décrite également vers le globus pallidus externe (Parent and Smith, 1987; Parent et al., 1989; Smith et al., 1990b) Histologie Les neurones du striatum peuvent être groupés en cinq classes (Graveland et al., 1985): 1. Les neurones de taille moyenne (15-20 /im de diamètre) dont les arborisations dendritiques sont abondamment pourvues d épines ("spiny neurons type I"). Ces neurones représentent 90 à 95% des neurones striataux chez les rongeurs et 70 à 80% chez les primates. A l opposé des autres types neuronaux dont les axones ne quittent pas le striatum et qui sont donc des interneurones, les "spiny neurons type r sont à l origine des voies efférentes du striatum vers ses structures cibles. 2. Les neurones dont les arborisations dendritiques sont modérément pourvues d épines ("spiny neurons type II"), de taille moyenne (15-20 /im de diamètre) ou de grande taille (20-30 /im de diamètre). 3. Les neurones de taille moyenne (15-20 /im de diamètre) dont les arborisations dendritiques sont dépourvues d épines ("aspiny neurons type I"). Ces neurones représentent moins de 5% des neurones striataux chez les rongeurs et 15 à 20% chez les primates.
im) et possèdent un arbre dendritique organisé en disque perpendiculaire aux afférences striatales (Percheron et al., 1984).")
26 Les grands neurones (25-35 /im de diamètre) dont les arborisations dendritiques sont dépourvues d épines ("aspiny neurons type II"). 5. Les petits neurones (8-15 nm de diamètre). Les neurones pallidaux efférents sont de grande taille (20 à 60 )im) et possèdent un arbre dendritique organisé en disque perpendiculaire aux afférences striatales (Percheron et al., 1984). Leurs dendrites sont entièrement couverts de synapses striato-pallidales démontrées par la microscopie électronique (Fox et al., 1974). Cette disposition des synapses striatopallidales rend compte de l aspect en fibres laineuses ou "woolly fibers" observé après certaines colorations (fig. II.6) (Haber and Eide, 1981; Haber and Nauta, 1983; Haber and Watson 1985). Les dendrites des neurones du pallidum ventral présentent le même aspect (Haber and Nauta, 1983; Haber and Watson 1985). Fig. II.6: Aspect en fibres laineuses ou "woolly fibers" après détection immunohistochimique de substance P dans le globus pallidus interne. Barre d échelle = 20 iim. Les neurones sous-thalamiques sont majoritairement de grande taille (25-40 /im de diamètre). Chaque neurone efférent semble émettre un seul axone qui bifurque en une collatérale ascendante qui se termine dans le globus pallidus interne et une collatérale descendante qui projette vers la substance noire pars reticulata (Parent and Smith, 1987; Smith et al., 1990b). Une population différente de neurones sous-thalamiques innerve le globus pallidus externe (Parent and Smith, 1987; Smith et al., 1990b). Les neurones de la substance noire pars compacta sont de grande taille avec un diamètre moyen de 35 /im et leurs dendrites peuvent fréquemment s étendre dans la pars reticulata.

27 -18- II.4. Neurochimie Outre les neurotransmetteurs dits "classiques", la présence intraneuronale de nombreux neuropeptides a été largement décrite (Bjôrklund and Hôkfelt (Eds), 1985). Les localisations précises de ces neuromédiateurs ainsi que des enzymes spécifiques de synthèse. des neurotransmetteurs "classiques" ont été décryptées dans un premier temps par les techniques immunohistochimiques (cf. paragraphe IV.2.a, article 8). Le clonage des gènes encodant les précurseurs de ces divers peptides et protéines a permis de localiser les cellules exprimant TARN messager spécifique de chacun d entre eux par l utilisation de sondes génétiques et des techniques d hybridation in situ (cf. paragraphes III.2.C, IV.2.b et IV.2.C, articles 6, 9, 10, 11, 12). Par ailleurs, la pharmacologie dans un premier temps, la biologie moléculaire dans un second temps ont permis d identifier les récepteurs et les sous-classes de récepteurs de ces neuromédiateurs. La mise au point de ligands spécifiques marqués permet la localisation de ces récepteurs sur coupes de tissu par autoradiographie (cf. paragraphes III.2.d et IV.2.d, articles 7, 13) ou leur caractérisation sur des préparations membranaires (cf. paragraphe III.2.a, articles 2, 3). De plus, le clonage récent de certains d entre eux a nuancé voire infirmé les classifications pharmacologiques. Comme pour les neuromédiateurs eux-mêmes, ces clonages permettent l identification des cellules exprimant ces divers récepteurs par hybridation in situ (cf. chapitre 111.2, articles 1, 4, 5, 6, 7). Le système des ganglions de la base et en particulier le striatum, se caractérise par sa diversité en neuromédiateurs ainsi qu en leurs récepteurs (pour revue voir Graybiel, 1990). Néanmoins, les neurotransmetteurs "classiques" tels le GABA, l acétylcholine, la dopamine et le glutamate en sont les éléments principaux. Ils peuvent coexister avec une variété de neuropeptides et établir de ce fait des sous-populations neuronales caractérisées par leurs phénotypes neurochimiques au sein de populations morphologiquement homogènes. La figure II.7 synthétise la localisation de ces quatre neurotransmetteurs majeurs dans les différentes connexions du système.
28 -19- Fig. II.7: Neurotransmetteurs classiques dans les connexions majeures du circuit principal des ganglions de la base et de ses deux voies modulatrices. De Graybiel, (ACh) acétylcholine, (DA) dopamine, (GABA) acide gamma-aminobutyrique, (GLU) glutamate, (STN) noyau sous-thalamique, (SNc) substance noire pars compacta, (SNr) substance noire pars reticulata II.4.a. Le striatum Le GABA est le neurotransmetteur majeur du striatum et des voies striato-pallidales et striato-nigrale dont il est l origine (Oertel and Mugnaini, 1984; Mugnaini, 1985; Penny et al., 1986; Smith et al., 1987; Chesselet and Robbins, 1989a; Schiffmann and Vanderhaeghen, 1990). Dans le striatum, plusieurs types de neurones GABAergiques ont été identifiés. La grande majorité d entre eux appartiennent à la classe des neurones efférents "spiny type I" (Smith et al., 1987), mais d autres neurones de taille moyenne, interneurones "aspiny type I", sont également GABAergiques (Bolam et al., 1983). De plus, chez l homme (Schiffmann and Vanderhaeghen, 1990) et le singe (Smith et al., 1987), il existe une population striatale de neurones GABAergiques de grande taille qui correspondent probablement aux neurones "spiny type II" de grande taille. Ces derniers ont pu être caractérisés par l utilisation d anticorps dirigés directement contre le neurotransmetteur GABA (Seguela et al., 1985; Schiffmann et al., 1988; Schiffmann and Vanderhaeghen, 1990) lui-même et non plus contre son enzyme de biosynthèse la glutamic acid decarboxylase (GAD). Les neurones GABAergiques efférents ou "spiny type I" synthétisent également certains neuropeptides (pour revue voir Graybiel, 1990). Il s agit des tachykinines substance P (SP)
 et substance K (SK) (Linderfors et al.")
29 -20- (Graybiel et al., 1981; Haber and Eide, 1981; Haber and Nauta, 1983; Haber and Watson 1985; Christesson-Nylander et al., 1986; Penny et al., 1986; Young et al., 1986; Gerfen and Young, 1988; Chesselet and Robbins, 1989b; Anderson and Reiner, 1990; Besson et al., 1990; Gerfen et al., 1991) et substance K (SK) (Linderfors et al., 1986), des peptides opioïdes endogènes met-enképhaline (ENK) (Graybiel et al., 1981; Aroniri et al., 1984; Haber and Eide, 1981; Haber and Nauta, 1983; Haber and Watson 1985; Penny et al., 1986; Young et al., 1986; Gerfen and Young, 1988; Besson et al., 1990; Gerfen et al., 1991) et dynorphine (DYN) (Cuello and Paxinos, 1977; Haber and Nauta, 1983; Haber and Watson 1985; Christesson-Nylander et al., 1986; Young et al., 1986; Gerfen and Young, 1988; Anderson and Reiner, 1990; Besson et al., 1990; Gerfen et al., 1991) et de la neurotensine (NT) (Sugimoto and Mizuno, 1987). Cependant, ces neuropeptides ne sont pas exprimés par tous les neurones GABAergiques efférents et on peut schématiquement reconnaître deux souspopulations différentes. La première est constituée des neurones GABAergiques exprimant l ENK et la NT et la seconde par ceux exprimant les tachykinines SP et SK et la DYN (Sugimoto and Mizuno, 1987; Anderson and Reiner, 1990; pour revue voir Reiner and Anderson, 1990). De plus et de manière remarquable, ces deux sous-populations ont des cibles différentes. Les neurones GABA-ENK-NT innervent principalement le globus pallidus externe et les neurones GABA-SP-SK-DYN le globus pallidus interne et la substance noire pars reticulata (Cuello and Paxinos, 1977; Christesson-Nylander et al., 1986; Gerfen and Young, 1988; Gerfen et al., 1990). Ceci peut être distingué très aisément par l enrichissement du globus pallidus externe en fibres à ENK et celui du globus pallidus interne en fibres à SP (fig. II.8) (Haber and Eide, 1981; Haber and Watson, 1985). Ces fibres ont l aspect en fibres laineuses ou "woolly fibers" (fig. II.6) caractéristique des fibres striatopallidales (Haber and Eide, 1981; Haber and Watson, 1985). Fig. II.8: Projections striato-pallidales sélectives. Coupes transversales au niveau du striatum caudal humain après détection immunohistochimique de substance P (a) et de met-enképhaline (b). La substance P est abondante dans le globus pallidus interne et la met-enképhaline dans le globus pallidus externe. Les deux peptides sont présents et distribués de manière hétérogène (cf. paragraphe II.5.) dans le putamen. (Ce) noyau central de l amygdale, (GPe) globus pallidus externe, (GPi) globus pallidus interne, (P) putamen, (SI) substance innominée Le striatum contient également des interneurones non-gabaergiques. Les plus importants sont sans conteste les neurones "aspiny type II" dont le neurotransmetteur est
 et la somatostatine (SRIF)")
30 -21- l acétylcholine (Bolam et al., 1984). D autres interneurones appartenant à la classe des neurones "aspiny type I" sont majoritairement non-gabaergiques et contiennent des neuropeptides: le neuropeptide Y (NPY) et la somatostatine (SRIF) (Difiglia and Aronin, 1982; Vincent et al., 1982; Chesselet and Graybiel, 1986; Smith and Parent, 1986a). Ces neurones sont peu nombreux mais ont un rôle local important. Ils ont de plus acquis leurs lettres de noblesse par la découverte de leur résistance relative au processus dégénératif de la maladie de Huntington (Aronin et al., 1983; Dawbarn et al., 1985) ainsi qu à l injection de certaines excitotoxines qui constitue un modèle expérimental de cette pathologie (cf. paragraphe II.6.) (Beal et al., 1986). De nombreux récepteurs sont présents dans le striatum. En plus des récepteurs de la dopamine et du glutamate détaillés ci-dessous (paragraphes II.4.b, et II.4.C.), on y trouve notamment des récepteurs cholinergiques muscariniques Ml et M2, GABA-A et GABA-B, opioïdes de type mu, delta et kappa, NT etc...(pour revue voir Graybiel, 1990) ainsi que les récepteurs Al et A2 de l adénosine (Wojcik and Neff, 1983; Jarvis and Williams, 1989; Alexander and Reddington, 1989; Parkinson and Fredholm, 1990) et CCK-A et CCK-B de la cholécystokinine (Dietl et al., 1987; Dietl and Palacios, 1989; Kritzer et al., 1991) qui sont étudiés dans le présent mémoire (chapitres III et IV) b La voie cortico-striatale Le transmetteur de la voie cortico-striatale est un acide aminé excitateur; le glutamate (Divac et al., 1977; Ottersen and Storm-Mathysen, 1984). Aucun neuropeptide n a pu être détecté dans ces neurones. Néanmoins, la richesse en cholécystokinine du striatum permet d envisager une telle coexistence. Cette question est discutée dans la deuxième partie du présent mémoire (chapitre IV). Les terminaisons glutamatergiques font synapse sur les épines dendritiques des neurones striataux "spiny type I" qui constituent la voie efférente du striatum. Par ailleurs, les afférences glutamatergiques cortico-striatales innervent également les interneurones de grande taille ("aspiny type II") et de taille moyenne ("aspiny type I") (pour revue voir Gerfen, 1988). Différentes classes de récepteurs glutamatergiques ont été décrits par les techniques pharmacologiques parmi lesquels les récepteurs de type NMDA et les récepteurs de type non- NMDA sont présents dans le striatum (Monaghan and Cotman, 1985; Greenamyre and Young, 1989; Samuel et al., 1990). La biologie moléculaire et le clonage d un grand nombre de sous-types de récepteurs glutamatergiques y compris le récepteur métabotropique couplé aux protéines G, compliquent considérablement leur classification (Miller, 1991a,b) C. La voie nigro-striatale (substance noire pars compacta) La voie nigro-striatale utilise la dopamine comme neurotransmetteur classique (Dalhstrôm and Fuxe, 1964). Le striatum est de ce fait la structure cérébrale la plus riche en dopamine (Fuxe, 1965) et également en récepteurs de la dopamine (Joyce et al., 1986; Savasta et al., 1986; Besson et al., 1988; Caboche et al., 1991). Les terminaisons dopaminergiques font synapse sur les épines dendritiques des neurones striataux "spiny type I" en aval des synapses glutamatergiques. Une innervation dopaminergique des interneurones cholinergiques ("aspiny type II") et des terminaisons glutamatergiques est également décrite (pour revue voir Gerfen, 1988). Les neurones nigro-striataux synthétisent également la NT et la cholécystokinine (Hôkfelt et al., 1980a,b; Vanderhaeghen et al., 1980; Seroogy et al., 1987,

31 a,b). Ce point est également discuté au chapitre IV dans la deuxième partie du présent mémoire. L importance des drogues agissant sur les récepteurs de la dopamine en clinique neurologique et psychiatrique expliquent les très nombreuses recherches dont ils sont l objet. La pharmacologie a permis de classer ces récepteurs en deux classes: DI et D2 (Stoof and Kebabian, 1981). La biologie moléculaire et le clonage des gènes encodant ces récepteurs ont démontré qu il existait de multiples sous-types de récepteurs de la dopamine (pour revue voir Sibley and Monsma, 1992): les récepteurs Dl-like: Dla, Dlb ou D5 et les récepteurs D2-like: D2a, D2b, D3 et D4 (Bunzow et al., 1988; Sokoloff et al., 1990; Sunahara et al., 1990, 1991; Tiberi et al., 1991; Van Toi et al., 1991). La signification physiologique de tous ces récepteurs dopaminergiques n est pas définitivement établie. La technique d hybridation in situ a permis de démontrer que les récepteurs Dla et D2 sont très abondamment exprimés dans le striatum et majoritairement exprimés par des sous-populations neuronales différentes: le Dla par les neurones GABA-SP striato-nigraux et le D2 par les neurones GABA-ENK striato-pallidaux (Gerfen et al., 1990; Le Moine et al., 1990, 1991). La dopamine stimule les neurones GABA-SP striato-nigraux par l activation du récepteur Dla alors qu elle inhibe les neurones GABA-ENK striato-pallidaux par celle du D2 (Gerfen et al., 1990, 1991) d. Le globus pallidus et la substance noire pars reticulata Le GABA est le neurotransmetteur des neurones efférents du pallidum et de la substance noire pars reticulata (Mugnaini and Oertel, 1985; Smith et al., 1987, 1990a) et donc des voies pallido- et nigro-thalamiques et pallido-luysiennes. Le globus pallidus interne est riche en fibres à GABA et SP et le globus pallidus externe en fibres à GABA et ENK, d origine striatale (fig. 11.8) (Haber and Eide, 1981; Haber and Nauta, 1983; Haber and Watson 1985). Le pallidum ventral est constitué du même type de neurones GABAergiques. Néanmoins, la ségrégation des "woolly fibers" ENK et SP est moins nette que celle observée dans les subdivisions externe et interne du pallidum dorsal, indiquant que la division du pallidum ventral est incomplète (Haber and Nauta, 1983; Haber and Watson 1985) e. Le noyau sous-thalamique Le glutamate est le neurotransmetteur des neurones efférents du noyau sous-thalamique (Smith and Parent, 1988). Ces derniers reçoivent une innervation GABAergique du globus pallidus externe (Smith et al., 1990a). ILS. Compartimentalisation du striatum Le striatum a été longtemps considéré comme une structure homogène. Trois éléments décrits dans les années 1970 démontreront qu il n en est rien et que cette structure présente au contraire une organisation hétérogène mais structurée. Oison et al. (1972) ont en effet démontré qu au cours du développement du striatum certaines régions, îlots ou "patches" sont enrichis en dopamine. Peu après, Graybiel and Ragsdale (1978), et Herkenham and Pert (1981) rapportent également une distribution hétérogène dans le striatum de l acétylcholine estérase (AChE) et des récepteurs opioïdes de type mu. L AChE est peu abondante dans certaines zones striatales ou "patches" alors qu au contraire ces zones ou "patches" sont considérablement enrichies en récepteurs opioïdes de type mu (Graybiel and Ragsdale 1978; Herkenham and Pert, 1981) (fig. II.9).
 et immunohistochimiquement pour celle de met-enképhaline (b).")
32 -23- Fig. II.9: Compartimentalisation du striatum. Coupes transversales au niveau du striatum humain rostral marquées histochimiquement pour la détection de l acétylcholine estérase (a) et immunohistochimiquement pour celle de met-enképhaline (b). (c) Coupe transversale de télencéphale de rat comprenant le striatum rostral et montrant la distribution des récepteurs opioïdes de type mu. Les flèches indiquent des striosomes pauvres en acétylcholine estérase chez l homme, des striosomes riches en récepteurs opioïdes mu chez le rat, et la richesse en met-enképhaline à la limite entre les striosomes et la matrice. (C) noyau caudé, (P) putamen La reproductibilité de ces résultats a poussé les auteurs à définir deux compartiments striataux; les "patches" ou striosomes qui sur des coupes transversales ont une forme ronde ou elliptique et la matrice extrastriosomale. Le compartiment striosomal occupe 10 à 20% du volume striatal. De nombreuses études ont par la suite permis d étendre cette notion et ce principalement grâce aux travaux des équipes de Graybiel, Gerfen et Glowinski (Graybiel and Ragsdale, 1978, 1981; Graybiel et al., 1981; Gerfen 1984; Graybiel, 1984; Gerfen, 1985; Malach and Graybiel, 1986; Gerfen et al., 1987; Graybiel et al., 1987; Jimenez-Castellanos and Graybiel, 1987; Besson et al., 1988; Gerfen and Young, 1988; Ragsdale and Graybiel, 1988; Desban et al., 1989; Gerfen 1989; Nastuk and Graybiel, 1989; Graybiel 1990; Kernel et al., 1989; Krebs et al., 1991). En effet, depuis la définition neurochimique initiale, striosomes pauvres en AChE et riches en récepteurs opioïdes de type mu, la compartimentalisation de neurotransmetteurs, enzymes de métabolisme de neurotransmetteurs.
. Les neurones GABAergiques pris dans leur ensemble ne démontrent pas de compartimentalisation.")
33 -24- neuropeptides, ARN messagers spécifiques de neurotransmetteurs ou neuropeptides, récepteurs, sites de recapture ("uptake sites") et d autres substances a été décrite (pour revue voir Graybiel,, 1990). Les neurones GABAergiques pris dans leur ensemble ne démontrent pas de compartimentalisation. Par contre, la distribution de la SP présente un enrichissement striosomal en neurones et fibres (fig. li.s.a). Celle d ENK est plus complexe, il semble que les corps cellulaires soient plus abondants dans la matrice mais que les striosomes soient plus riches en fibres (pour revue voir Graybiel,, 1990). Chez l homme, un tel enrichissement en fibres est observé à la jonction entre les striosomes et la matrice résultant en une image en anneau (fig. II.9.b). L étude par hybridation in situ des neurones exprimant les ARN messagers de la SP et de l ENK n a pas encore permis de trancher définitivement la question. En effet, Gerfen and Young (1988) n observent pas de différences striosomes/matrice alors que Chesselet and Robbins (1989b) démontrent une expression plus importante de SP dans les neurones striosomaux. Une reconstruction tridimensionnelle à partir de séries de coupes parallèles a démontré que les striosomes constituent en fait un système de canaux orientés rostro-caudalement ou médio-ventralement (Desban et al., 1989). De plus, la localisation de ces canaux au sein du striatum semble comparable d un individu à l autre (Desban et al., 1989). Par leurs prolongements, les interneurones cholinergiques ainsi que les interneurones à SRIF et NPY principalement présents dans la matrice jouent un rôle de connexion entre matrice et striosomes. La signification fonctionnelle de cette hétérogénéité intrinsèque est renforcée par une hétérogénéité des afférences et efférences qui correspond à la compartimentalisation neurochimique intrinsèque décrite (Herkenham and Pert, 1981; Gerfen 1984, 1985, Malach and Graybiel, 1986; Gerfen et al., 1987; Jimenez-Castellanos and Graybiel, 1987; Ragsdale and Graybiel, 1988). En effet, différents groupes neuronaux dopaminergiques du mésencéphale projettent vers la matrice et les striosomes (Gerfen et al., 1987; Jimenez- Castellanos and Graybiel, 1987). Les projections cortico-striatales innervant la matrice et les striosomes ont également différentes origines: les régions limbiques projetant préférentiellement vers les striosomes (Gerfen, 1984). De plus, il a également été démontré que les neurones pyramidaux de là couche V du cortex projettent de manière prédominante dans la matrice alors que ceux de la couche III projettent vers les striosomes (Gerfen, 1989). Les neurones des striosomes projettent eux principalement vers la substance noire pars compacta alors que les neurones de la matrice projettent vers les structures cibles classiques du striatum que sont le globus pallidus et la substance noire pars reticulata (Gerfen, 1985). Il est cependant important de noter que cette compartimentalisation se caractérise le plus souvent par un enrichissement ou un appauvrissement relatif (fig. II.9) et très rarement par l absence complète d un marqueur dans un compartiment. Un même neurotransmetteur peut avoir une fonction différente selon le compartiment. Ainsi, le glutamate par stimulation de ses récepteurs NMDA (Krebs et al., 1991) et l acétylcholine (Kernel et al., 1990) modulent différemment la libération de dopamine dans les striosomes et la matrice. De même, la régulation de l expression de dynorphine par la dopamine est différente dans les deux compartiments (Gerfen et al., 1991).
34 -25- II.6. Rôle des ganglions de la base dans la physiologie du mouvement et du comportement. Applications en pathologie. Le modèle simple utilisé pour comprendre les processus de contrôle du mouvement par les ganglions de la base est celui de la désinhibition (Chevalier and Deniau, 1990). Ce mécanisme résulte de l existence de deux synapses GABAergiques inhibitrices en série: la projection du striatum vers le globus pallidus interne et la substance noire pars reticulata et la projection de ces dernières structures vers le thalamus. Au repos, les neurones du striatum sont majoritairement silencieux alors que ceux du globus pallidus interne et de la substance noire pars reticulata sont toniquement actifs à environ 100 "spikes''/sec. Une stimulation des neurones striataux par leurs afférences corticales entraîne une inhibition phasique des neurones toniquement actifs du globus pallidus interne et de la substance noire pars reticulata et de ce fait une désinhibition ou excitation des neurones thalamo-corticaux (fig. IL 10). A SNr Fig. IL 10: Mécanisme de déshinibition. Séquence des événements éléctrophysiologiques, illustrée par des histogrammes de fréquences d activité, menant à la déshinibition du thalamus après injection de glutamate dans le striatum (flèche). De Chevalier et Deniau, (SNr) substance noire pars reticulata, (VM) noyau ventromédian du thalamus. Néanmoins, ce mécanisme ne génère pas la commande motrice elle-même, mais spécifie plutôt la direction, la graduation de la force engagée et donc l amplitude et la vitesse; la séquence des mouvements ayant lieu par la sélection des neurones moteurs qui doivent

35 -26- décharger. Dans ce modèle, l exécution d une réponse motrice nécéssite une coïncidence temporelle entre cette désinhibition évoquée par les noyaux de la base et des signaux de commande d autres sources. Plus largement, le système des ganglions de la base peut jouer le rôle de moniteur ou comparateur de l activité motrice. L information de commande motrice provenant du cortex prémoteur serait comparée avec l information proprioceptive concernant un mouvement en cours. Cette information proprioceptive est en effet susceptible de provoquer une réponse striatale même pendant l exécution de ce mouvement. Ce rôle de comparateur serait important pour l analyse des conséquences d un mouvement ou la programmation motrice future. Les différentes pathologies dégénératives des ganglions de la base que sont par exemple les maladies de Parkinson et de Huntington sont caractérisées par des anomalies de programmation, de vitesse, d amplitude des mouvements. Divers modèles animaux de ces pathologies ont été utilisés: lésion sélective des neurones dopaminergiques de la substance noire pars compacta comme modèle de Parkinson par injection de 6-hydroxydopamine (pour revue voir Zigmond, 1990) ou de MPTP; lésion sélective des neurones efférents du striatum comme modèle de Huntington par injection intrastriatale d excitotoxines (acides quinolinique ou iboténique) (Beal et al., 1986). Les données neurochimiques et neuroanatomiques obtenues par l étude du système des ganglions de la base normal, dans les modèles expérimentaux de pathologies (Crossmann, 1987) et chez les patients atteints de ces pathologies a permis à Albin et al. (1989) de proposer un modèle explicatif des pathologies hypokinétiques et hyperkinétiques (fig ). Celui-ci est basé principalement sur l existence de deux sous-populations neuronales striatales: les neurones striato-nigraux GABA-SP et les neurones striato-pallidaux GABA-ENK. Ceux-ci sont à l origine de deux voies différentes vers le thalamus: l une excitatrice (désinhibitrice) pour les neurones striato-nigraux GABA-SP et l autre inhibitrice pour les neurones striato-pallidaux GABA-ENK. De plus, comme décrit plus haut (cf. paragraphe II.4), la dopamine stimule les neurones striato-nigraux GABA-SP et inhibe les neurones striato-pallidaux GABA-ENK. La lésion dopaminergique entraîne donc une hypoactivité de la boucle excitatrice et une hyperactivité de la boucle inhibitrice provoquant une extinction de l activité thalamique rendant compte de l hypokinésie (fig. II. 11). La levée d un parkinsonisme expérimental par une lésion du noyau sous-thalamique confirme l hypothèse d hyperactivité de la boucle inhibitrice (Bergman et al., 1990). L atteinte relativement sélective de la population striatale GABA-ENK par rapport aux neurones GABA-SP dans la maladie de Huntington (Reiner et al., 1988) et la lésion du noyau sous-thalamique entraînent une hypoactivité de la boucle inhibitrice provoquant une augmentation de l activité thalamique rendant compte de l hyperkinésie (fig ); choréoathétose dans la première affection et hémiballisme dans la seconde. L implication du système des ganglions de la base dans le contrôle de comportements plus complexes est généralement admise (Phillips and Carr, 1987). Les boucles préfrontales et limbique (cf. paragraphe II.2.c) jouent un rôle dans l exécution de taches comportementales. Il a été proposé que les troubles cognitifs observés dans la maladie de Huntington soient partiellement liés à la pathologie striatale et plus particulièrement du noyau caudé, plutôt qu à une pathologie corticale surajoutée (Folstein et al., 1990). Enfin des anomalies neurochimiques ont été observées dans des pathologies psychiatriques telles que la schizophrénie dans laquelle une hyperactivité dopaminergique relative est suggérée dans le striatum ventral (Wong et al., 1986).
36 Normal Hemiballism Eaiiy Stage Huntington's Disease

37 -28- III. PREMIERE PARTIE: LOCALISATION ET FONCTION DES RECEPTEURS DE L ADENOSINE. IDENTIFICATION DES GENES ENCODANT LES RECEPTEURS Al ET A2 Localisation de TARN messager encodant le récepteur A2 de l adénoisne dans le cerveau de rat au niveau du striatum. Image en pseudo-couleur, le rouge correspondant aux régions les plus riches.
38 -29- Les données discutées dans le présent chapitre sont exposées dans les articles suivants; 1. SCHIFFMANN, S.N., LIBERT, F., VASSART, G., DUMONT, J.E. and VANDERHAEGHEN, J-J. : A cloned G protein-coupled protein with a distribution restricted to striatal medium-sized neurons. Possible relationship with DI dopamine receptor. Brain Res., 519, , MAENHAUT,C., VAN SANDE, J., LIBERT, F., ABRAMOVICZ, M., PRAMENTIER, M., VANDERHAEGHEN, J.-J., DUMONT, J.E., VASSART, G. and SCHIFFMANN, S.N.: RDC8 codes for an A2 receptor with physiological constitutive activity. Biochem. Biophys. Res. Comm. 173(3), , LIBERT, F., SCHIFFMANN, S.N., LEFORT, A., PARMENTIER, M., GERARD, C., DUMONT, J.E., VANDERHAEGHEN, J.-J. and VASSART, G.: The orphan receptor cdna RDC7 encodes an Al adenosine receptor. EMBO J., 10: , (Note: F. Libert and S.N. Schiffmann contributed equally to this study.) 4. SCHIFFMANN, S.N., LIBERT, F., VASSART, G. and VANDERHAEGHEN, J-J. : Distribution of adenosine A2 receptor mrna in the human brain. Neurosci. Lett., 130: , SCHIFFMANN, S.N., JACOBS, O.P. and VANDERHAEGHEN, J.-J.: Striatal Restricted Adenosine A2 Receptor (RDC8) Is Expressed by Enkephalin but Not by Substance P Neurons: An In Situ Hybridization Histochemistry Study. J. Neurochem., 57: , SCHIFFMANN, S.N. and VANDERHAEGHEN J.-J.: Adenosine A2 receptors regulate the gene expression of striatopallidal and striatonigral neurons. Submitted, SCHIFFMANN, S.N. and VANDERHAEGHEN J.-J.: Ontogeny of Gene Expression of Adenosine A2 Receptor in the Striatum; Early Localization in the Patch Compartment. J. Comp. Neurol., in press, 1992

39 -30- III.l. Introduction et Objectifs Deux purines, l adénosine et l ATP, sont reconnues actuellement comme des neuromodulateurs dans les systèmes nerveux central et/ou périphérique (pour revues voir Stone, 1981; Dunwiddie, 1985; Snyder, 1985). Néanmoins, la participation de l adénosine à de multiples fonctions métaboliques cellulaires rend difficile l identification d un "pool" neuromodulateur et explique également l homogénéité des concentrations mesurées dans les diverses régions cérébrales. La production d adénosine et sa libération neuronale, synaptique ou non-synaptique, reste un sujet non résolu. En effet, s il est bien établi qu une dépolarisation induit une libération d adénosine, il reste à démontrer que celle-ci n est pas secondaire à une libération d ATP qui serait directement dégradé en adénosine par une 5 nucléotidase ectomembranaire. L adénosine pourrait également provenir de la dégradation de l ATP juste avant sa libération. Un système de recapture par un transporteur spécifique dont un antagoniste est le dipyridamole, ainsi que dans une moindre mesure la présence intracellulaire d un enzyme dégradant l adénosine, l adénosine déaminase, assurent l inactivation de cette molécule, comme cela est décrit pour les autres neurotransmetteurs (pour revues voir Stone, 1981; Dunwiddie, 1985; Snyder, 1985). L adénosine a également des effets différents de ses fonctions métaboliques cellulaires dans divers systèmes non-neuronaux. L adénosine agit par sa liaison à des récepteurs spécifiques. Comme pour toute molécule participant aussi au métabolisme cellulaire et ayant donc une distribution ubiquitaire, la spécificité et la distribution de ses récepteurs est d importance pour comprendre les fonctions neuromodulatrices de l adénosine. Les récepteurs de l adénosine ont été subdivisés en deux classes: les récepteurs Al et A2. Cette classification a été déterminée à l origine par la capacité de l adénosine d inhiber et de stimuler par sa liaison respectivement aux récepteurs Al et A2 la production d AMP cyclique par l adénylate cyclase (Van Calker et al., 1979). Le couplage de ces récepteurs à d autres effecteurs n est pas exclu et a été récemment démontré pour le récepteur Al (Trussel and Jackson, 1985). Le développement d agonistes et d antagonistes a permis de caractériser la pharmacologie de ces deux récepteurs (Bruns et al., 1981, 1986). De plus, l existence d un récepteur A3 (Riberio and Sébastian, 1986) et de soustypes A2a et A2b du récepteur A2 a été proposée sur base de différentes données pharmacologiques (Bruns et al., 1986; Van Calker et al., 1991). Le sous-type A2b serait, comparé au récepteur A2a, un récepteur de plus basse affinité. Le récepteur A3 serait un récepteur de l adénosine régulant la libération de neurotransmetteur à la jonction neuromusculaire et présentant une pharmacologie qui le différencie des types Al et A2 (Riberio and Sébastian, 1986). Les antagonistes classiques des récepteurs Al et A2 de l adénosine sont les méthylxanthines parmi lesquelles la caféine et la théophyline (Bruns et al., 1986). La présence de certaines de ces molécules dans diverses boissons à effet psychotrope à des concentrations compatibles avec une action in vivo via les récepteurs de l adénosine met en lumière l importance potentielle de ceux-ci. Il est intéressant de noter que ces xanthines ont tout d abord été reconnues comme inhibiteurs de la phosphodiesterase et que l on attribuait leurs effets physiologiques à cette inhibition. Cependant les concentrations requises pour inhiber la phosphodiesterase étant très largement supérieures à celles requises pour agir comme antagonistes des récepteurs de l adénosine, il est hautement probable que ce dernier mécanisme constitue le principal mode d action de ces molécules. L effet majeur de l adénosine dans le système nerveux est globalement inhibiteur. Celuici est médié principalement par des récepteurs Al situés présynaptiquement sur des
.")
40 -31- terminaisons axonales utilisant une grande variété de neurotransmetteurs. A ce niveau, la stimulation du récepteur Al entraîne une inhibition de la libération de ces neurotransmetteurs (Fredholm and Dunwiddie, 1988). Ce mécanisme a notamment été démontré pour le GABA, la dopamine, l acétylcholine et le glutamate (Fredholm and Dunwiddie, 1988). Une action inhibitrice de l adénosine est aussi médiée par des récepteurs Al postsynaptiques. Cet effet inhibiteur de l adénosine via son(es) récepteur(s) Al permet d évoquer un rôle de neuroprotection de l adénosine dans des circonstances physiopathologiques telles que l ischémie et l épilepsie (Dragunow and Faull, 1988; Chin, 1989). Cette action neuroprotectrice a en effet été démontrée dans divers modèles expérimentaux reproduisant ces deux conditions (Petersen, 1991; Von Lubitz et al., 1988). L absence jusqu en 1989 d agonistes sélectifs du récepteur A2 (Jarvis et al., 1989) explique que ses fonctions ont été moins bien étudiées. Néanmoins, les interactions de ce récepteur avec le système dopaminergique ont suggéré son intervention dans le contrôle du mouvement ou de l activité locomotrice chez les rongeurs (Fuxe and Ungerstedt, 1974; Snyder et al., 1981; Green et al., 1982; Jarvis and Williams, 1987; Durcan and Morgan, 1989; Ferre et al., 1991a). En effet, divers modèles comportementaux chez l animal indiquent l existence d une telle interaction entre les récepteurs A2 et les récepteurs dopaminergiques D2 (Ferre et al., 1991a). Dans ces modèles, une stimulation du récepteur A2 évoque la même réponse comportementale qu un antagoniste D2 (Ferre et al., 1991a). Les articles 1, 4, 5, 6 et 7 du présent chapitre apportent des données nouvelles et discutent la(es) fonction(s) de ce récepteur dans le système des ganglions de la base (cf. paragraphes III.2.b, III.2.C et III.2.d). La compréhension des mécanismes de liaison des médiateurs à leurs récepteurs et des mécanismes d activation des systèmes effecteurs est grandement améliorée par la connaissance précise de la structure primaire de ces récepteurs. Cette approche est favorisée par le développement des techniques de biologie moléculaire et de clonage. La compréhension de ces mécanismes par ces nouveaux outils devraient permettre de développer de nouvelles molécules agonistes ou antagonistes plus sélectives des récepteurs ainsi caractérisés. Les clonages des gènes encodant ces récepteurs permettent également de développer des méthodes d analyse des modifications de leur expression dans diverses conditions physiopathologiques et d étudier leur expression à l échelle cellulaire. Les récepteurs Al et A2 de l adénosine appartiennent à la famille très large des récepteurs couplés aux protéines G (Birnbaumer et al., 1990). Le clonage des deux premiers récepteurs de cette famille, la rhodopsine et le récepteur beta-2 adrénergique (Nathans and Hogness, 1983; Dixon et al., 1986), a démontré qu ils avaient une structure monomérique caractérisée par sept régions hydrophobes transmembranaires, une extrémité amino-terminale et trois boucles extracellulaires, et une extrémité carboxy-terminale et trois boucles intracellulaires (cf. fig. III.3 et III.4). Le clonage d autres membres de la famille a démontré une relative conservation de séquence dans les régions transmembranaires (Libert et al., 1991b) (cf. fig. III.3 et III.4). Cette caractéristique a été mise à profit pour le clonage de récepteurs orphelins. En effet, Libert et al. (1989) ont défini deux séries d oligonucléotides dégénérés complémentaires des troisième et sixième régions transmembranaires en compilant les séquences des récepteurs beta-1, beta-2 et alpha-2 adrénergiques, muscarinique ml, sérotinergique 5-HTla et de la substance K. Des cdnas obtenus par la transcription inverse d ARN poly-adénylés de thyroïde humaine ont été soumis à une amplification par PCR ("polymerase chain reaction) à l aide de ces "primers". Après sous-clonage des produits PCR
41 -32- obtenus, une librairie de cdna de thyroïde de chien a été criblée pour obtenir des clones complets. Parmi les clones obtenus, quatre d entre eux, RDCl, RDC4, RDC7 et RDC8, encodent des membres alors inconnus ou "orphelins" de la famille des récepteurs couplés aux protéines G (Libert et al., 1989). Dans le présent chapitre, notre but a été tout d abord d identifier ces récepteurs orphelins RDC7 et RDC8 par la caractérisation de leurs distributions cérébrales par hybridation in situ (paragraphe III.2.a article 1) et par la caractérisation des paramètres de liaison spécifique de ligands radiomarqués et sélectifs de récepteurs donnés et des systèmes effecteurs auxquels ces récepteurs sont couplés après leur expression dans divers systèmes cellulaires (paragraphe III.2.a, articles 2, 3). Identifiés et caractérisés comme étant les récepteurs Al et A2 de l adénosine, nous disposons de nouveaux outils pour étudier par hybridation in situ et Northern blot d ARN la distribution de l ARN messager encodant le récepteur A2 de l adénosine dans les ganglions de la base de plusieurs espèces y compris l homme, à l age adulte et au cours du développement. Cette approche a permis également de caractériser les neurones striataux exprimant le récepteur A2a de l adénosine sur base de leur phénotype neurochimique et de leur(s) site(s) de projection (paragraphes III.2.b et III.2.d, articles 1,4,5,6,7). Les diverses données ainsi obtenues nous ont permis de poser des hypothèses fonctionnelles que nous avons vérifié par l étude du(des) rôle(s) des récepteurs A2 de l adénosine dans la régulation de l expression génétique dans le striatum (paragraphe III.2.C, article 6).

42 -33- III. 2. RESULTATS ET DISCUSSION III.2.a Identification et caractérisation de RDC7 et RDC8 comme récepteurs Al et A2 de l adénosine (articles 1, 2, 3) L identification de récepteurs orphelins doit être abordée par deux voies parallèles. L étude de leur distribution anatomique précise par hybridation in situ et par Northern blotting d ARN permet la comparaison avec les données pharmacologiques antérieures. Leur expression dans des systèmes cellulaires (cellules Cos-7, CHO, Yl, oocytes de Xenopes) fournit le matériel adéquat pour l étude des paramètres de liaison spécifique, du couplage aux systèmes effecteurs et de l activation spécifique de canaux ioniques. Ces deux voies d approche ont été utilisées pour l identification de RDC7 et RDC8 et ont toutes deux fourni des résultats déterminants. Libert et al. (1989) ont montré que l expression des récepteurs orphelins RDC7 et RDC8 étudiée par Northern blotting d ARN dans une série de 9 tissus est relativement limitée au cerveau. Au vu de ces localisations sélectives, la distribution de ces deux récepteurs dans le cerveau de chien a été étudiée par hybridation in situ et confirmée par Northern blotting d ARN (article 1). Deux sondes oligonucléotidiques par récepteur sont utilisées et donnent le même résultat. Elles sont complémentaires de régions spécifiques des séquences de RDC7 et RDC8 dans la troisième boucle intracellulaire et l extrémité carboxy-terminale. La distribution de RDC8 dans le cerveau de chien est remarquable. En effet, l ARN messager encodant RDC8 n est détecté que dans le système striatal incluant les noyaux caudé, putamen et accumbens et le tubercule olfactif (fig. 1, article 1). Aucune autre région cérébrale étudiée ne démontre de positivité détectable par nos techniques. A l échelle microscopique, environ 50-60% des neurones de taille moyenne expriment ce récepteur (fig. 1, article 1). Les cellules gliales et les grands neurones qui représentent probablement les interneurones cholinergiques ("aspiny neurons type II") ne sont pas marqués. L important pourcentage de neurones de taille moyenne exprimant ce récepteur implique que, pour le moins, il inclut une proportion significative de neurones efférents ("spiny neurons type I). Ce résultat obtenu par hybridation in situ est confirmé par la comparaison de l expression de RDC8 dans le striatum et le cortex cérébral par Northern blotting d ARN. Une bande unique de taille attendue, 2,4 kb (Libert et al., 1989), est présente dans le striatum alors qu aucun signal n est détecté dans le cortex. De plus, des conditions d hybridation et de rinçage moins sévères ont permis la détection d ARN messagers encodant RDC8 sur des coupes de cerveau de rat mais uniquement avec une des deux sondes complémentaires de la séquence du RDC8 canin. Chez le rat également, ce récepteur est exclusivement exprimé par les neurones de taille moyenne du système striatal: caudé-putamen, accumbens et tubercule olfactif (fig. 1, article 1 et fig. III. 1).

43 -34- Fig. III. 1: Par hybridation in situ, TARN messager encodant RDC8 est exclusivement détecté dans le striatum. (ACC) accumbens, (C-P) caudé-putamen, (TO) tubercule olfactif. La distribution de RDC7 est beaucoup plus étendue. En effet les premiers résultats démontrent l expression de RDC7 dans les couches II-III et V-VI du cortex cérébral, le claustrum, la couche pyramidale de la corne d Ammon de l hippocampe, la couche granulaire du gyrus dentate, l amygdale et faiblement dans le striatum (fig. 1, article 1). Ceci est confirmé par l analyse par Northern blotting d ARN de cortex cérébral, de corne d Ammon et de striatum. Des résultats ultérieurs ont permis d étendre cette distribution à divers noyaux thalamiques (fig. III.2) et à la couche granulaire du cervelet. Fig. III.2: Expression de l ARN messager encodant RDC7 dans la couche pyramidale de l hippocampe (Py), la couche granulaire du gyrus dentate (DG), le cortex cérébral (CX), le cortex piriforme (Pir), le thalamus (TH) et l amygdale (AMY) détectée chez le chien par hybridation in situ.

44 -35- La distribution de RDC8 remarquablement sélective du système striatal nous a amené à proposer une première hypothèse sur la nature de ces deux récepteurs. En effet, le striatum étant particulièrement enrichi en dopamine et donc en récepteurs dopaminergiques, ceux-ci ont constitué une hypothèse séduisante. Le récepteur dopaminergique D2 étant déjà cloné à cette phase du travail (Bunzow et al., 1988), nous avons proposé que RDC8 encode un récepteur Dl. De plus, l homologie structurelle de RDC7 et RDC8, nous a amené à proposer que RDC7 et RDC8 encodent deux sous-types de récepteurs Dl. Des données pharmacologiques antérieures suggéraient déjà l existence de deux sous-types de récepteurs Dl dont l un serait plutôt striatal et l autre présent dans le cortex cérébral, l hippocampe et l amygdale. Nous étions néanmoins déjà conscients à ce stade que divers arguments structurels tels que la faible homologie avec le récepteur D2 déjà cloné et l absence d un résidu aspartate caractéristique de tous les récepteurs aminergiques dans le troisième domaine transmembranaire, allaient à l encontre de cette hypothèse. Bien qu effectivement cette hypothèse se soit avérée fausse (cf. ci-dessous), les résultats récents de diverses équipes impliquées dans le clonage de récepteurs dopaminergiques confirment qu il existe bien deux sous-types de récepteurs Dl, le Dla et le Dlb ou D5 (Sunahara et al., 1990, 1991; Tiberi et al., 1991). De plusj le Dla a une distribution essentiellement striatale alors que le Dlb est exprimé dans des régions extrastriatales telles que l hippocampe et l amygdale (Tiberi et al., 1991). Outre les récepteurs dopaminergiques, un autre récepteur possède la caractéristique d être particulièrement abondant dans le système striatal: le récepteur A2 de l adénosine (Wojcik and Neff, 1983; Bruns et al., 1986, Alexander and Reddington, 1989; Jarvis and Williams, 1989; Parkinson and Fredholm, 1990). Celui-ci est donc un excellent candidat pour l identité de RDC8. Néanmoins, des données physiologiques et/ou pharmacologiques ont démontré la présence de récepteurs A2 dans de nombreuses autres régions cérébrales et dans de nombreux tissus périphériques (Bruns et al., 1986). Nous n éliminons cependant pas cette hypothèse sur base de cette caractéristique. D une part, divers arguments pharmacologiques et physiologiques ont suggéré l existence de deux sous-types de récepteurs A2, A2a et A2b, le récepteur A2a étant le récepteur striatal (Bruns et al., 1986; Van Calker et al., 1991). Dans ce cas, avec les sondes utilisées, le récepteur A2b pourrait ne pas être mis en évidence dans les régions cérébrales extrastriatales où l on soupçonne sa présence. D autre part. Bruns et al. (1986) ont démontré que le nombre de récepteurs A2 dans divers tissus périphériques et dans diverses régions cérébrales extrastriatales n excède jamais 10% du nombre de ces récepteurs dans le striatum. Un faible niveau d expression dans tout tissu ou région cérébrale extrastriatale pourrait rendre celui-ci indétectable s il est en-dessous du seuil de détection de nos techniques. Cette hypothèse basée sur des données anatomiques a permis d expliquer les premières données d expression dans des systèmes cellulaires (article 2). En effet, dans divers modèles cellulaires, cellules Yl, cellules thyroïdiennes de chien en culture primaire et oocytes de Xenopes, l injection d ARN messagers encodant RDC8 résulte en une activation de l adénylate cyclase et de la cascade dépendante de l AMP cyclique sans addition d agent exogène. Toute cellule libère de l adénosine dans le milieu extracellulaire et de plus des préparations de membranes sont elles aussi susceptibles de libérer de l adénosine (Stone, 1981). L activation de RDC8 par cette adénosine endogène peut rendre compte de ses résultats surprenants si l on retient l hypothèse que RDC8 encode le récepteur A2 de

45 -36- l adénosine. Ceci est définitivement confirmé par l observation d une réduction majeure de la stimulation de l adénylate cyclase par l addition dans le milieu soit d adénosine déaminase soit d antagonistes du récepteur A2. Cet effet est par contre reproduit, en présence d adénosine déaminase, par un agoniste non-hydrolysable du récepteur A2; le N- ethylcarboxamideadénosine (NECA). Enfin, une liaison spécifique d un agoniste A2 radiomarqué ([^H]NECA) est démontrée sur une préparation de membranes de cellules Cos-7 transfectées par une construction plasmidique comprenant le cdna encodant RDC8. Ces différents éléments nous ont permis de conclure que RDC8 encode le récepteur A2 de l adénosine. RDC8 et RDC7 possèdent une homologie de 51% dans les régions transmembranaires et plusieurs caractéristiques communes parmi lesquelles une courte extrémité amino-terminale dépourvue de sites putatifs de N-glycosylation (fig. III.3, III.4). Cette proximité structurelle nous permit de proposer l hypothèse selon laquelle RDC7 encode le récepteur Al de l adénosine. Par ailleurs, la distribution cérébrale de RDC7 dans des structures de type cortical, le thalamus et le cervelet cadre également avec cette hypothèse. Pour vérifier celle- ci, des préparations de membranes de cellules Cos-7 transfectées par une construction plasmidique comprenant le cdna encodant RDC7 sont utilisées pour caractériser les paramètres de liaison spécifique d un agoniste Al sélectif radiomarqué ([^HJcyclohexyladénosine) (CHA) (article 3). Des préparations de membranes de cortex cérébral de chien servent de source de récepteur Al naturel. Ces deux préparations de membranes lient de manière spécifique et saturable le CHA tritié et ces liaisons démontrent des Kd similaires. Des expériences de compétition avec différents agonistes (cyclopentyladénosine (CPA), NECA, 2-chloroadénosine (2-CADO)) démontrent un ordre identique de leur capacité de déplacement de la liaison de CHA tritié et des Ki similaires pour les deux types de préparations. Une approche équivalente par liaison spécifique d un agoniste A2 sélectif radiomarqué ([^H]CGS21680) sur des préparations de membranes de cellules Cos-7 transfectées par une construction plasmidique comprenant le cdna encodant RDC8 et de striatum de chien comme source de récepteur A2 naturel, permet la caractérisation définitive de RDC8 comme récepteur A2 et établit la comparaison entre ces deux récepteurs. Les variations d espèces des différents paramètres étudiés justifient la comparaison des récepteurs canins naturels et recombinants (Ferkany et al., 1986; Stone et al., 1988). Enfin, la transfection stable de RDC7 dans des cellules CHO démontre une inhibition de l activité adénylate cyclase de ces cellules après stimulation par un agoniste Al sélectif (CPA). Ce couplage négatif à l adénylate cyclase est également un comportement attendu d un récepteur Al de l adénosine (Van Calker et al., 1979). Ces divers éléments expérimentaux démontrent que RDC7 et RDC8 encodent respectivement les récepteurs Al et A2 de l adénosine. Ces résultats n excluent pas l existence d autre sous-types de récepteurs de l adénosine. En effet, la localisation sélective dans le striatum de RDC8 suggère fortement qu il encode le récepteur A2a. De plus la localisation chromosomique des gènes des récepteurs de l adénosine suggère également l existence d un autre sous-type de récepteur A2. En effet, avec une sonde RDC8, deux pics d hybridation sont observés sur deux chromosomes différents, un pic majeur sur le 11 (llqll-llql3) et un pic mineur sur le 10 (10q q26.3) (Libert et al., 1991a). Ces questions pourront être probablement résolues dans un proche avenir par l identification de gène(s) encodant d autre(s) récepteur(s) de l adénosine.
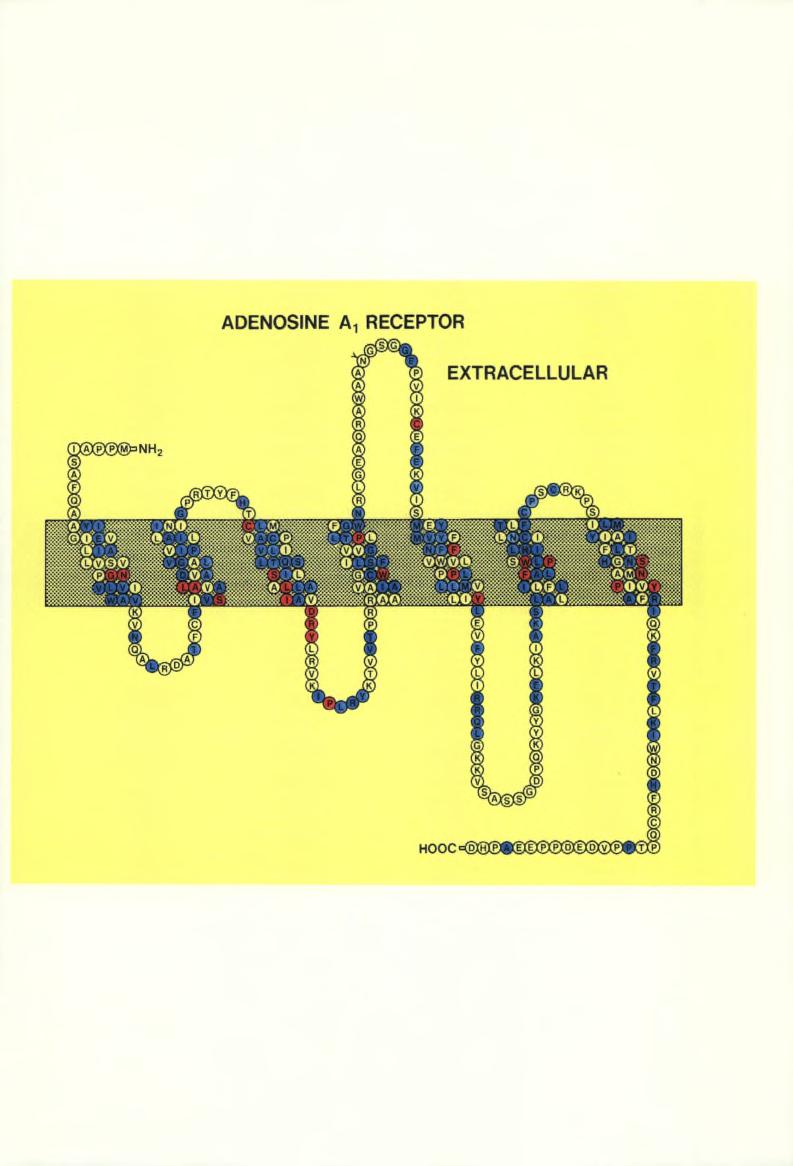
46

47 ADENOSINE A, RECEPTOR

48 -39- III.2.b Localisation du récepteur A2a de l adénosine dans les ganglions de la base et identification phénotypique des neurones qui l expriment (articles 1, 4, 5, 6) Comme décrit ci-dessus (cf. paragraphe III.2.a), la technique d hybridation in situ nous a permis d établir l expression sélective du récepteur A2a de l adénosine (RDC8) par les neurones de taille moyenne ("spiny neurons type I) des différents constituants du striatum chez le chien et le rat: striatum dorsal (caudé et putamen) et striatum ventral (accumbens et tubercule olfactif) (article 1 et fig. III. 1). Dans ces espèces, étudiées à l age adulte, aucune hétérogénéité n est observée. Ceci suggère une expression identique de ce récepteur dans les striosomes et la matrice extrastriosomale. Par contre, de très légers gradients dorso-ventral et latéro-médian sont observés, le striatum ventral et la région médiane du striatum dorsal étant en effet moins intensément marqués (fig. III. 1). Nous avons également discuté l absence de signal dans toute autre région cérébrale alors que des données pharmacologiques et physiologiques indiquent la présence d une activité de type A2 dans diverses régions extrastriatales. Nous avons présenté l existence d un autre sous-type de récepteur A2, le A2b, et/ou les limites de détection de nos techniques comme explications plausibles de ce fait (cf. paragraphe III.2.a). L hypothèse d une fonction importante du récepteur A2 de l adénosine dans la physiologie et la physiopathologie des ganglions de la base déjà suggérée par des données comportementales, se trouve renforcée par la démonstration de l expression sélective de ce récepteur dans le striatum chez deux espèces de mammifères. La compréhension des mécanismes pathogéniques des maladies dégénératives des ganglions de la base et des mécanismes d adaptation par neuroplasticité, requière naturellement l exploration de modèles expérimentaux (cf. ci-dessous). Néanmoins, les différences d espèces observées dans de nombreuses situations impliquent une grande prudence dans toute extrapolation d observations animales à l homme et nécessitent, quand cela est possible, une confirmation chez cette espèce. Ainsi, nous avons étudié par hybridation in situ la distribution des cellules exprimant le récepteur A2 dans le cerveau humain normal (article 4). Ceci devrait permettre ultérieurement de comparer les distributions de l ARN messager encodant ce récepteur dans le cerveau normal avec celles observées dans le cerveau de patients souffrant de pathologies telles que les maladies de Huntington et de Parkinson. Le séquençage partiel du récepteur A2 humain (Libert et al., 1989) (équivalent du RDC8 canin) nous a permis de construire une sonde oligonucléotidique totalement complémentaire de l ARN messager encodant celui-ci. Chez l homme, comme chez le chien et le rat, les différents éléments du striatum (caudé, putamen et accumbens) sont les seules structures cérébrales positives (cf. article 4, fig. 1). De même, les cellules striatales exprimant le récepteur A2 sont également exclusivement des neurones de taille moyenne et non des cellules gliales ou de grands neurones. Enfin, aucune différence d expression n est notée entre les striosomes et la matrice extrastriosomale alors qu un léger gradient dorso-ventral est observé. Cette expression sélective du récepteur A2 de l adénosine dans le striatum observée de manière identique chez trois espèces y compris l homme, nous autorise à utiliser le rat comme modèle expérimental. Dans le chapitre d introduction (cf. paragraphes II.2.a. et H.3.), nous avons insisté sur le fait qu il existe plusieurs classes morphologiques de neurones striataux et que, de plus, au sein d une même classe, des sous-populations neuronales peuvent être reconnues sur base de leur phénotype neurochimique. L identification phénotypique précise
 adjacentes.")
49 -40- des neurones exprimant le récepteur A2 est donc indispensable pour mieux comprendre la fonction du système adénosinergique dans la physiologie des ganglions de la base. Pour ce faire, nous avons utilisé la technique d hybridation in situ sur coupes fines (4-5 /im) adjacentes. Ceci permet de reconnaître de manière convaincante les mêmes neurones sur ces coupes adjacentes grâce à la sélection de champs caractérisés par un point de repère particulier, vaisseau ou neurone de grande taille. Les co-expressions potentielles du récepteur A2 et de trois éléments neurochimiques majeurs du striatum, ENK, SP et choline acétyltransférase (ChAT) caractéristiques de trois populations neuronales différentes sont analysées (article 5). Ceci démontre qu une très large majorité (92%) des neurones enképhalinergiques expriment le récepteur A2 de l adénosine (cf. fig. 2, article 5). Par contre, aucun neurone cholinergique ou à SP ne démontre d expression de ce récepteur détectable par nos techniques (cf. fig. 2, article 5). L importance des récepteurs dopaminergiques et principalement des récepteurs DI a et D2 dans la régulation de la fonction striatale, justifie également l analyse de leur co-expression éventuelle avec le récepteur A2 (article 6). Dans cette étude, nous n avons pas différencié les deux formes de récepteur D2 obtenues par épissage alternatif, D2a et D2b. Une co-expression quasi absolue des récepteurs A2 et D2 est observée alors qu elle est quasi nulle avec le récepteur DI (cf. fig. 1, article 6). Cette ségrégation neuronale de l expression du récepteur A2 implique donc que sa(es) fonction(s) s exercerai(en)t uniquement dans un des deux circuits striato-pallido-thalamique: la boucle inhibitrice (cf. paragraphe II.5.). En effet, il a été proposé que la sous-population neuronale striatale co-exprimant l ENK et le récepteur D2 (Le Moine et al., 1990), projette vers le globus pallidus (Gerfen et al., 1990) par opposition aux neurones co-exprimant la SP et le récepteur DI (Le Moine et al., 1991) qui innervent la substance noire (Gerfen et al., 1990). Pour confirmer définitivement le site de projection des neurones striataux exprimant le récepteur A2 de l adénosine, les neurones striato-nigraux sont marqués par traçage rétrograde après injection de fluorogold dans la substance noire et caractérisés neurochimiquement par hybridation in situ (article 6). La grande majorité des neurones striataux marqués par le fluorogold injecté dans la substance noire expriment comme attendu la SP et le récepteur DI et très rarement l ENK, le récepteur D2 ou le récepteur A2. Par contre, une large majorité des neurones exprimant l ENK, le récepteur D2 ou le récepteur A2 ne sont pas marqués par le fluorogold et considérés comme striato-pallidaux (fig. 1, article 6) (Gerfen et al., 1990). Ces différents éléments démontrent l expression cellulaire sélective du récepteur A2 de l adénosine par les neurones striato-pallidaux qui expriment également l ENK et le récepteur dopaminergique D2. Ils constituent un substratum plausible des interactions entre récepteurs A2 et D2 suggérées par certaines observations comportementales (Jarvis and Williams, 1987; Ferre et al., 1991a). III. 2.C Rôle(s) du(es) récepteur(s) A2 de l adénosine dans la régulation de l expression génétique dans le striatum (article 6). La voie dopaminergique nigro-striée inhibe l activité des neurones GABA-ENK striatopallidaux par l activation du récepteur D2 et stimule celle des neurones GABA-SP striatonigraux par l activation du récepteur Dl. Ce contrôle par la dopamine inclut la régulation de l expression génétique de différents constituants neurochimiques de ces neurones (Young et al., 1986; Gerfen et al., 1990; pour revue voir Graybiel, 1990). Ainsi, la destruction de la
. Ces observations démontrent que la dopamine inhibe toniquement la synthèse d ENK et stimule celle de SP.")
50 -41- voie dopaminergique nigro-striée induit une augmentation du contenu cellulaire en ARN messager encodant la proenk et le récepteur D2 et une diminution de celui des ARN messagers encodant la protachykinine et le récepteur DI (Young et al., 1986; Gerfen et al., 1990). Ces observations démontrent que la dopamine inhibe toniquement la synthèse d ENK et stimule celle de SP. De plus, un traitement avec un agoniste D2 sélectif, subséquent à la lésion nigro-striatale, rétablit une expression presque normale d ENK sans effet sur l expression diminuée de SP (Gerfen et al., 1990). Inversement, un traitement subséquent avec un agoniste DI sélectif rétablit une expression presque normale de SP sans effet sur l expression augmentée d ENK (Gerfen et al., 1990). Ces observations sont totalement cohérentes avec l expression sélective des récepteurs DI et D2 respectivement dans les neurones à SP et ENK (Le Moine et al., 1990, 1991). La co-expression des récepteurs D2 et A2 par les neurones enképhalinergiques, et le fait que ces deux récepteurs agissent respectivement en inhibant (Stoof and Kebabian, 1981) et en stimulant (Van Calker et al., 1979) l activité de l adénylate cyclase, nous suggère la possibilité d une intervention du récepteur A2 dans la régulation de l expression génétique dans le striatum et ce dans un sens opposé à celui du récepteur D2. Pour tester cette hypothèse, des rats porteurs d une lésion unilatérale de la substance noire supérieure à 90% et induite par l injection unilatérale de 6-hydrodxydopamine (6- OHDA), sont traités subséquemment par un antagoniste des récepteurs de l adénosine, la caféine (75 mg/kg/j). Si l hypothèse se vérifie nous devons observé, comme après un traitement avec un agoniste D2 sélectif, un amoindrissement de l augmentation d expression d ENK sans effet sur l expression diminuée de SP. Une étude autoradiographique quantitative (cf. fig. 2, 3 article 6) des taux d ARN messagers encodant l ENK et la SP confirme que chez les animaux lésés et ensuite traités comme contrôle avec 0,9% NaCl, l expression d ENK est augmentée dans le striatum ipsilatéral à la lésion avec un rapport des densités optiques du côté lésé versus côté intact (L/I) de 2.33 alors que l expression de SP est diminuée avec un rapport L/I de Chez les rats traités subséquemment avec de la caféine, on observe une chute dramatique du rapport L/l (1.45) pour l ENK. Néanmoins, si l on tient compte des valeurs absolues de densité optique, on constate que ce résultat est dû non seulement à une diminution prédite d expression d ENK dans le striatum lésé, mais aussi à une augmentation d expression dans le striatum controlatéral intact. De plus, contrairement à toute attente, pour la SP, le rapport L/I augmente sensiblement (0.78). Cette augmentation est due uniquement à une diminution de l expression de SP dans le striatum controlatéral intact et non à une normalisation de l expression de SP du côté lésé. En effet, dans le striatum ipsilatéral à la lésion, l expression de SP n est pas, en valeur absolue, significativement différente de celle observée dans le striatum ipsilatéral des rats contrôles traités avec 0,9% NaCl, où elle est diminuée par rapport au coté intact. En résumé, dans le striatum dépiété en dopamine, nous constatons après traitement par un antagoniste A2, une diminution de l expression d ENK sans modification d expression de SP comme cela est décrit après traitement par un agoniste D2. Ceci confirme notre hypothèse d un rôle du récepteur A2 dans la régulation d expression d ENK dans les neurones striato-pallidaux et démontre l existence d une balance D2/A2 dans la régulation de l activité de ces neurones. Néanmoins, un autre effet du traitement par la caféine a été observé dans le striatum non dépiété en dopamine. Pour comprendre cet effet, quatre groupes de rats normaux sont soumis aux traitements suivants pendant 15 jours (article 6): 1. véhicule (0,9% NaCl), 2. caféine (75 mg/kg/j), 3. caféine (75 mg/kg/j) + NECA (0,25 mg/kg/j), un agoniste equipotent des récepteurs A2 et Al, 4. caféine (75 mg/kg/j) + CHA (0,5 mg/kg/j), un

51 agoniste sélectif du récepteur Al La caféine induit une augmentation de 67% de l expression d ENK. Le NECA bloque de manière statistiquement significative cet effet de la caféine en ramenant l expression d ENK au niveau contrôle. Le CHA n a aucun effet sur les modifications d expression induites par la caféine et il existe une différence significative entre les effets des traitements caféine + NECA et caféine + CHA. L implication d un récepteur de l adénosine dans cette action de la caféine est donc confirmée par le traitement combiné avec un agoniste de ces récepteurs. Les faits observés plaident pour l implication d un récepteur A2 et non d un récepteur Al. En effet, bien que le NECA soit un agoniste équipotent des récepteurs Al et A2, il est dix fois moins puissant que le CHA vis-à-vis du récepteur Al et de plus il a été administré à une dose deux fois moins importante. Il y aurait donc un facteur 20 en faveur du CHA pour l activation du récepteur Al (Bruns et al., 1986). Contrairement à cela et en faveur d un récepteur A2, seul le NECA démontre un effet significatif alors que le CHA est lui sans aucun effet. Quels seraient le site d action et le type de récepteur impliqués? La caféine n exerce cet effet que si la voie dopaminergique nigro-striée est intacte (cf. ci-dessus), suggérant que l intégrité des terminaisons axonales dopaminergiques est requise et que la dopamine elle- même est impliquée. Par ailleurs, le tableau observé, augmentation de l expression d ENK et diminution d expression de SP, est caractéristique des situations de déficit d activité dopaminergique, déplétions en dopamine ou blocages des récepteurs dopaminergiques post- synaptiques (Young et al., 1986; Gerfen et al., 1990; pour revue voir Graybiel, 1990). Plusieurs auteurs ont décrit un récepteur A2 situé pré-synaptiquement sur les terminaisons nigro-striatales, dont l activation stimule la synthèse et la libération de dopamine (Onali et al., 1988; Zetterstrôm and Fillenz, 1990). En agissant comme antagoniste A2 de ce récepteur, la caféine pourrait donc provoquer une déplétion en dopamine dans le striatum. L absence de signal détectable dans la substance noire en hybridation in situ à l aide des sondes spécifiques du récepteur A2 striatal (A2a) (cf. paragraphe III.2.b), l absence dans cette même substance noire de liaison d un agoniste A2a spécifique radiomarqué, [^H]CGS21680 (Jarvis and Williams, 1989; Parkinson and Fredholm, 1990), et enfin l incapacité de ce même ligand de stimuler la libération de dopamine (Lupica et al., 1990) suggèrent très fortement que le récepteur impliqué soit le sous-type A2b. Cette hypothèse ne pourra être démontrée définitivement que par l identification du gène encodant ce récepteur. III. 2.d Modifications d expression du récepteur A2a de l adénosine dans le striatum au cours du développement (article 7) L importance fonctionnelle du récepteur A2a de l adénosine à la lumière des éléments rapportés ci-dessus et des études comportementales antérieures imposent l étude de son expression au cours du développement du système striatal. En effet, ce récepteur pourrait avoir une fonction dans la mise en place du contrôle de la fonction motrice au cours du développement. Ainsi, un traitement maternel pré-natal et/ou post-natal par un antagoniste du récepteur A2 telle que la caféine, induit des anomalies de l activité locomotrice chez les ratons issus de ces portées (Guillet, 1990). Les mécanismes de détermination et d établissement de la compartimentalisation du striatum constituent le point d intérêt principal de l étude du développement de cette structure. L identité striosomale ou matricielle des neurones striataux est-elle déterminée génétiquement (facteurs intrinsèques) ou nécessite-t-elle des influences extrinsèques telles
 Bien que cette question ne soit pas encore définitivement résolue à l heure actuelle, de multiples arguments plaident pour la première hypothèse.")
52 -43- que les afférences dopaminergiques pour se développer? (Fishell and Van der Kooy, 1987) Bien que cette question ne soit pas encore définitivement résolue à l heure actuelle, de multiples arguments plaident pour la première hypothèse. De nombreux éléments neurochimiques du striatum démontrent une distribution caractéristique consistant en un enrichissement striosomal au cours du développement et une distribution homogène ou un enrichissement matriciel à l âge adulte (Lowenstein et al., 1989, Murrin and Zeng, 1989; Nastuk and Graybiel, 1989). Nous avons étudié les distributions striatales de l ARN messager encodant le récepteur A2a de l adénosine par hybridation in situ et des sites de liaison d un ligand A2 spécifique, [3h]CGS21680, par autoradiographie au cours du développement pré- et post-natal du rat, post-natal du chien et chez un foetus humain de 18 semaines de gestation (article 7). Ses distributions ont été comparées avec celle des sites de liaison d un antagoniste sélectif du récepteur dopamine Dl, [^H]SCH23390, décrits comme marqueurs des striosomes au cours du développement (Murrin and Zheng, 1989). La distribution des récepteurs opioïdes de type mu qui sont les marqueurs les plus adéquats des striosomes chez le rat (Herkenham and Pert, 1981, Caboche et al., 1991), a été également déterminée chez cette espèce à l aide du ligand spécifique [^HJDAGO. Chez le rat, l ARN messager encodant le récepteur A2 est détecté dans le striatum latéro-ventral et également légèrement dans la couche germinative à E19. Cependant, nous ne pouvons pas exclure une expression plus précoce comme cela est décrit pour le récepteur dopaminergique D2 (Guennoun and Bloch, 1991). En effet, la sonde oligonucléotidique que nous utilisons est complémentaire à la séquence canine et n est probablement pas totalement complémentaire de la séquence de rat qui n est pas encore connue à ce jour. Une sensibilité moindre et un rapport bruit de fond/signal plus élevé sont possibles. Dans la période postnatale précoce, PI à PI3 chez le rat et P2 et P9 chez le chien, les distributions striatales des récepteurs A2 et de leur ARN messager se présentent de manière hétérogène avec un enrichissement dans des zones circulaires ou elliptiques (fig. 2,4 article 7). Ces zones enrichies en récepteurs A2 et leur ARN messager correspondent sur des coupes adjacentes aux striosomes enrichis en récepteurs dopaminergiques Dl ou opioïdes de type mu (fig. 2,4 article 7). Le striatum de foetus humain de 18 semaines démontre également une hétérogénéité de marquage avec des zones enrichies en récepteurs A2 et leur ARN messager qui correspondent sur des coupes adjacentes avec celles enrichies en récepteurs dopaminergiques Dl (fig. 5 article 7). L hybridation in situ permet d examiner l expression de ce récepteur à l échelle cellulaire. Ceci indique que l enrichissement en récepteur A2 de l adénosine dans les striosomes au cours du développement est lié à une plus grande densité de neurones exprimant ce récepteur dans ce compartiment et également à une expression individuelle plus élevée des neurones striosomiaux comparée à celle des neurones de la matrice (fig. 5-7 article 7). Cette observation démontre que les différences de capacité de liaison de certains récepteurs entre striosomes et matrice sont au moins partiellement liés à une différence du niveau d expression génétique de ces récepteurs. III. 3. Conclusions et perspectives L action neuromodulatrice de l adénosine est médiée par sa liaison à des récepteurs membranaires appartenant à la famille des récepteurs couplés aux protéines G. L action inhibitrice de l adénosine via son récepteur Al constitue la base d un effet neuroprotecteur dans des conditions physiopathologiques telles que l ischémie ou l épilepsie (Dragunow and

53 -44- Faull, 1988; Chin, 1989; Petersen, 1991; Von Lubitz et al., 1988). Par sa localisation préférentielle dans le système des ganglions de la base, le récepteur A2 est lui impliqué dans le contrôle du mouvement ou de l activité locomotrice (Fuxe and Ungerstedt, 1974; Snyder et al., 1981; Green et al., 1982; Jarvis and Williams, 1987; Durcan and Morgan, 1989; Ferre et al., 1991a). De plus, ces récepteurs sont la cible de molécules comme la caféine, la théophyline et la théobromine (Bruns et al., 1986) présentes dans des boissons de consommation courante comme le café, le thé, le cacao ou le cola. Nous avons, dans ce travail, identifié et caractérisé deux récepteurs orphelins, RDC7 et RDC8, de la famille des récepteurs couplés aux protéines G comme étant les récepteurs Al et A2 de l adénosine. La combinaison de techniques différentes a permis de poser cette hypothèse sur base d un faisceau d arguments convergents et ensuite de la confirmer. En effet, de même qu un récepteur orphelin a été identifié comme étant le récepteur aux cannabinoïdes par l examen de sa localisation (Matsuda et al., 1990), notre étude des distributions anatomiques précises de RDC7 et RDC8 par hybridation in situ s est révélée prépondérante. L identification des gènes de ces deux récepteurs ouvre la voie d une nouvelle pharmacologie des récepteurs de l adénosine car tant au niveau du système nerveux central qu au niveau périphérique (cardiaque, pulmonaire,...) le développement d agonistes et d antagonistes Al et A2 sélectifs s avère indispensable pour envisager leur utilisation clinique. Par ailleurs, cette identification devrait permettre dans un proche avenir le clonage d autres sous-types de récepteurs de l adénosine dont l existence est proposée (A2b, A3, sous-types de Al) et donc d affiner notre compréhension des mécanismes d action de l adénosine. Dans ce travail nous nous sommes également attachés à mieux comprendre l implication du récepteur A2 de l adénosine dans la physiologie des ganglions de la base. Alors que les éléments existants à ce sujet proviennent essentiellement de données obtenues à l aide de modèles comportementaux (Fuxe and Ungerstedt, 1974; Snyder et al., 1981; Green et al., 1982; Jarvis and Williams, 1987; Durcan and Morgan, 1989; Ferre et al., 1991a), notre contribution repose sur l utilisation de techniques de neuroanatomie et de neurochimie et de modèles expérimentaux de lésions et traitements pharmacologiques. Nous avons établi l expression sélective de ce récepteur par les neurones de taille moyenne ("spiny neurons type I") du striatum chez trois espèces de mammifères: le rat, le chien et l homme. De plus, nous avons démontré que parmi ces neurones, seule la souspopulation exprimant l ENK et le récepteur dopaminergique D2 et projettant vers le globus pallidus (globus pallidus externe des primates) exprime le récepteur A2 de l adénosine. Cette observation constitue très probablement le substratum anatomique de l interaction A2/D2 démontrée par les études comportementales. Nous avons également étudié les modifications de l expression génétique des neuropeptides du striatum chez des animaux dépiétés ou non en dopamine par une lésion de la substance noire et traités par un antagoniste des récepteurs de l adénosine. Ceci a démontré que l adénosine est impliquée dans la régulation de cette expression génétique via des récepteurs A2 par deux mécanismes d action différents. Le premier est lié directement à l activation des récepteurs A2a post-synaptiques présents sur les neurones striato-pallidaux exprimant l ENK et le récepteur D2 et évoque un effet anti-d2 qui permet de proposer l existence d une balance régulatrice D2/A2 dans ces neurones.

54 -45- La localisation sélective de ce récepteur A2 dans les neurones striato-pallidaux et le contrôle positif qu il exerce sur leur expression génétique signifie qu il est impliqué essentiellement dans le fonctionnement de la boucle inhibitrice "striatum ->globus pallidus externe ->noyau sous-thalamique ->globus pallidus interne et substance noire pars reticulata ->thalamus". Il faut rappeler que cette boucle est hypoactive dans les désordres hyperkinétiques, hémiballisme et maladie de Huntington, alors qu elle est hyperactive dans les désordres hypokinétiques de type parkinsonien (Albin et al., 1989) (cf. paragraphe II.6). Outre l apport en dopamine ou agonistes dopaminergiques, la diminution de l hyperactivité de cette boucle par d autres moyens doit améliorer le parkinsonisme. Ceci a, en effet, été observé après lésion du noyau sous-thalamique dans un modèle animal de parkinsonisme (Bergman et al., 1990), Nos résultats démontrent que le blocage du récepteur A2a par des antagonistes sélectifs diminue également l activité de cette boucle. Le développement de telles molécules devrait donc permettre d accroître l arsenal thérapeutique de la maladie de Parkinson, sachant que tout patient parkinsonien traité par la L-DOPA développe tôt ou tard des fluctuations de ses performances motrices et/ou des mouvements involontaires. Le second mécanisme par lequel l adénosine est impliquée dans la régulation de l expression génétique du striatum, dépend de la présence des fibres dopaminergiques nigrostriées et consiste probablement en l activation d un récepteur pré-synaptique A2b stimulant la libération de dopamine. Bien que nos données suggèrent l existence de ce mécanisme, celui-ci ne pourra être définitivement caractérisé que par la démonstration par clonage de l existence de ce récepteur A2b et de son expression par les neurones dopaminergiques de la substance noire et/ou le développement et l utilisation de ligands A2b spécifiques. Ce travail ouvre la voie à l étude des mécanismes d interaction entre les récepteurs D2 de la dopamine et A2a de l adénosine à l échelle cellulaire. La démonstration d une diminution de l affinité du récepteur D2 résultant d une activation du récepteur A2 suggère également l existence d une interaction intramembranaire (Ferre et al., 1991b). De plus, les canaux ioniques auxquels ces récepteurs pourraient être couplés sont, comme les seconds messagers tels que l AMP cyclique, des cibles communes potentielles d une interaction entre ces deux récepteurs. L étude, par combinaison de techniques biochimiques, pharmacologiques et électrophysiologiques, de cellules striatales ou de lignées cellulaires manipulées génétiquement devrait permettre une meilleure compréhension de ces mécanismes. L ontogenèse du récepteur A2a dans le striatum a démontré des modifications de sa distribution au cours du développement. Le compartiment striosomal est transitoirement enrichi en récepteur A2a par rapport à la matrice. Le traitement par de la caféine pendant la gestation et la lactation, à des doses compatibles avec une consommation moyenne de café chez l homme, provoque des effets délétères sur l activité locomotrice des jeunes animaux issus de ces portées (Guillet, 1990). La connaissance du développement normal et l étude des modifications d expression du récepteur A2 ainsi que des neuropeptides et des récepteurs dopaminergiques dans le striatum à différents stades du développement post-natal chez ces jeunes animaux devraient permettre d élucider les mécanismes menant à ces comportements anormaux.

55 -46- IV. DEUXIEME PARTIE: LOCALISATION ET FONCTION DE LA CHOLECYSTOKININE ET SES RECEPTEURS Localisation de TARN messager encodant la cholécystokinine dans le cerveau de rat au niveau du mésencéphale. Image en pseudo-couleur, le rouge correspondant aux régions les plus riches.
56 -47- Les données discutées dans le présent chapitre sont exposées dans les articles suivants: 8. SCHIFFMANN, S.N., MAILLEUX, P., PRZEDBORSKI, S., HALLEUX, P., LOTSTRA, F. and VANDERHAEGHEN, J-J. : Cholecystokinin distribution in the human striatum and related subcortical structures, Neurochem. Int., 14, , SCHIFFMANN, S.N., and VANDERHAEGHEN J.-J.: Distribution of cells containing mrna encoding cholecystokinin in the rat central nervous System. J. Comp. Neurol., 304, , SCHALLING, M., FRIBERG, K., BIRD, E., SCHIFFMANN, S., MAILLEUX, P., VANDERHAEGHEN, J-J. and HÔKFELT, T. : Presence of cholecystokinin mrna in dopamine cells in the ventral mesencephalon of a human with schizophrenia. Acta Physiol. Scand., 137, , SCHALLING, M., FRIBERG, K., SEROOGY, K., RIEDERER, P., BIRD, E., SCHIFFMANN, S.N., MAILLEUX, P., VANDERHAEGHEN, J.-J., KUGA, S., GOLDSTEIN, M., KITAHAMA, K., LUPPI, P.H., JOUVET, M. and HÔKFELT, T.: Analysis of expression of cholecystokinin in dopamine cells in the ventral mesencephalon of several species and in humans with schizophrenia. Proc. Ntl. Acad. Sci. USA, 87, , SCHIFFMANN S.N. and VANDERHAEGHEN J.-J.: Dopaminergic Régulation of cholecystokinin mrna expression in the rat striatum. An in situ hybridization histochemistry study. Submitted, SCHIFFMANN, S.N., GOLDMAN, S., HEYMAN, P., DE VUYST, E., DE ROY, G. and VANDERHAEGHEN J.-J.: Ontogeny of cholecystokinin receptors in the human striatum. Submitted, 1992.

57 -48- IV.1. Introduction et Objectifs En 1975, Vanderhaeghen et al. décrivent la présence d un peptide reconnu par un anticorps dirigé contre la portion commune de la gastrine et de la cholécystokinine dans le cerveau de nombreuses espèces animales dont l homme. La majeure partie de cette immunoréactivité est constituée par l octapeptide C-terminal de la cholécystokinine (CCK-8) dont la forme sulfatée (CCK-8S) est biologiquement active (Dockray et al., 1976; Robberecht et al., 1978). Dans les années qui ont suivi cette observation, la présence dans le système nerveux d un nombre sans cesse croissant de peptides biologiquement actifs dans les tissus périphériques a été démontrée, (pour revue voir Bjôrklund and Hôkfelt, 1985) Les premières études rapportant la localisation cérébrale de la cholécystokinine par des techniques de radioimmunoessai ont démontré une distribution étendue chez le rat et l homme (Vanderhaeghen et al., 1975; Rehfeld, 1978; Beinfeld et al., 1981; Emson et al., 1982; Vanderhaeghen, 1985). Néanmoins, les données plus précises obtenues par les techniques immunohistochimiques ont permis de nuancer cette notion et de localiser ce neuropeptide dans des populations et des sous-populations neuronales bien spécifiques (Innis et al., 1979; Hôkfelt et al., 1980a,b; Vanderhaeghen et al., 1980a,b; Vanderhaeghen, 1985; Hôkfelt et al., 1988). La fonction de neuromodulateur des neuropeptides tels que la cholécystokinine a été proposée en raison de leur synthèse de novo dans le cerveau, leur présence dans les terminaisons axonales, leur libération synaptique par des mécanismes semblables à ceux décrits pour la libération des neurotransmetteurs classiques, la démonstration de récepteurs post-synaptiques spécifiques et la capacité par activation de ces récepteurs de modifier le status électrique des neurones post-synaptiques (ex: excitation des neurones hippocampiques de la corne d Ammon). Deux récepteurs spécifiques de la cholécystokinine ont été décrits: le récepteur CCK-B (B pour Brain) est prédominant dans le système nerveux central alors que le récepteur CCK-A l est dans les tissus périphériques bien qu il soit également présent dans diverses régions cérébrales (Hill et al., 1987). Outre le cortex cérébral, le striatum est une des régions cérébrales dont la concentration en cholécystokinine est la plus élevée. De nombreuses études immunohistochimiques se sont attachées à définir la distribution précise de ce neuropeptide dans les différentes structures des ganglions de la base chez le rat. Dans le striatum de rat, l immunoréactivité détectée par un anticorps dirigé contre la cholécystokinine (cholécystokinine-ir) n est pas distribuée de manière homogène. Elle est plus abondante dans la partie ventrale et plus précisément dans l accumbens médian (Hôkfelt et al., 1980a,b; Vanderhaeghen et al., 1980a,b). Dans le striatum dorsal, quatre systèmes différents de fibres ont été décrits (Hôkfelt et al., 1988): 1. des fibres périventriculaires fortement marquées dans le striatum caudal, 2. des fines fibres punctiformes dans la partie médiane du striatum, 3. des fines fibres punctiformes faiblement marquées dans l ensemble du striatum et 4. des îlots ("patches") de fibres fortement marquées principalement dans le striatum médian. L origine des fibres à cholécystokinine présentes dans le striatum a constitué un sujet de débat pendant de longues années (Hôkfelt et al., 1980a,b; Vanderhaeghen et al., 1980a; Meyer et al., 1982; Gilles et al., 1983; Hôkfelt et al., 1988; Meyer et al., 1988). Le problème principal rencontré dans les études immunohistochimiques est la visualisation des corps

58 -49- cellulaires neuronaux positifs. En effet dans la plupart des cas, y compris pour la cholécystokinine dans de nombreuses régions, les taux de neuropeptides dans les somas sont inférieurs au seuil de détection de ces techniques probablement car le peptide est quasi complètement exporté dans les prolongements dendritiques et axonaux. Pour pallier à cet inconvénient, l injection de colchicine, poison des microtubules bloquant le flux axonal, permet de concentrer les neuropeptides dans certains corps cellulaires, les rendant ainsi détectables par immunohistochimie. La présence de neurones arborant une cholécystokinine-ir dans le striatum lui-même a été très discutée. Certaines études rapportent la présence d un petit nombre de tels neurones (Hôkfelt et al., 1988) alors que d autres n en détectent pas (Vanderhaeghen et al., 1980a; Gilles et al., 1983). Néanmoins, même les auteurs rapportant la présence de ces neurones insistent sur le fait que ceux-ci participent peu à l élaboration des différents systèmes de fibres cholécystokininergiques du striatum puisqu une lésion sélective des neurones intrinsèques du striatum par l acide iboténique (cf. paragraphe II.6.) ne modifie pas la concentration striatale de cholécystokinine-ir (Gilles et al., 1983). La diminution des concentrations de cholécystokinine-ir au niveau du globus pallidus et de la substance noire dans la maladie de Huntington (Emson et al., 1980) a néanmoins suggéré la possibilité d une synthèse striatale intrinsèque de cholécystokinine. L apport des techniques d hybridation in situ et de Northern blotting d ARN a permis de progresser rapidement dans la résolution de ce problème et sera discuté dans le présent travail (cf. paragraphes IV.2.b et IV.2.c). La source de la cholécystokinine striatale est donc principalement extrinsèque. Deux sources majeures ont été proposées et démontrées par combinaison de techniques immunohistochimiques et de lésions expérimentales d afférences striatales. Dans ce cas également, l hybridation in situ a apporté de nouveaux développements que nous discutons dans le présent travail (cf. paragraphe IV.2.b). La cholécystokinine et la dopamine coexistent dans les neurones dopaminergiques de l aire tegmentale ventrale ou A10 (Hôkfelt et al., 1980a, 1980b; Vanderhaeghen et al., 1980a, Seroogy et al., 1987, 1989a). Cette coexistence rend compte de l abondance de l innervation cholécystokininergique du striatum ventral. La présence de cholécystokinine dans des fibres dopaminergiques provenant de la substance noire pars compacta ou A9 est également fortement suggérée par la disparition des systèmes de fibres 1 et 2 dans le striatum dorsal après une lésion de cette substance noire par la 6-hydroxydopamine (cf. paragraphe II.6.) (Hôkfelt et al., 1988). La seconde source de fibres à cholécystokinine dans le striatum est constituée par des structures telles que le claustrum, le cortex piriforme et l amygdale. Ceci a été démontré par la diminution des concentrations striatales de cholécystokinine après des lésions ayant isolé ces structures du striatum (Meyer et al., 1982). L abondance de la cholécystokinine dans la partie ventrale du striatum et l aire tegmentale ventrale suggère sa participation dans la boucle dite limbique (cf. paragraphes II.2.b. et II.2.C.) du système des ganglions de la base et donc dans les comportements d initiation des mouvements en réponse aux stimuli émotionnels. Cette localisation et sa relation avec le système dopaminergique du système ventral a poussé certains auteurs a
59 -50- envisagé la participation de la cholécystokinine dans la pathogénie de la schizophrénie (pour revue voir Wang et al., 1984). Par ailleurs, sa présence significative dans le striatum dorsal et la substance noire pars compacta permet d envisager également un rôle de ce peptide dans le contrôle de la motricité. A l appui de cette hypothèse, il faut noter que la cholécystokinine modifie la libération de dopamine dans le noyau caudé (Markstein and Hôkfelt, 1984) comme dans l accumbens (Voigt and Wang, 1984). Inversement, la libération de cholécystokinine est modulée par la dopamine (Meyer and Krauss, 1983). Dans cette seconde partie du mémoire, nous avons étudié la distribution de la cholécystokinine dans les ganglions de la base chez l homme normal avec une référence particulière au striatum ventral et ses extensions dans la substance innominée (paragraphe IV.2.a, article 8). L étude de la compartimentalisation des récepteurs de la cholécystokinine dans le striatum humain à Page adulte. et au cours du développement a été menée par autoradiographie (paragraphe IV.2.d, article 13). La distribution des cellules exprimant l ARN messager de la cholécystokinine dans les ganglions de la base ainsi que dans les régions cérébrales qui y projettent est analysée par hybridation in situ chez le rat (paragraphe IV.2.b, article 9). Ces résultats sont complétés par la caractérisation des neurones exprimant l ARN messager de la cholécystokinine dans le mésencéphale ventral chez l homme normal ou souffrant de schizophrénie et leur relation avec le système dopaminergique (paragraphe IV.2.b, articles 10, 11). Enfin, la possibilité d une capacité d expression de l ARN messager encodant la cholécystokinine par les neurones intrinsèques du striatum a été investiguée par hybridation in situ après modifications expérimentales de la neurotransmission dopaminergique (paragraphe IV.2.C, article 12)

60 -51- IV. 2. RESULTATS ET DISCUSSION IV.2.a Localisation immunohistochimique de la cholécystokinine dans les ganglions de la base chez l homme (article 8) Un très grand nombre de fines fibres positives est présent dans le striatum humain, avec une densité plus élevée dans sa partie ventrale, noyau accumbens, que dans sa portion dorsale. Ces fibres sont distribuées de manière hétérogène avec des îlots circulaires ou elliptiques ("patches") de plus haute densité observés principalement dans le noyau caudé (fig. 2A article 9). Sur des coupes adjacentes, ces îlots sont en parfaite correspondance avec les striosomes dont les limites sont enrichies en ENK (fig. IV. 1) suggérant que les fibres à cholécystokinine sont plus abondantes dans le compartiment striosomal. La projection préférentielle vers les striosomes des fibres amygdalostriatales provenant du noyau basolatéral (Ragsdale and Graybiel, 1988), l abondance de neurones à cholécystokinine dans ce noyau, et le fait que chez le rat, les fibres cholécystokininergiques en îlots ne sont pas issues du mésencéphale (Hôkfelt et al., 1988) constituent autant d arguments en faveur de l origine amygdalienne de ces fibres cholécystokininergiques dans les striosomes. Fig. IV. 1: Localisation immunohistochimique de la cholécystokinine (a) et de la met-enképhaline (b) sur des coupes transversales adjacentes du striatum rostral. Les flèches indiquent la correspondance des striosomes dont les limites sont enrichies en fibres enképhalinergiques avec des zones riches en fibres à cholécystokinine. (C) noyau caudé, (P) putamen Aucun neurone positif n est détecté dans le striatum alors que, sur les mêmes coupes, de très nombreux neurones clj^olécystokininergiques sont présents dans le claustrum (fig. 3A-C article 9). De même, aucune structure positive, fibre ou neurone, n est détectée dans les divisions externe et interne du globus pallidus. Dans la substance innominée et le noyau du lit de la strie terminale, des faisceaux de fibres positives sont observés. Ces fibres ont un aspect caractéristique en fibres laineuses ou

61 -52- "woolly fibres" (cf. paragraphe II.2.a.) (fig. 3F,G article 9). Elles sont présentes rostralement dans les portions sus- et sous-commissurales du noyau du lit de la strie terminale, plus caudalement le faisceau s incurve latéro-ventralement pour pénétrer dans la substance innominée et finalement, différents îlots de ces fibres sont présents dans la partie caudale de cette même région (fig. 2B,C article 9). Un complément à ce travail démontre que de telles fibres sont également présentes dans la région du noyau central de l amygdale dans lequel l on trouve aussi des neurones positifs (fig. IV.2). De plus, sur des coupes adjacentes, tous ces îlots de fibres laineuses cholécystokininergiques correspondent très exactement à des îlots de fibres laineuses à ENK (fig. IV.3) et le noyau central de l amygdale contient également des neurones enképhalinergiques (fig. IV.2). Cette localisation en continuum du noyau du lit de la strie terminale au noyau central de l amygdale via la substance innominée suggère que ce système à cholécystokinine et ENK fait partie de l "extended amygdala" (Alheid and Heimer, 1988) (cf. paragraphe II.2.b.). Ceci va à l encontre de la notion de spécificité striato-pallidale, dorsale ou ventrale, de la structure en fibres laineuses (Haber and Eide, 1981; Haber and Nauta, 1983; Haber and Watson, 1985). L absence ou la rareté de neurones à cholécystokinine dans le striatum, du moins dans des conditions normales (cf. paragraphe IV.2.C), plaide également contre l origine striato-pallidale de ces fibres. m A V - i V > " ^, <ï MT ^ 1 à V - ^ * ; B» è. V. ^ U* V i.t' ^ ^,1* Fig. IV.2: Détection immunohistochimique de neurones contenant de la met-enképhaline (A) et de la cholécystokinine (B) dans le noyau central de l amygdale. Barres d échelle = 20 «* % % *

62 -53- Fig. IV.3: Localisation immunohistochimique de la met-enképhaline (A,C) et de la cholécystokinine (B,D) sur des coupes transversales adjacentes au niveau du noyau lenticulaire. Correspondance parfaite des îlots de fibres laineuses à cholécystokinine et à enképhaline dans la substance innominée et la région du noyau central amygdalien. Une zone de la substance innominée sous-lenticulaire en A et B est montrée à plus fort grossissement en C et D. Barres d échelle = 20 /im. (Ce) noyau central de l amygdale, (SI) substance innominée IV.2.b Localisation de l ARN messager encodant la cholécystokinine dans les ganglions de la base et les structures cérébrales qui y projettent avec référence particulière au mésencéphale ventral (articles 9, 10, 11) La technique d hybridation in situ a permis d apporter de nouvelles informations concernant la localisation de neurones qui synthétisent la cholécystokinine et donc sur les voies cérébrales susceptibles de l utiliser. Chez le rat adulte, aucun neurone positif n est observé dans le striatum (fig 1 article 9), caudé-putamen, accumbens et tubercule olfactif, confirmant ainsi qu il n existe peu voire pas

63 -54- de synthèse striatale intrinsèque de cholécystokinine dans des conditions normales. De même, aucun neurone positif n est observé dans le globus pallidus ou le pallidum ventral. L étude de la distribution de l ARN messager encodant la cholécystokinine démontre deux situations non prévues par les observations immunohistochimiques antérieures. D une part, un très grand nombre de neurones positifs sont présents dans des régions dépourvues ou pauvres en cholécystokinine-ir telles que le thalamus ou les colliculi inférieurs (cf. article 9). D autre part, la densité de neurones positifs est très supérieure à celle observée par immunohistochimie dans des régions riches en cholécystokinine-ir telles que le cortex cérébral, la formation hippocampique, le cortex piriforme, le claustrum et le noyau endopiriforme, certains noyaux amygdaliens (cf. article 9) et les motoneurones de la corne antérieure de la moelle épinière (Schiffmann et al., 1991) (cf. Thèse Annexe III.). Dans toutes ces régions, une très grande proportion des neurones expriment l ARN messager de la cholécystokinine. Dans le premier groupe, ces neurones sont faiblement à modérément marqués alors que dans le second groupe, outre une vaste majorité de neurones faiblement à modérément marqués, il existe une petite minorité de neurones fortement marqués (cf. article 10). La densité de ces derniers neurones est équivalente à celle des neurones visualisés par immunohistochimie suggérant que cette dernière technique ne détecterait essentiellement que les neurones à forte expression. Ainsi, dans le cortex cérébral, seulement 0.5 à 2% (Demeulemeester et al., 1985) des neurones, reconnus comme des interneurones, contiennent de la cholécystokinine-ir alors que plus de 50% des neurones expriment l ARN messager de la cholécystokinine parmi lesquels des neurones pyramidaux. Bien que plusieurs explications puissent être avancées, la plus plausible est une meilleure sensibilité de l hybridation in situ par rapport à l immunohistochimie surtout en ce qui concerne des neurones faiblement positifs qui ont de plus des projections longues comme les neurones pyramidaux du cortex et de l hippocampe, les neurones thalamiques ou les motoneurones. Parmi ces structures cérébrales, le cortex cérébral, la formation hippocampique, le claustrum, l amygdale et le thalamus projettent vers le striatum. L abondance des neurones exprimant l ARN messager de la cholécystokinine dans ces structures constitue donc un argument solide en faveur de leur apport en fibres cholécystokininergiques dans le striatum. Ceci avait déjà été proposé en ce qui concerne le claustrum, l amygdale et le cortex piriforme (Meyer et al., 1982). Par ailleurs, la combinaison de l hybridation in situ et de marquage rétrograde par injection de fluorogold a démontré que dans les noyaux thalamiques intralaminaires (centromédian, paracentral, centrolatéral) et dans le cortex cérébral, des neurones exprimant l ARN messager de la cholécystokinine projettent bien vers le striatum (Burgunder and Young, 1988, 1990). Les neurones du mésencéphale ventral constituent une autre source de cholécystokinine striatale (Hôkfelt et al.., 1980a, 1980b; Vanderhaeghen et al., 1980a, Seroogy et al., 1987, 1989a). Dans ce cas également, l hybridation in situ démontre que chez le rat, une grande proportion de ces neurones expriment l ARN messager de la cholécystokinine y compris dans la substance noire pars compacta ou A9 (fig 1,7 article 9). De plus, par la combinaison des
 ont démontré définitivement que ces neurones exprimant l ARN messager de la cholécystokinine sont bien des neurones dopaminergiques du mésencéphale ventral.")
64 -55- techniques d immunohistochimie et d hybridation in situ, Seroogy et al. (1989b) ont démontré définitivement que ces neurones exprimant l ARN messager de la cholécystokinine sont bien des neurones dopaminergiques du mésencéphale ventral. Ainsi, une proportion significative des fibres dopaminergiques innervant toutes les composantes du striatum utilisent également la cholécystokinine comme neuromédiateur. L hypothèse de l implication de la cholécystokinine dans les processus pathologiques tels que la schizophrénie (cf. paragraphe IV. 1.) impose d étudier la distribution de neurones contenant l ARN messager de la cholécystokinine dans le mésencéphale humain. Une première étude démontrant l absence de tels neurones conclut à l existence d une différence d espèce (Palacios et al., 1989). Pour investiguer cette possibilité, nous avons analysé l expression de cholécystokinine dans les neurones dopaminergiques du mésencéphale ventral de différentes espèces de mammifères, hamster, cobaye, chat, singe et homme avec de plus l étude de patients atteints de schizophrénie (articles 10, 11). Ceci démontre que l expression de cholécystokinine dans le mésencéphale ventral est différente selon les espèces. En effet, chez le chat et le singe comme chez le rat les neurones dopaminergiques expriment la cholécystokinine alors qu ils ne l expriment pas chez le cobaye et le hamster. Dans le ganglion spinal également l expression de cholécystokinine diffère selon les espèces avec une expression chez le cobaye (Seroogy et al., 1990) et non chez le rat (Seroogy et al., 1990; Schiffmann et al., 1991) (cf. Thèse Annexe III.). Chez l homme normal, des neurones exprimant la cholécystokinine sont essentiellement présents dans la région centrale paranigrale {paranigralis area). Néanmoins, dans deux cas sur cinq, un faible marquage est également observé dans la substance noire pars compacta. Par contre, chez tous les patients souffrant de schizophrénie, un signal significatif correspondant à la présence d ARN messager de la cholécystokinine est observé dans les différentes subdivisions du mésencéphale ventral. Certaines cellules exprimant la cholécystokinine sont caractérisées par leur contenu en pigment mélanique ou leur expression de tyrosine hydroxylase, marqueurs des neurones dopaminergiques. La distribution de l ARN messager encodant la tyrosine hydroxylase est, pour sa part, strictement identique chez les contrôles et chez les patients souffrant de schizophrénie. Ceci démontre que le nombre de neurones dopaminergiques et leur capacité d expression de l enzyme clé de la synthèse de dopamine ne sont pas altérés chez ces patients. La signification précise de ces résultats reste à démontrer, d autant qu une augmentation d activité cholécystokininergique dans la voie mésolimbique chez les patients schizophrènes serait en opposition avec l effet de type neuroleptique décrit pour ce peptide chez l animal (Wang et al., 1984). Cette augmentation d expression d ARN messager de la cholécystokinine pourrait être directement liée à la pathologie schizophrénique mais elle pourrait également être la conséquence des traitements neuroleptiques auxquels, contrairement aux contrôles, ces patients sont soumis. Des études ultérieures seront nécessaires pour interpréter valablement cette observation.
 Comme mentionné ci-dessus, la présence de neurones arborant une")
65 -56- IV.2.C Régulation dopaminergique de l expression de l ARN messager encodant la cholécystokinine dans le striatum (article 12) Comme mentionné ci-dessus, la présence de neurones arborant une cholécystokinine-ir dans le striatum lui-même a été très discutée (cf. paragraphe IV.l.)- L hybridation in situ nous a permis de démontrer l absence de neurones exprimant l ARN messager de la cholécystokinine dans le striatum adulte normal (cf. paragraphe IV.2.b). Néanmoins, des études utilisant l analyse par Northern blot décrivent une quantité faible mais significative de cet ARN messager dans le striatum (Yoshikawa and Aizawa, 1988; Takeda et al., 1989; Ding and Mocchetti, 1992). De plus, par la même technique, une augmentation transitoire mais faible est observée au cours des premières semaines de développement post-natal (Yoshikawa and Aizawa, 1988; Takeda et al., 1989). Des lésions de déafférentation ou des manipulations pharmacologiques de récepteurs post-synaptiques induisent une plasticité fonctionnelle résultant en des modifications des niveaux d expression de neuromédiateurs (pour revue voir Graybiel, 1990) (cf. paragraphe II.6. et III.4.). De plus, de telles lésions ou traitements peuvent induire l expression d ARN messagers non détectés dans des conditions normales (Noguchi et al., 1989; Cortès et al., 1990; Delrée et al., 1992). De telles données et la fonction déterminante de la voie dopaminergique dans le contrôle de l expression génétique dans le striatum, nous ont mené à investiguer son rôle dans l expression éventuelle de l ARN messager de la cholécystokinine (article 12). La lésion de la voie nigro-striée par la 6-hydroxydopamine induit l expression de cet ARN messager dans un nombre important de neurones du striatum, constituant probablement une sous-population de neurones efférents ("spiny type I"), et ceci principalement dans le quadrant dorso-latéral (fig. 1 article 12). Par contre, l administration chronique d antagonistes des récepteurs dopaminergiques D2 tel que l halopéridol ou des récepteurs DI tel que le SCH23390 n ont aucun effet. Malgré cette absence d effet d un antagoniste D2, différents arguments reposant sur la localisation du récepteur D2 et les modifications de son expression (Brené et al., 1990), suggèrent que cette induction d expression d ARN messager de la cholécystokinine dans le striatum est médiée par le récepteur D2. Néanmoins, et de manière surprenante, au moment où nous avons clôturé cette étude, Ding et Mogenson (1992) ont publié un travail similaire utilisant la technique de Northern blot et démontrant des résultats strictement opposés aux nôtres, à savoir une diminution de l expression de l ARN messager de la cholécystokinine dans le striatum induite par la lésion de la voie nigro-striée. Des différences méthodologiques quant à la cinétique des événements étudiés constituent l explication la plus probable de ces divergences. En effet, nous avons effectué nos observations à des temps éloignés de la lésion, 18 et 28 jours, alors que Ding et Mogenson (1992) ont utilisé un temps précoce de 11 Jours. Cette évolution biphasique peut être corrélée avec la démonstration par Angulo et al. (1991) d une diminution transitoire et précoce à 8 jours suivie d une augmentation marquée et soutenue à 15 et 21 jours de l expression de l ARN messager du récepteur D2 dans le striatum après cette même lésion. Des études ultérieures seront nécessaires pour préciser le(s) mécanisme(s) impliqué(s).

66 -57- Quoiqu il en soit, cette étude démontre qu une large population de neurones striataux possèdent la capacité d exprimer l ARN messager de la cholécystokinine. Le signal étant faible, il reste probable que ces neurones ne constituent pas, même dans des conditions de stimulation, une source prépondérante de cholécystokinine dans le striatum. Néanmoins, la nature efférente probable de ces neurones suggère que la cholécystokinine ait une fonction dans les structures cibles du striatum que sont le globus pallidus et la substance noire pars reticulata et que celle-ci se trouve renforcée dans des conditions de déplétion dopaminergique. IV.2.d Ontogenèse des récepteurs de la cholécystokinine dans le striatum humain (article 13) Comme nous l avons rappelé dans le paragraphe III.5., la compartimentalisation d un élément neurochimique du striatum peut varier au cours du développement démontrant que les mécanismes de détermination et d établissement de cette compartimentalisation sont complexes. En effet, nombre de ces éléments, y compris le récepteur A2 de l adénosine (paragraphe III.5.), sont abondants dans les striosomes au cours du développement et ont une distribution homogène ou prépondérante dans la matrice à l âge adulte (Lowenstein et al., 1989; Murrin and Zeng, 1989; Nastuk and Graybiel, 1989; pour revue voir Graybiel, 1990). Nous avons étudié la distribution des sites de liaison de la cholécystokinine par autoradiographie à l aide de CCK8 marqué à l ^^Sj p^j. jg réactif de Bolton et Hunter dans le striatum humain foetal (18 semaines de gestation), néonatal et adulte. Cette distribution a été comparée avec celle de l activité de l AChE chez l adulte et le nouveau-né et celle des sites de liaison d un antagoniste sélectif du récepteur dopamine Dl, [^H]SCH23390 chez le foetus. Une distribution hétérogène des récepteurs de la cholécystokinine est observée dans le striatum adulte avec des zones circulaires ou allongées de moindre intensité de marquage. Dans le noyau caudé, elles correspondent aux striosomes pauvres en AChE, alors que dans le putamen leur correspondance avec les compartiments striataux ne peut être clairement établie. Dans le striatum néonatal également, les récepteurs de la cholécystokinine sont distribués de manière hétérogène avec des zones de moindre intensité de marquage. A cette période, dans le caudé comme dans le putamen, celles-ci correspondent aux striosomes enrichis en AChE. Dans le striatum foetal, par contre, les récepteurs de la cholécystokinine sont abondants voire exclusivement présents dans des aires de forme allongée dorsoventralement qui correspondent sur des coupes adjacentes aux régions enrichies en récepteurs dopaminergiques Dl. Ces derniers sont comme l AChE, des marqueurs des striosomes au cours du développement. Ces résultats démontrent que comme d autres constituants du striatum, les récepteurs de la cholécystokinine sont exprimés de manière prépondérante dans les striosomes au cours du développement et dans la matrice à l âge adulte. Néanmoins, ils démontrent aussi que les mécanismes de maturation de la compartimentalisation ne sont pas nécessairement simultanés pour tous les marqueurs ayant une évolution pourtant qualitativement semblable. En effet, à la naissance, les récepteurs de la cholécystokinine ont déjà, contrairement à l AChE (Graybiel and Ragsdale, 1980), la distribution matricielle caractéristique de l adulte.
.")
67 -58- De même, chez le chat, la tyrosine hydroxylase, marqueur des afférences dopaminergiques, évolue vers sa distribution matricielle adulte légèrement plus tardivement encore que l AChE (Graybiel, 1984). On peut ainsi indirectement supposer qu il existe une étape intermédiaire de développement se caractérisant par des striosomes riches en fibres dopaminergiques et pauvres en récepteurs de la cholécystokinine et une matrice pauvre en fibres dopaminergiques et riche en récepteurs de la cholécystokinine. Cette période constitue une transition entre deux situations où ces deux éléments sont distribués de manière similaire soit dans les striosomes soit dans la matrice. IV.3. Conclusions et perspectives Nous avons dans le présent chapitre étudié la distribution de la cholécystokinine par immunohistochimie et hybridation in situ et celle de ses récepteurs par autoradiographie dans les noyaux de la base, avec une référence particulière à l homme ainsi que la régulation de l expression de ce neuropeptide dans le striatum de rat. Nous avons démontré l existence dans le noyau caudé humain d une concentration de fines fibres à cholécystokinine dans certains striosomes ainsi qu un net enrichissement en fibres dans le striatum ventral. Dans la substance innominée, de nombreuses fibres cholécystokininergiques à aspect laineux ou "woolly fibers" sont présentes et leur distribution suggère qu elles soient plutôt des constituants de l extension amygdalienne ou "extended amygdala" que des fibres striato-pallidales du système ventral. Des modèles animaux devraient permettre de démontrer l origine exacte de ces fibres. Ces résultats confirment chez l homme l abondance de l innervation cholécystokininergique du striatum ventral décrite chez le rat (Hôkfelt et al., 1980a,b; Vanderhaeghen et al., 1980a,b). En outre, l abondance de ce neuropeptide dans la substance innominée qui contient des éléments des striatum et pallidum ventraux, de l amygdale ("extended amygdala") et les neurones cholinergiques du noyau basal de Meynert (cf. paragraphe II.2.b), démontre également son implication prépondérante dans le système limbique et potentielle dans certaines affections psychiatriques (Alheid and Heimer, 1988). Dans le striatum adulte humain. Inversement à la cholécystokinine elle-même, les récepteurs de la cholécystokinine sont généralement plus abondants dans la matrice que dans les striosomes. Au cours du développement les récepteurs de la cholécystokinine sont, comme d autres éléments du striatum, sélectivement exprimés par les neurones des striosomes. Nos résultats indiquent de plus que les mécanismes de maturation de la compartimentalisation ne sont pas nécessairement simultanés pour tous les éléments neurochimiques du striatum. L hybridation in situ, confirmant les principales données immunohistochimiques, démontre l absence de neurones exprimant l ARN messager de la cholécystokinine dans le striatum de rat normal. Par contre, et contre toute attente à l analyse de ces mêmes données immunohistochimiques, dans les diverses structures afférentes du striatum telles que le cortex
68 -59- cérébral, le thalamus ou l amygdale, une proportion très importante des neurones expriment TARN messager de la cholécystokinine. Burgunder et Young (1988, 1990) ont démontré que parmi ceux-ci, certains projettent effectivement vers le striatum et sont donc susceptibles d être source de fibres cholécystokininergiques. Ces différentes données obtenues par hybridation in situ indiquent que la cholécystokinine est très probablement présente dans la voie thalamo-corticale ainsi que dans les voies cortico- et thalamo-striatales dans lesquelles elle co-existerait avec le glutamate. Le(s) fonction(s) de ce peptide dans ces voies majeures du système des ganglions de la base reste(nt) à déterminer. L existence d une expression de l ARN messager de la cholécystokinine dans les neurones mésencéphaliques dopaminergiques a été démontrée dans diverses espèces y compris chez l homme où cette expression était controversée. Cette expression de l ARN messager de la cholécystokinine dans les neurones mésencéphaliques dopaminergiques est plus abondante chez les patients souffrant de schizophrénie que chez des patients contrôles. Chez ces derniers elle est en effet soit détectable mais plus faible (2cas/5) ou non-détectable (3cas/5). La participation de la cholécystokinine dans la pathogénie de la schizophrénie a été évoquée (cf. paragraphe IV. 1) par son abondance dans le système mésolimbique. La signification précise de nos résultats dans le cadre de cette hypothèse reste néanmoins à démontrer car cette augmentation d expression pourrait être liée à d autres paramètres que la pathologie schizophrénique. L utilisation de modèles expérimentaux chez le rat indique que certains neurones intrinsèques du striatum possèdent la capacité d exprimer l ARN messager de la cholécystokinine. De plus cette expression est régulée, comme celle d autres neuromédiateurs du striatum, par la voie dopaminergique nigro-striée, probablement par l intermédiaire du récepteur D2 de la dopamine. La cinétique de ces événements de régulation, le(s) récepteur(s) dopaminergique(s) impliqué(s), la caractérisation phénotypique et celle des projections de ces neurones et la fonction de la cholécystokinine dans leurs structures cibles en conditions normales et pathologiques sont autant de paramètres qui restent à déterminer.
69 -60- V, BIBLIOGRAPHIE

70 -61- Albin R.L., Young A.B. and Penney B. The functional anatomy of basal ganglia disorders. Trends Neurosci., 12: , Alexander G.E. and Crutcher M.D. Functional architecture of basal ganglia circuits: neural substrates of parallel Processing. Trends Neurosci., 13: , Alexander G.E., DeLong M.R. and Strick P.L. Parallel organization of functionally segregated circuits linking basal ganglia and cortex. Ann. Rev. Neurosci., 9: , Alexander S.P. and Reddington M. The cellular localization of adenosine receptors in rat neostriatum. Neuroscience, 28: , Alheid G.F. and Heimer L. New perspectives in basal forebrain organization of spécial relevance for neuropsychiatrie disorders: The striatopallidal, amygdaloid, and corticopetal components of substantia innominata. Neuroscience., 27: 1-39, Anderson K.D. and Reiner A. Extensive co-occurence of substance P and dynorphin in striatal projection neurons: an evolutionarily conserved feature of basal ganglia organization. J. Comp. Neurol., 295: , Angulo J.A,, Corini H., Ledoux M. and Schumacher M. Régulation by dopaminergic neurotransmission of dopamine D2 mrna and receptor levels in the striatum and nucléus accumbens of the rat. Mol. Brain Res., 11, , Aronin N., Cooper P.E., Lorenz L.J., Bird E.D., Sagar S.M., Leeman S.E. and Martin J.B. Somatostatin is increased in the basal ganglia in Huntington s disease. Ann. Neurol., 13; , Aronin N., DiFiglia M., Graveland G.A., Schwartz W.F. and Wu J.Y. Localization of immunoreactive enkephalins in GABA synthesizing neurons of the rat neostriatum. Brain Res., 300: , Beal F.M., Kowall N.W., Ellison D.W., Mazurek M.F., Swartz K.J. and Martin J.B. Réplication of the neurochemical characteristics of Huntington s disease by quinolinic acid. Nature, 321: , Beckstead R.M. Long collateral branches of substantia nigra pars reticulata axons to thalamus, superior colliculus and reticular formation in monkey and cat. Multiple rétrogradé neuronal labeling with fluorescent dyes. Neuroscience, 10: , Beckstead R.M. and Cruz C.J. Striatal axons to the globus pallidus, entopeduncular nucléus and substantia nigra corne mainly from separate cell populations in cat. Neuroscience, 19: , Beinfeld M.C., Meyer D.K., Eskay R.L., Jensen R.T. and Brownstein M.J. The distribution of cholecystokinin immunoreactivity in the central nervous System of the rat as determined by radioimmunoassay. Brain Res., 212; 51-57, Bergman H., Wichmann T. and DeLong M.R. Reversai of experimental parkinsonism by lésions of the subthalamic nucléus. Science, 249: , Besson M.J., Grayhiel A.M. and Nastuk M.A. (^H)SCH binding to Dj dopamine receptors in the basal ganglia of the cat and primate: délinéation of striosomal compartments and pallidal and nigral subdivisions. Neuroscience, 26: , Besson M.J., Grayhiel A.M. and Quinn B. Co-expression of neuropeptides in the cat s striatum: an immunohistochemical study of substance P, dynorphin B and enkephalin. Neuroscience, 39: 33-58, Birnhaumer L., Ahramowitz J. and Brown A.M. Receptor-effector coupling by G proteins. Bioch. Biophys. Acta, 1031: , 1990.

71 -62- Bjôrklund A. and Hôkfelt T. (Eds), Handbook of Chemical Neuroanatomy, GABA and Neuropeptides in the CNS, Part I, Vol. 4, Elsevier, Amsterdam. Bolam J.P., Clarke D.J., Smith A.D. and Somogyi P. A type of aspiny neuron in the rat neostriatum accumulâtes [^H]gamma-amino-butyric acid: Combination of Golgi-staining, autoradiography, and électron microscopy. J. Comp. Neurol., 213: , Bolam J.P., Wainer B.H. and Smith A.D. Characterization of cholinergic neurons in the rat neostriatum. A combination of choline acetyltransferase immunocytochemistry, golgi imprégnation and électron microscopy. Neuroscience, 12: , Brené S., Lindefors N., Herrera-Marschitz M. and persson H. Expression of dopamine D2 receptor and choline acetyltransferase mrna in the dopamine deafferented rat caudateputamen. Exp. Brain Res., 83: , Bruns R.F., Daly J.W. and Snyder S.H. Adenosine receptors in brain membranes: Binding of V^-cyclohexyl[3H]adenosine and l,3-diethyl-8-[^h]phenylxanthine. Proc. Natl. Acad. Sci. USA, 77: , Bruns R.F., Lu G.H. and Pugsley T.A. Characterization of the A2 adenosine receptor labeled by [^H]NECA in rat striatal neurons. Mol. Pharmacol., 29: , Bunzow J.R., Van Toi Grandy D.K., Albert P., Salon J., Christie McDonald, Machida C.A., Nere K.A. and Civelli O. Cloning and expression of a rat D2 dopamine receptor cdna. Nature, , Burgunder J.M. and Young III W.S. The distribution of thalamic projection neurons containing cholecystokinin messenger RNA, using in situ hybridization histochemistry and rétrogradé labeling. Mol. Brain Res., 4: , Burgunder J.M. and Young III W.S. Cortical neurons expressing the cholecystokinin gene in the rat: distribution in the adult brain, ontogeny, and some of their projections. J. Comp. Neurol., 300: 26-46, Caboche J., Rogard M., and Besson M.-J. Comparative development of Dj-dopamine and /i opiate receptors in normal and 6-hydroxydopamine-lesioned néonatal rat striatum: dopaminergic fibers regulate /x but not Dj receptor distribution. De\. Brain Res., 58: , Carpenter M.B., Batton R.R.,III, Carleton S.C. and Keller J.T. Interconnections and organization of pallidal and subthalamic neurons in the monkey. J. Comp. Neurol., 197: , Chang H.T., Wilson C.J. and Kitai S.T. Single neostriatal efferent axons in the globus pallidus: A light and électron microscopie study. Science, 213: , Chesselet M.-F. and Graybiel A.M. Striatal neurons expressing somatostatin-iike immunoreactivity: evidence for a peptidergic interneuronal System in the cat. Neuroscience, 17: , Chesselet M.-F. and Robbins E. Régional différences in substance P-like immunoreactivity in the striatum correlate with levels of pre-protachykinin mrna. Neurosci. Lett., 96: 47-53, Chesselet M.-F. and Robbins E. Characterization of striatal neurons expressing high levels of glutamic acid decarboxylase messenger RNA. Brain Res., 492: , Chevalier G. and Deniau J.M. Disinhibition as a basic process in the expression of striatal fonctions. Prends Neurosci., 13: 211-HQ, 1990.

72 -63- Chin J.H. Adenosine receptors in brain: Neuromodulation and rôle in epilepsy. Ann. Neurol., 26: , Christensson-Nylander M., Herrera-Marschitz M., Staines W., Hôkfelt T., Terenius L., Ungerstedt U., Cuello C., Oertel W.H. and Goldstein M. Striato-nigral dynorphin and substance P pathways in the rat. I. biochemical and immunohistochemical studies. Exp. Brain Res., 64: , Cortès R., Ceccatelli S., Schalling M. and Hôkfelt T. Differential effects of intracerebroventricular colchicine administration on the expression of mrnas for neuropeptides and neurotransmitter enzymes, with spécial emphasis on galanin: an in situ hybridization study. Synapse, 6: , Crossmann A.G. Parkinsonism degeneration of dopamine cells in pars compacta and ventral tegmental area. Neuroscience, 21: 1-40, Cuello A.C. and Paxinos G. Evidence for a long leu-enkephalin striopallidal pathway in rat brain. Nature (London), 271: Dahlstrôm A, and Fuxe K. Evidence for the existence of monoamine-containing neurons in the central nervous System. I. Démonstration of monoamines in the cell bodies of brain stem neurons. Acta Physiol. Scand., 62 (Suppl.232): 1-55, Dawbarn D., De Quidt M.E. and Emson P.C. Survival of basal ganglia neuropeptide Y- somatostatin neurones in Huntington s disease. Brain Res., 340: , DeLong M.R. Primate models of movement disorders of basal ganglia origin. Trends Neurosci., 13: , DeLong M.R. and Georogopoulos A.P. Functional organization of the substantia nigra, globus pallidus and subthalamic nucléus in the monkey. Can. J. Neurol. Sci., 6: 77, Delrée P., Schoenen J., Martin D., Rogister B., Leprince P., Sadzot C. and Moonen G. In vitro and in vivo modulation of neurotransmitters phenotype in adult rat DRG neurons. J. Neurosci. Res., in press, Demeulemeester H., Vandesande F. and Orban G.A. Immunocystochemical localization of somatostatin and cholecystokinin in the cat visual cortex. Brain Res., 332: , Desban M., Gauchy C., Kernel M.L., Besson M.J. and Glowinski J. Three-dimensional organization of the striosomal compartment and patchy distribution of striatonigral projections in the matrix of the cat caudate nucléus. Neuroscience, 29: , Dietl M.M. and Palacios J.M. The distribution of cholecystokinin receptors in the vertebrate brain: species différences studied by receptor autoradiography. J. Chem. Neuroanat., 2: , Dietl M.M., Probst A. and Palacios J.M. On the distribution of cholecystokinin receptor binding sites in the human brain: an autoradiographic study. Synapse, 1: , DiFiglia M. and Aronin N. Ultrastructural features of immunoreactive somatostatin neurons in the rat caudate nucléus. J. Neurosci., 2: , Ding X.-Z. and Mocchetti, I. Dopaminergic régulation of cholecystokinin mrna in rat content. Mol. Brain Res., 12: 77-83, Divac I., Fonnum F. and Storm-Mathisen J. High affinity uptate of glutamate in terminais of corticostriatal axons. Nature, 266: , Dixon R.A.F., Kobilka B.K., Strader D.J., Benovic J.L., Dohlman H.G., Frielle T., Boianowski M.A., Bennett C.D., Rands E., Diehl R.E., Mumford R.A., Slater E.E., Sigal I.S., Caron

73 -64- M. G., Lefkowitz R.J. and Strader C.D. Cloning of the gene and cdna for mammalian 13- adrenergic receptor and homology with rhodopsin. Nature, 321: 75-79, Dockray G.J., Gregory R.A., Hutchinson J.B., Harris J.I. and Runswick M.J. Immunochemical evidence of cholecystokinin-like peptides in brain. Nature, 274: , Dragunow M. and Faull R.L.M. Neuroprotective effects of adenosine. TiPS, 9: , Dunlap C.B. Pathologie changes in Huntington s chorea, with spécial reference to corpus striatum. Arch. Neurol. Psychiatry, 18: 867, Dunwiddie T.V. The physiological rôle of adenosine in the central nervous System. Int. Rev. Neurobiol., 27: , Durcan M.J. and Morgan P.F. Evidence for adenosine A2 receptor involvement in the hypomobility effects of adenosine analogues in mice. Eur. J. PharmacoL, 168: , Emson P.C., Rehfeld J.F., Langevin H. and Rossor M.N. Réduction in cholecystokinin-like immunoreactivity in the basal ganglia in Huntington s disease. Brain Res., 198: , Emson P.C., Rehfeld J.F. and Rossor M.N. Distribution of cholecystokinin-like peptides in the human brain. J. Neurochem., 38: , Ferkany J.W., Valentine H.L., Stone G.A. and Williams M. Adenosine Al receptors in mammalian brain: Species différences in their interactions with agonists and antagonists. Drug Dev. Res., 9: 85-93, Ferre S., Herrera-Marschitz M., Grabowska-Anden M., Ungerstedt U., Casas M. and Anden N. -E. Postsynaptic dopamine/adenosine interaction: I. Adenosine analogues inhibit dopamine D2-mediated behaviour in short-term reserpinized mice. Eur. J. PharmacoL, 192: 25-30, 1991a. Ferre S., Von Euler G., Johansson B., Fredholm B.B. and Fuxe K. Stimulation of adenosine A2 receptors decreases the affinity of dopamine D2 receptors in rat striatal membranes. Proc. Natl. Acad. Sci. USA, 88: , 1991b. Fishell G. and van der Kooy D. Pattern formation in the striatum: developmental changes in the distribution of striatonigral neurons. J. Neurosci., 7: , Folstein M.F., Brandt J. and Starkstein S. Cognition in Huntington s disease: characteristics and correlates. In; Function and dysfunction in the basal ganglia. Franks A.J. et al. (Eds), pp , University Press Manchester, Manchester and New York, Fox C.A., Andrade A.N., LuQui I.J. and Rafois J.A. The primate globus pallidus: A Golgi and électron microscopie study. J. Hirnforsch., 15: 75-93, Fredholm B.B. and Dunwiddie T.V. How does adenosine inhibit transmitter release? TiPS, 9; , Fuxe K. Evidence for the existence of monoamine neurones in the central nervous System. IV. Distribution of monoamine nerve terminais in the central nervous System. Acta Physiol. Scand., 64 (Suppl.247): , Fuxe K. and Ungerstedt U. Action of caffeine and theophyllamine on supersensitive dopamine receptors: Considérable enhancement of receptor response to treatment with dopa and dopamine receptor agonists. Med. Biol., 52: 48-54, Gerfen C.R. The neostriatal mosaic: Compartmentalization of corticostriatal input and striatonigral output System. Nature (London), 311: , Gerfen C.R. The neostriatal mosaic. I. Compartmental organization of projections from the striatum to the substantia nigra in the rat. J. Comp. Neurol., 236: , 1985.

74 -65- Gerfen C.R. Synaptic organization of the striatum. J. Electron Miscosc. Tech., 10: , Gerfen C.R. The neostriatal mosaic: Striatal patch-matrix organization is related to cortical lamination. Science, 246: , Gerfen C.R., Engber T.M., Mahan L.C., Susel Z., Chase T.N., Monsma Jr. F.J. and Sibley D.R. DI and D2 dopamine receptor-regulated gene expression of striatonigral and striatopallidal neurons. Science, 250: , 1990 Gerfen C.R., Herkenham M. and Thibault J. The neostriatal mosaic. II. Compartmental organization of mesostriatal dopaminergic and non-dopaminergic Systems. J. Neurosci., 7: , Gerfen C.R., McGinty J.F. and Young III W.S. Dopamine differentially régulâtes dynorphin, substance P, and enkephalin expression in striatal neurons: In situ hybridization histochemical analysis. J. Neurosci., 11: , Gerfen C.R. and Young III W.S. Distribution of striatonigral and striatopallidal peptidergic neurons in both patch and matrix compartments: an in situ hybridization histochemistry and fluorescent rétrogradé tracing study. Brain Res., 460: , Gilles C., Lotstra F. and Vanderhaeghen J.-J. CCK nerve terminais in the rat striatum and limbic areas originate partly in the brain stem and partly in telencephalic structures. Life Sci., 32: , Graveland G.A., Williams R.S. and DiFiglia M. A Golgi study of the human neostriatum: Neurons and afferent fibers. J. Comp. Neurol., 234: , Grayblel A.M. Correspondence between the dopamine islands and striosomes of the mammalian striatum. Neuroscience, 13: , Graybiel A.M. Neurotransmitters and neuromodulators in the basal ganglia. Trends Neurosci., 13: , Graybiel A.M., Hirsch E.C. and Agid Y.A. Différences in tyrosine hydroxylase-like immunoreactivity characterize the mesostriatal innervation of striosomes and extrastriosomal matrix at maturity. Proc. Natl. Acad. Sci. USA, 84: , Graybiel A.M. and Ragsdale C.W. Jr. Histochemically distinct compartments in the striatum of human, monkey and cat demonstrated by acetylthiocholinesterase staining. Proc. Nat. Acad. Sci. USA, 75: , Graybiel A.M. and Ragsdale C.W. Jr. dumping of acetylcholinesterase activity in the developing striatum of the human fétus and young infant. Proc. Natl. Acad. Sci. USA, 77: , Graybiel A.M., Ragsdale C.W. Jr., Yoneika E.S. and Eide R.P. An immunohistochemical study of enkephalins and other neuropeptides in the striatum of the cat with evidence that the opiate peptides are arranged to form mosaic patterns in register with the striosomal compartments visible with acetylcholinesterase staining. Neuroscience, 6: , 1981a. Graybiel A.M., Pickel V.M., Joh T.H., Reis D.J. and Ragsdale C.W.Jr. Direct démonstration of a correspondence between the dopamine islands and acetylcholinesterase patches in the developing striatum. Proc. Natl. Acad. Sci. USA, 78: , 1981b. Green R.D., Proudfit H.K. and Yeung S.-M. H. Modulation of striatal dopaminergic fonction by local injection of 5 -N-ethylcarboxamide adenosine. Science, 218: 58-61, Greenamyre J.T. and Young A.B. Synaptic localization of striatal NMDA, quisqualate and kainate receptors. Neurosci. Lett., 101: , 1989.

75 -66- Groenewegen H.J. Organization of the afferent connections of the mediodorsal thalamic nucléus in the rat, related to the mediodorsal-prefrontal topography. Neuroscience, 24: , Groenewegen H.J., Arnolds D.E.A.T. and Lopes, da Silva F.H. Afferent connections of the nucléus accumbens in the cat, with spécial emphasis on the projections from the hippocampal région. An anatomical and electrophysiological study. In: The Neurobiology of the Nucléus Accumbens (R.B. Chronister and J.F. DeFrance, eds), pp , Hear Institute for Electrophysiological Research, Brunswick, Maine. Groenewegen H.J. and Russchen F.T. Organization of the efferent projections of the nucléus accumbens to pallidal, hypothalamic and mesencephalic structures: A tracing and immunohistochemical study in the cat. J. Comp. Neurol., 223: , Guennoun R. and Bloch B., D2 dopamine receptor gene expression in the rat striatum during ontogeny: an in situ hybridization study. Dev. Brain Res. 60: 79-87, Guillet R. Néonatal caffeine exposure alters adenosine receptor control of locomotor activity in the developing rat. Dev. Pharmacol. Ther., 15: , Haber S.N. and Eide R. Corrélation between met-enkephalin and substance P immunoreactivity in the primate globus pallidus. Neuroscience, 6: , Haber S.N. and Nauta W.J.H. Ramifications of the globus pallidus in the rat as indicated by patterns of immunohistochemistry. Neuroscience, 9: , Haber S.N. and Watson S.J. The comparative distribution of enkephalin, dynorphin and substance P in the human globus pallidus and basal forebrain. Neuroscience, 14: , Heimer L., Switzer R.D. and Van Hoesen G.W. Ventral striatum and ventral pallidum. Components of the motor System? Trends Neurosci., 5: 83-87, Heimer L., Zaborszky L., Zahm D.S. and Alheid G.F. The ventral striatopallidothalamic projection. I. The striatopallidal link originating in striatal parts of the olfactory tubercle. J. Comp. Neurol., 255: , Herkenham M. and Pert C.B. Mosaic distribution of opiate receptors, parafascicular projections and acetylcholinesterase in rat striatum. Nature, 291: , Hill D.R., Campbell N.J., Shaw T.M. and Woodruff G.N. Autoradiographic localization and biochemical characterization of peripheral type CCK receptors in rat CNS using highly sélective non-peptide antagonists. J. Neurosci., 7: , Hôkfelt T., Herrera-Marschitz M., Seroogy K., Gong J., Staines W.A., Holets V., Schalling M., Ungerstedt U., Post C., Rehfeld J.F., Frey P., Fischer J., Dockray G., Hamaoka T., Walsh J.H. and Goldstein M. Immunohistochemical studies on cholecystokinin (CCK)- immunoreactive neurons in the rat using sequence spécifie antisera and with spécial reference to the caudate nucléus and primary sensory neurons. J. Chem. Neuroanat., 1: 11-52, Hôkfelt T., Rehfeld J.F., Skirboll L., Ivemark B., Goldstein M. and Markey K. Evidence for coexistence of dopamine anf CCK in meso-limbic neurons. Nature, 285: , 1980a. Hôkfelt T., Skirboll L., Goldstein M., Markey K. and Dann O. A subpopulation of mesencephalic dopamine neurons projecting to limbic areas contains a cholecystokinin-like peptide: evidence from immunohistochemistry combined with rétrogradé tracing. Neuroscience, 5: , 1980b.

76 -67- Hornykiewicz O. Die topische lokalisation und das verhalten von noradrenalin und dopamin (3- Hydroxytyramin) in der substantia nigra des normalen und Parkinsonkranken menschen. Klin. Wochenschr., 75: , Innis R.B., Correa F.M.A., Uhl G.R., Schneider B. and Snyder S,H. Cholecystokinin octapeptide-like immunoreactivity: histochemical localization in the rat brain. Proc. Natl. Acad. Sci. USA, 76: , Jarvis M.F. and Williams M. Adenosine and dopamine function in the CNS. TiPS, 8: , Jarvis M.F. and Williams M. Direct autoradiog^raphic localization of adenosine A2 receptors in rat brain using the A2-selective agonist, [^H]-CGS Eur. J. Pharmacol., 168: , Jarvis M.F., Schulz R., Hutchinson A.J., Do U.H., Sills M. and Williams M. [^H]CGS21680, a sélective A2 adenosine agonist directly labels A2 receptors in rat brain. J. Pharmacol. Exp. Ther., 251:^88-893, Jimenez-Castellanos J. and Graybiel A.M. Subdivisions of the dopamine-containing A8-A9- AlO complex identified by their differential mesostriatal innervation of striosomes and extrastriosomal matrix. Neuroscience, 23: , Joyce J.N., Sapp D.W. and Marshall J.F. Human striatal dopamine receptors are organized in compartments. Proc. Natl. Acad. Sci. USA, 83: , Kernel M.-L., Desban M., Glowinski J. and Gauchy C. Distinct presynaptic control of dopamine release in striosomal and matrix areas of the cat caudate nucléus. Proc. Natl. Acad. Sci., 86: , Kemp J.M. and Powell T.P.S. The corticostriate projection in the monkey. Brain, 93: , Krebs M.-O., Trovero F., Desban M., Gauchy C., Glowinski J. and Kemei M.-L. Distinct presynaptic régulation of dopamine release through NMDA receptors in striosome- and matrix-enriched areas of the rat striatum. J. Neurosci., 11: , Kritzer M.F., Innis R.B. and Goldman-Rakic P.S. Régional distribution of cholecystokinin binding sites in macaque basal ganglia determined by in vitro receptor autoradiography. Neuroscience, 38: 81-92, Le Moine C., Normand E. and Bloch B. Phenotypical characterization of the rat striatal neurons expressing the Di dopamine receptor gene. Proc. Natl. Acad. Sci. USA, 88: , Le Moine C., Normand., Guitteny A.F., Fouque B., Teoule R. and Bloch B. Dopamine receptor gene expression by enkephalin neurons in rat forebrain. Proc. Natl. Acad. Sci. USA, 87: , Lewy F.H. Zur pathologisch-anatomischen differential Diagnose der Paralysis agitans und der Huntingtonschen Chorea. Z. Gesamte Neurol. Psychiat., 73: , Libert F., Parmentier M., Lefort A., Dinsart C., Van Sande J., Maenhaut C., Simons M.-J., Dumont J.E. and Vassart G. Sélective amplification and cloning of four new members of the G protein-coupled receptor family. Science, 244: , Libert F., Passage E., Parmentier M., Simons M.-J., Vassart G. and Mattéi M.-G. Chromosomal mapping of Al and A2 adenosine receptors, VIP receptor, and a new subtype of serotonin receptor. Genomics, 11: , 1991a.

77 -68- Libert F., Vassart G. and Parmentier M. Current developments in G-protein-coupled receptors. Current Opinion in Cell Biology, 3: , 1991b. Lindefors N., Brodin E. and Ungerstedt U. Neurokinin A and substance P in striatonigral neurons in rat brain. Neuropeptides, 8: , Lowenstein P.R., Slesinger P.A., Singer H.S., Walker L.C., Casanova M.F., Raskin L.S., Price D.L. and Coyle J.T. Compartment-specific changes in the density of choline and dopamine uptake sites and muscarinic and dopaminergic receptors during the development of the baboon striatum; a quantitative receptor autoradiographic study. J. Comp. Neurol., 288: , Lupica C.R., Cass W.A., Zahniser N.R. and Dunwiddie T.V. Effects of the sélective adenosine A2 receptor agonist CGS on in vitro electrophysiology, camp formation and dopamine release in rat hippocampus and striatum. J. Pharmacol. Exp. Ther., 252: , Malach R. and Graybiel A.M. Mosaic architecture of the somatic sensory-recipient sector of the cat s striatum. J. Neurosci., 6: , Markstein R. and Hôkfelt T. Effect of cholecystokinin octapeptide on dopamine release from slices of cat caudate nucléus. J. Neurosci., 4: , Matsuda L.A., Lolait S.J., Brownstein M.J., Young A.C. and Bonner T.I, Structure of a cannabinoid receptor and functional expression of the cloned cdna. Nature, 346: , Meyer D.K., Beinfeld M.C., Oertel W.H. and Brownstein M.J. Origin of the cholecystokinincontaining fibers in the rat caudatoputamen. Science, 215: , Meyer D.K. and Krauss J. Dopamine modulâtes cholecystokinin release in neostriatum. Nature, 301: , Meyer D.K., Schultheiss K. and Hardung M. Bilateral ablation of frontal cortex reduces concentration of cholecystokinin-like immunoreactivity in rat dorsolateral striatum. Brain Res., 452: , Miller R.J. The revenge of the kainate receptor. Trends Neurosci., 14; , 1991a. Miller R.J. Metabotropic excitatory amino acid receptors reveal their true colors. TiPS, 12: , 1991b. Monaghan D.T. and Cotman C.W. Distribution of N-methyl-D-aspartate-sensitive L- [3H]Glutamate-binding sites in rat brain. J. Neurosci., 5: , Mugnaini E. and Oertel W.H. An atlas of the distribution of GABAergic neurons and terminais in the CNS as revealed by GAD immunohistochemistry. In: Handbook of Chemical Neuroanatomy, GABA and Neuropeptides in the CNS, Part I (Bjôrklund A. and Hôkfelt T., eds). Vol. 4, pp , Elsevier, Amsterdam. Murrin L.C. and Zeng W. Dopamine DI receptor development in the rat striatum: early localization in striosomes. Brain Res., 480: , Nastuk M.A. and Graybiel A.M. Ontogeny of Ml and M2 muscarinic binding sites in the striatum of the cat relationships to one another and to striatal compartmentalization. Neuroscience, 33: , Nathans J. and Hogness D.S. Isolation, sequence analysis, and intron-exon arrangement of the gene encoding bovine rhodopsin. Cell, 34: , 1983.

78 -69- Nauta H.J.W. Projections of the pallidal complex: An autoradiographic study in the cat. Neuroscience, 4; , 1979a. Nauta H.J.W. A proposed conceptual reorganization of the basal ganglia and telencephalon. Neuroscience, 4: , 1979b. Noguchi K., Senba E., Morita Y., Sato M. and Tohyama M. Prepro-VIP and preprotachykinin mrnas in the rat dorsal root ganglion cells following peripheral axotomy. Mol. Brain Res., 6: , Oertel W.H. and Mugnaini E. Immunocytochemical studies of gabaergic neurons in rat basal ganglia and their relations to other neuronal Systems. Neurosci. Lett., 47: , Onali P., Olianas M.C. and Bunse B. Evidence that adenosine A2 and dopamine autoreceptors antagonistically regulate tyrosine hydroxylase activity in rat striatal synaptosomes. Brain Res., 456: , Oison L., Seiger A. and Fuxe K. Heterogeneity of striatal and limbic dopamine innervation: highly fluorescent islands in developing and adult rats. Brain Res., 44: , Ottersen O.P. and Storm-Mathisen J. Neurons containing or acumulating transmitter amino acids. In: Handbook of Chemical Neuroanatomy, Classical Transmitter and Transmitter Receptors in the CNS, Part II (Bjôrklund A., Hôkfelt T. and Kuhar M.J., eds). Vol. 3, pp , Elsevier, Amsterdam. Palacios J.M., Savasta M. and Mengod G. Does cholecystokinin colocalize with dopamine in the human substantia nigra? Brain Res., 488: , Parent A. and Smith Y. Organization of efferent projections of the subthalamic nucléus in the squirrel monkey as revealed by rétrogradé labeling methods. Brain Res., 436: , Parent A., Smith Y., Filion M. and Dumas J. Distinct afferents to internai and external pallidal segments in the squirrel monkey. Neurosci. Lett., 96: , Parkinson F.E. and Fredholm B.B. Autoradiographic evidence for G-protein coupled A2- receptors in rat neostriatum using [^H]-CGS as a ligand. Naunyn-Schmiedeberg s Arch. Pharmacol., 342: 85-89, Penny G.R., Afsharpour S. and Kitai S.T. The glutamate decarboxylase-, leucine enkephalin-, méthionine enkephalin- and substance P-immunoreactive neurons in the neostriatum of the rat and cat: Evidence for partial population overlap. Neuroscience, 17: , Percheron G., Yelnik J. and François C. A Golgi analysis of the primate globus pallidus. III. Spatial organization of the striato-pallidal complex. J. Comp. Neurol., 227: , Petersen E.N. Sélective protection by adenosine receptor agonists against DMCM-induced seizures. Eur. J. Pharmacol., 195: , Phillips A.G. and Carr G.D. Cognition and the basal ganglia: A possible substrate for procédural knowledge. Can. J. Neurol. Sci., 14: , Ragsdale C.W.,Jr. and Graybiel A.M. Fibers from the basolateral nucléus of the amygdala selectively innervate striosomes in the caudate nucléus of the cat. J. Comp. Neurol., 269: , Reale R.A. and Imig T.J. Auditory cortical field projections to the basal ganglia of the cat. Neuroscience, 8: 67-86, 1983.

79 -70- Rehfeld J.F. Immunochemical studies on cholecystokinin. II. Distribution and molecular heterogeneity in the central nervous System and small intestine of man and hog. J. Biol. Chem., 253: , Reiner A., Albin R.L., Anderson K.D., D Amato C.J., Penney J.B. and Young A.B. Differential loss of striatal projection neurons in Huntington disease. Proc. Natl. Acad. Sci. USA, 85: , Reiner A. and Anderson K.D. The patterns of neurotransmitter and neuropeptide co-occurence among striatal projection neurons: conclusions based on recent findings. Brain Res. Rev., 15: , Riberio J.A. and Sébastian A.M, Adenosine receptors and calcium: Basis for proposing a third (A^) adenosine receptor. Prog. NeurobioL, 26: , Robberecht P., Deschodt-Lanckman M. and Vanderhaeghen J.-J. Démonstration of biological activity of brain gastrin-like peptidic material in the human: its relationship with the COOH-terminal octapeptide of cholecystokinin. Proc. Natl. Acad. Sci. USA, 75: , Samuel D., Errami M. and Nieoullon A. Localization of N-methyl-D-aspartate receptors in the rat striatum: Effects of spécifie lésions on the [3H]3-(2-carboxypiperazin-4-yl)propyl-lphosphonic acid binding. J. Neurochem., 54: , Savasta M., Dubois A. and Scatton B. Autoradiographic localization of DI dopamine receptors in the rat brain with (^H) SCH Brain Res., 375: , Schiffmann S.N., Campistron G., Tugendhaft P., Brotchi J., Flament-Durand J., Geffard M. and Vanderhaeghen J.-J. Immunocytochemical détection of GABAergic nerve cells in the human temporal cortex using a direct gamma-aminobutyric acid antiserum. Brain Res., 442: , Schiffmann S.N. and Vanderhaeghen J.-J. Immunocytochemical détection of gabaergic nerve cells in the human striatum and cerebellum using a gamma-aminobutyric acid antiserum. Neurochem. Int., 17: , Schiffmann S.N., Teugels E., Halleux P., Menu R. and Vanderhaeghen J.-J. Cholecystokinin mrna détection in rat spinal cord motoneurons but not in dorsal root ganglia neurons. Neurosci. Lett., 123: , Seguela P., Geffard M., Buijs R.M. and Le Moal M. Antibodies against gamma-aminobutyric acid: specificity studies and immunocytochemical results. Proc. Natl. Acad. Sci. USA, 81: , Seroogy K.B., Mehta A. and Fallon J.H. Neurotensin and cholecystokinin coexistence within neurons of the ventral mesencephalon: projections to forebrain. Exp. Brain Res., 68: , Seroogy K., Dangaran K., Lim S., Haycock J.W. and Fallon J.H. Ventral mesencephalic neurons containing both cholecystokinin and tyrosine hydroxylase-like immunoreactivities Project to forebrain régions. J. Comp. Neurol., 279: , 1989a. Seroogy K., Schalling M., Brene S., Dagerlind A., Chai S.Y., Hôkfelt T., Persson H., Brownstein M., Huan R., Dixon J.E., Filer D., Schlessinger D. and Goldstein M. Cholecystokinin and tyrosine hydroxylase messenger RNAs in neurons of rat mesencephalon: peptide/monoamine coexistence studies using in situ hybridization combined with immunocytochemistry. Exp. Brain Res., 74: , 1989b. Seroogy K., Mohapatra N.K., Lund P.K., Rethelyi M., McGehee D.S. and Perl E.R. Speciesspecific expression of cholecystokinin messenger RNA in rodent dorsal root ganglia. Mol. Brain Res., 7: , 1990.

80 -71- Sibley D.R. and Monsma F.J.,Jr. Molecular biology of dopamine receptors. TiPS, 13: 61-69, Smith Y., Bolam J.P. and Von Krosigk M. Topographical and synaptic organization of the GABA-containing pallidosubthalamic projection in the rat. Eur. J. Neurosci., 2: , 1990a. Smith Y., Hazrati L.-N. and Parent A. Efferent projections of the subthalamic nucléus in the squirrel monkey as studied by the PHA-L anterograde tracing method. J. Comp. Neurol., 294: , 1990b. Smith Y. and Parent A. Neuropeptide Y-immunoreactive neurons in the striatum of cat and monkey: morphological characteristics, intrinsic organization and co-localization with somatostatin. Brain Res., 372: , 1986a. Smith Y. and Parent A. Differential connections of caudate nucléus and putamen in the squirrel monkey (Saimiri sciureus). Neuroscience, 18: , 1986b. Smith Y. and Parent A. Neurons of the subthalamic nucléus in primates display glutamate but not GABA immunoreactivity. Brain Res., 453: , Smith Y., Parent A., Seguela P. and Descarries L. Distribution of GABA-Immunoreactive neurons in the basal ganglia of the squirrel monkey (Saimiri sciureus). J. Comp. Neurol., 259: 50-64, Snyder S.H. Adenosine as a neuromodulator. Ann. Rev. Neurosci., 8: , Snyder S.H., Katims J.J., Annau Z., Bruns R.F. and Daly J.W. Adenosine receptors and behavioral actions of methylxanthines. Proc. Natl. Acad. Sci. USA, 78: , Sokoloff P., Giros B., Martres M.-P., Bouthenet M.-L. and Schwartz J.-C. Molecular cloning and characterization of a novel dopamine receptor (D,) as a target for neuroleptics. Nature, 347: , Stone G.A., Jarvis M.F., Sills M.A., Weeks B., Snowhill E.W. and Williams M. Species différences in high-affinity adenosine A2 binding sites in striatal membranes from mammalian brain. Drug Dev. Res., 15: 31-46^, Stone T.W. Physiological rôles for adenosine and adenosine 5 -triphosphate in the nervous System. Neurosci., 6: , Stoof J.C. and Kebabian J.W. Opposing rôles for D-1 and D-2 dopamine receptors in efflux of cyclic AMP from rat neostriatum. Nature, 294: , Sugimoto T. and Mizuno N. Neurotensin in projection neurons of the striatum and nucléus accumbens, with reference to coexistence with enkephalin and GABA: an immunohistochemical study in the cat. J. Comp. Neurol., 257: , Sunahara R.K., Guan H.-C., O Dowd B.F., Seeman P., Laurier L.G., Ng G., George S.R., Torchia J., Van Toi H.H.M. and Niznik H.B. Cloning of the gene for a human dopamine Dj receptor with higher affinity for dopamine than Dj. Nature, 350: , Sunahara R.K., Niznik H.B., Weiner D.M., Stormann T.M., Brann M.R., Kennedy J.L., Gelernter J.E., Rozmahel R., Yang Y., Israël Y., Seeman P. and O Dowd B.F. Human dopamine D, receptor encoded by an intronless gene on chromosome 5. Nature, 347: 80-83, Takeda K., Koshimoto H., Uchiumi F., Haun S., Dixon J.E. and Kato T. Postnatal development of cholecystokinin-like immunoreactivity and its mrna level in rat brain régions. J. Neurochem., 53: , 1989.

81 -72- Tiberi M., Jarvie K.R., Silvia C., Falardeau P., Gingrich J.A., Godinot N., Bertrand L., Yang- Feng T.L., Fremeau R.T.,Jr. and Caron M.G. Cloning molecular characterization, and chromosomal assignment of a gene encoding a second D. dopamine receptor subtype: Differential expression pattern in rat brain compared with tne Dj^ receptor. Proc. Natl. Acad. Sci. USA. 88: , Trussell L.O. and Jackson M.B. Adenosine-activated potassium conductance in cultured striatal neurons. Proc. Natl. Acad. Sci. USA, 82: , Van Calker D., Muller M. and Hamprecht B. Adenosine régulâtes via two different types of receptors, the accumulation of cyclic AMP in cultured brain cells. J. Neurochem., 33: , Van Calker D., Steber R., Klotz K.-N. and Greil W. Carbamazepine distinguishes between adenosine receptors that médiate different second messenger responses. Eur. J. Pharmacol., 206: , Van Toi Bunzow J.R., Guan H.-C., Sunahara R.K., Seeman P., Niznik H.B. and Civelli O. Cloning of the gene for a human dopamine receptor with high affinity for the antipsychotic clozapine. Nature, 350: , Vanderhaeghen J.-J. Neuronal cholecystokinin. GABA and Neuropeptides in the CNS, Part I. In Handbook of Chemical Neuroanatomy (eds A. Bjôrklund and T. Hôkfelt), 4: pp Elsevier: Amsterdam, Vanderhaeghen J.-J., Lotstra, F., De Mey, J. and Gilles C. Immunohistochemical localization of cholecystokinin- and gastrin-like peptides in the brain and hypophysis of the rat. Proc. Natl. Acad. Sci. USA., 77: , 1980a. Vanderhaeghen J.-J., Lotstra F., Vierendeels G., Gilles C., Deschepper C. and Verbanck P. Cholecystokinins in the central nervous System and neurohypophysis. Peptides, 2(suppl2): 81-88, 1980b. Vanderhaeghen J.-J., Signeau J.C. and Gepts W. New peptide in the vertebrate CNS reacting with antigastrin antibodies. Nature, 257: , Vincent S.R., Skirboll L.R., Hôkfelt T., Johansson O., Lundberg J.M., Eide R.P., Terenius L. and Kimmel J. Coexistence of somatostatin and avian pancreatic polypeptide (APP)-like immunoreactivity in some forebrain neurons. Neuroscience, 7: , Voight M.M. and Wang R.Y. In vivo release of dopamine in the nucléus accumbens of the rat: Modulation by cholecystokinin. Brain Res., 296: , von Lubitz Dag K.J.E., Dambrosia J.M., Kempski O. and Redmond D.J. Cyclohexyl adenosine protects against neuronal death foliowing ischemia in the CAI région of Gerbil hippocampus. Stroke, 19: , Wojcik W.J. and Neff N.H. Differential location of adenosine Al and A2 receptors in striatum. Neurosci. Lett., 41: 55-60, Wang R.Y., White F.J. and Voigt M.M. Cholecystokinin, dopamine and schiophrenia. TIPS, 5: , Wong D.F., Wagner H.N.,Jr, Tune L.E., Dannals R.F., Pearlson G.D., Links J.M., Tamminga C.A., Broussolle E.P., Ravert H.T., Wilson A.A., Toung J.K.T., Malat J., Williams J.A., O Tuama L.A., Snyder S.H., Kuhar M.J. and Gjedde A. Positron émission tomography reveals elevated D2 dopamine receptors in drug-naive schizophrénies. Science, 234: , Yoshikawa, K. and Aizawa T. Transient increases and individual variations in preprocholecystokinin gene expression in rat telencephalic régions during development: a

82 -73- quantitative comparison between preprocholecystokinin mrna and preproenkephalin rarna. Mol. Brain Res., 4: 87-96, Young III W.S., Bonner T.I. and Brann M.B. Mesencephalic dopamine neurons regulate the expression of neuropeptides mrnas in the rat forebrain. Proc. Natl. Acad. Sci. USA, 83: , Zetler G. Neuropharmacological profile of cholecystokinin-like peptides. Arm. N.Y. Acad. Sci., 448: , Zetterstrôm T. and Fillenz M. Adenosine agonists can both inhibit and enhance in vivo striatal dopamine release. Eur. J. Pharmacol., 180: , Zigmond M.J., Abercrombie E.D., Berger T.W., Grâce A.A. and Stricker E.M. Compensations after lésions of central dopaminergic neurons: some clinical and basic implications. Prends in Neurosci., 13: , 1990.
83 -74- VI. THESES ANNEXES 1. Les neurones et fibres GABAergiques sont détectables dans le cerveau humain par immunohistochimie à l aide d un anticorps dirigé directement contre l acide gammaaminobutyrique. SCHIFFMANN, S., CAMPISTRON, G., TUGENDHAFT, P., BROTCHI, J., FLAMENT- DURAND, J., GEFFARD, M. and VANDERHAEGHEN, J-J. ; Immunocytochemical détection of GABAergic nerve cells in the human temporal cortex using a direct gammaaminobutyric antiserum. Brain Research, 442, , SCHIFFMANN, S.N. and VANDERHAEGHEN, J-J. : Immunocytochemical détection of GABAergic nerve cells in the human striatum and cerebellum using a gamma-aminobutyric acid antiserum. Neurochem. Int., 17, , Une augmentation de substance P et de met-enképhaline est observée dans le striatum sévèrement atrophié d un patient n ayant pas présenté de choréoathétose. SCHIFFMANN, S.N. and VANDERHAEGHEN, J-J. : Increase of Substance P and Metenkephalin in a severely atrophied striatum without clinical expression of chorea. Neurochem. Int., 14, , SCHIFFMANN, S.N. and VANDERHAEGHEN, J.-J.: Does the absence of clinical expression of choreoathetosis, despite severe striatal atrophy, correlate with plasticity of neuropeptide synthesis? J. Neural Transm., 33: , Les motoneurones de la corne antérieure de la moelle épinière du rat expriment l ARN messager encodant la cholécystokinine. SCHIFFMANN, S.N., TEUGELS, E., HALLEUX, P., MENU, R. and VANDERHAEGHEN, J.-J.: Cholecystokinin mrna détection in rat spinal cord motoneurons but not in dorsal root ganglia neurons. Neurosci. Lett., 123: , 1991.

84
Fonction de conduction de la moelle épinière. Dr F. TOUMI
 Fonction de conduction de la moelle épinière Dr F. TOUMI I. GENERALITES : Communication Moelle avec les centres supra spinaux Communication Intra segmentaire (ipsilatérale / controlatérale) Communication
Fonction de conduction de la moelle épinière Dr F. TOUMI I. GENERALITES : Communication Moelle avec les centres supra spinaux Communication Intra segmentaire (ipsilatérale / controlatérale) Communication
Onde P50 dans la schizophrénie
 37-285-A-14 Onde P50 dans la schizophrénie E. Houy, F. Thibaut Certains troubles cognitifs présentés par les patients schizophrènes sont interprétables en termes de dysfonctionnement du traitement de l
37-285-A-14 Onde P50 dans la schizophrénie E. Houy, F. Thibaut Certains troubles cognitifs présentés par les patients schizophrènes sont interprétables en termes de dysfonctionnement du traitement de l
DOCUMENTATION - FRANCAIS... 2
 DOCUMENTATION MODULE CATEGORIESTOPMENU MODULE CREE PAR PRESTACREA INDEX : DOCUMENTATION - FRANCAIS... 2 INSTALLATION... 2 CONFIGURATION... 2 LICENCE ET COPYRIGHT... 3 SUPPORT TECHNIQUE ET MISES A JOUR...
DOCUMENTATION MODULE CATEGORIESTOPMENU MODULE CREE PAR PRESTACREA INDEX : DOCUMENTATION - FRANCAIS... 2 INSTALLATION... 2 CONFIGURATION... 2 LICENCE ET COPYRIGHT... 3 SUPPORT TECHNIQUE ET MISES A JOUR...
AMENDMENT TO BILL 32 AMENDEMENT AU PROJET DE LOI 32
 THAT the proposed clause 6(1), as set out in Clause 6(1) of the Bill, be replaced with the following: Trustee to respond promptly 6(1) A trustee shall respond to a request as promptly as required in the
THAT the proposed clause 6(1), as set out in Clause 6(1) of the Bill, be replaced with the following: Trustee to respond promptly 6(1) A trustee shall respond to a request as promptly as required in the
AGRÉGATION DE SCIENCES DE LA VIE - SCIENCES DE LA TERRE ET DE L UNIVERS
 AGRÉGATION DE SCIENCES DE LA VIE - SCIENCES DE LA TERRE ET DE L UNIVERS CONCOURS EXTERNE ÉPREUVES D ADMISSION session 2010 TRAVAUX PRATIQUES DE CONTRE-OPTION DU SECTEUR A CANDIDATS DES SECTEURS B ET C
AGRÉGATION DE SCIENCES DE LA VIE - SCIENCES DE LA TERRE ET DE L UNIVERS CONCOURS EXTERNE ÉPREUVES D ADMISSION session 2010 TRAVAUX PRATIQUES DE CONTRE-OPTION DU SECTEUR A CANDIDATS DES SECTEURS B ET C
Dr E. CHEVRET UE2.1 2013-2014. Aperçu général sur l architecture et les fonctions cellulaires
 Aperçu général sur l architecture et les fonctions cellulaires I. Introduction II. Les microscopes 1. Le microscope optique 2. Le microscope à fluorescence 3. Le microscope confocal 4. Le microscope électronique
Aperçu général sur l architecture et les fonctions cellulaires I. Introduction II. Les microscopes 1. Le microscope optique 2. Le microscope à fluorescence 3. Le microscope confocal 4. Le microscope électronique
Fonctionnement neural et troubles cognitifs chez le patient bipolaire: preuve ou surinterprétation
 DIALOGUE ESPRIT / CERVEAU : CIRCULARITE CRP -OU Santé ANTAGONISME? Fonctionnement neural et troubles cognitifs chez le patient bipolaire: preuve ou surinterprétation Charles Pull Directeur du «Laboratoire
DIALOGUE ESPRIT / CERVEAU : CIRCULARITE CRP -OU Santé ANTAGONISME? Fonctionnement neural et troubles cognitifs chez le patient bipolaire: preuve ou surinterprétation Charles Pull Directeur du «Laboratoire
Les circuits neuronaux responsables du Sommeil paradoxal
 Les circuits neuronaux responsables du Sommeil paradoxal Pierre-Hervé Luppi DIU Physiologie et Pathologie du Sommeil, Lyon, Jeudi 18 Décembre 2014 Le sommeil paradoxal est présent chez le chat pontique
Les circuits neuronaux responsables du Sommeil paradoxal Pierre-Hervé Luppi DIU Physiologie et Pathologie du Sommeil, Lyon, Jeudi 18 Décembre 2014 Le sommeil paradoxal est présent chez le chat pontique
ROYAUME DE BELGIQUE / KINGDOM OF BELGIUM / KONINKRIJK BELGIE
 1 ROYAUME DE BELGIQUE / KINGDOM OF BELGIUM / KONINKRIJK BELGIE Service Public Fédéral Affaires Etrangères, Commerce extérieur et Coopération au Développement Federal Public Service Foreign Affairs, External
1 ROYAUME DE BELGIQUE / KINGDOM OF BELGIUM / KONINKRIJK BELGIE Service Public Fédéral Affaires Etrangères, Commerce extérieur et Coopération au Développement Federal Public Service Foreign Affairs, External
Calculation of Interest Regulations. Règlement sur le calcul des intérêts CONSOLIDATION CODIFICATION. Current to August 4, 2015 À jour au 4 août 2015
 CANADA CONSOLIDATION CODIFICATION Calculation of Interest Regulations Règlement sur le calcul des intérêts SOR/87-631 DORS/87-631 Current to August 4, 2015 À jour au 4 août 2015 Published by the Minister
CANADA CONSOLIDATION CODIFICATION Calculation of Interest Regulations Règlement sur le calcul des intérêts SOR/87-631 DORS/87-631 Current to August 4, 2015 À jour au 4 août 2015 Published by the Minister
DOCUMENTATION - FRANCAIS... 2
 DOCUMENTATION MODULE SHOPDECORATION MODULE PRESTASHOP CREE PAR PRESTACREA INDEX : DOCUMENTATION - FRANCAIS... 2 INSTALLATION... 2 Installation automatique... 2 Installation manuelle... 2 Résolution des
DOCUMENTATION MODULE SHOPDECORATION MODULE PRESTASHOP CREE PAR PRESTACREA INDEX : DOCUMENTATION - FRANCAIS... 2 INSTALLATION... 2 Installation automatique... 2 Installation manuelle... 2 Résolution des
AGROBASE : un système de gestion de données expérimentales
 AGROBASE : un système de gestion de données expérimentales Daniel Wallach, Jean-Pierre RELLIER To cite this version: Daniel Wallach, Jean-Pierre RELLIER. AGROBASE : un système de gestion de données expérimentales.
AGROBASE : un système de gestion de données expérimentales Daniel Wallach, Jean-Pierre RELLIER To cite this version: Daniel Wallach, Jean-Pierre RELLIER. AGROBASE : un système de gestion de données expérimentales.
La maladie de Huntington, une maladie du cerveau
 Actualités à propos de la recherche sur la maladie de Huntington. Expliqué simplement. Écrit par des scientifiques. Pour la communauté mondiale HD. La greffe de moelle osseuse dans la maladie de Huntington
Actualités à propos de la recherche sur la maladie de Huntington. Expliqué simplement. Écrit par des scientifiques. Pour la communauté mondiale HD. La greffe de moelle osseuse dans la maladie de Huntington
MEMORANDUM POUR UNE DEMANDE DE BOURSE DE RECHERCHE DOCTORALE DE LA FONDATION MARTINE AUBLET
 MEMORANDUM POUR UNE DEMANDE DE BOURSE DE RECHERCHE DOCTORALE DE LA FONDATION MARTINE AUBLET ATTENTION : Tout formulaire de candidature qui présentera des erreurs ou anomalies dans le remplissage des champs
MEMORANDUM POUR UNE DEMANDE DE BOURSE DE RECHERCHE DOCTORALE DE LA FONDATION MARTINE AUBLET ATTENTION : Tout formulaire de candidature qui présentera des erreurs ou anomalies dans le remplissage des champs
Credit Note and Debit Note Information (GST/ HST) Regulations
 Regulations") CANADA CONSOLIDATION CODIFICATION Credit Note and Debit Note Information (GST/ HST) Regulations Règlement sur les renseignements à inclure dans les notes de crédit et les notes de débit (TPS/ TVH) SOR/91-44
CANADA CONSOLIDATION CODIFICATION Credit Note and Debit Note Information (GST/ HST) Regulations Règlement sur les renseignements à inclure dans les notes de crédit et les notes de débit (TPS/ TVH) SOR/91-44
Application Form/ Formulaire de demande
 Application Form/ Formulaire de demande Ecosystem Approaches to Health: Summer Workshop and Field school Approches écosystémiques de la santé: Atelier intensif et stage d été Please submit your application
Application Form/ Formulaire de demande Ecosystem Approaches to Health: Summer Workshop and Field school Approches écosystémiques de la santé: Atelier intensif et stage d été Please submit your application
La lettre. Le Dossier. Neurosciences et clinique Je t aime, moi non plus. Internes en médecine ou pharmacie : les années-recherche
 O T O La lettre Automne - Hiver 1998 Le Dossier Neurosciences et clinique Je t aime, moi non plus Internes en médecine ou pharmacie : les années-recherche MD-PhD : l expérience américaine Photographie
O T O La lettre Automne - Hiver 1998 Le Dossier Neurosciences et clinique Je t aime, moi non plus Internes en médecine ou pharmacie : les années-recherche MD-PhD : l expérience américaine Photographie
Jean Sykes Nereus, la collaboration européenne, et le libre accès
 Jean Sykes Nereus, la collaboration européenne, et le libre accès Keynote Item Original citation: Originally presented at UNESCO DKN project steering group meeting, 29 June 2006, Paris, France [Name of
Jean Sykes Nereus, la collaboration européenne, et le libre accès Keynote Item Original citation: Originally presented at UNESCO DKN project steering group meeting, 29 June 2006, Paris, France [Name of
PROBABILITES ET STATISTIQUE I&II
 PROBABILITES ET STATISTIQUE I&II TABLE DES MATIERES CHAPITRE I - COMBINATOIRE ELEMENTAIRE I.1. Rappel des notations de la théorie des ensemble I.1.a. Ensembles et sous-ensembles I.1.b. Diagrammes (dits
PROBABILITES ET STATISTIQUE I&II TABLE DES MATIERES CHAPITRE I - COMBINATOIRE ELEMENTAIRE I.1. Rappel des notations de la théorie des ensemble I.1.a. Ensembles et sous-ensembles I.1.b. Diagrammes (dits
Archived Content. Contenu archivé
 ARCHIVED - Archiving Content ARCHIVÉE - Contenu archivé Archived Content Contenu archivé Information identified as archived is provided for reference, research or recordkeeping purposes. It is not subject
ARCHIVED - Archiving Content ARCHIVÉE - Contenu archivé Archived Content Contenu archivé Information identified as archived is provided for reference, research or recordkeeping purposes. It is not subject
First Nations Assessment Inspection Regulations. Règlement sur l inspection aux fins d évaluation foncière des premières nations CONSOLIDATION
 CANADA CONSOLIDATION CODIFICATION First Nations Assessment Inspection Regulations Règlement sur l inspection aux fins d évaluation foncière des premières nations SOR/2007-242 DORS/2007-242 Current to September
CANADA CONSOLIDATION CODIFICATION First Nations Assessment Inspection Regulations Règlement sur l inspection aux fins d évaluation foncière des premières nations SOR/2007-242 DORS/2007-242 Current to September
AMINES BIOGENIQUES. Dopamine/Noradrénaline/Adrénaline (CATECHOLAMINES) Sérotonine/Histamine/Dopamine
 Sérotonine/Histamine/Dopamine") AMINES BIOGENIQUES Dopamine/Noradrénaline/Adrénaline (CATECHOLAMINES) Sérotonine/Histamine/Dopamine Effets physiologiques des bioamines via le systeme nerveux autonome Cardiovasculaire: Augmentation du
AMINES BIOGENIQUES Dopamine/Noradrénaline/Adrénaline (CATECHOLAMINES) Sérotonine/Histamine/Dopamine Effets physiologiques des bioamines via le systeme nerveux autonome Cardiovasculaire: Augmentation du
Once the installation is complete, you can delete the temporary Zip files..
 Sommaire Installation... 2 After the download... 2 From a CD... 2 Access codes... 2 DirectX Compatibility... 2 Using the program... 2 Structure... 4 Lier une structure à une autre... 4 Personnaliser une
Sommaire Installation... 2 After the download... 2 From a CD... 2 Access codes... 2 DirectX Compatibility... 2 Using the program... 2 Structure... 4 Lier une structure à une autre... 4 Personnaliser une
Guide de rédaction d un protocole de recherche clinique à. l intention des chercheurs évoluant en recherche fondamentale
 V E R S I O N A V R I L 2 0 1 2 C E N T R E D E R E C H E R C H E C L I N I Q U E É T I E N N E - L E B E L D U C H U S Guide de rédaction d un protocole de recherche clinique à l intention des chercheurs
V E R S I O N A V R I L 2 0 1 2 C E N T R E D E R E C H E R C H E C L I N I Q U E É T I E N N E - L E B E L D U C H U S Guide de rédaction d un protocole de recherche clinique à l intention des chercheurs
L ELECTRICITE : SON IMPLICATION DANS LE FONCTIONNEMENT DU CERVEAU ET DANS LES SOINS
 LES RENDEZ-VOUS PRESSE DU NEURODON L ELECTRICITE : SON IMPLICATION DANS LE FONCTIONNEMENT DU CERVEAU ET DANS LES SOINS Dossier réalisé avec l aide des : Professeur Yehezkel BEN ARI Directeur de l INMED
LES RENDEZ-VOUS PRESSE DU NEURODON L ELECTRICITE : SON IMPLICATION DANS LE FONCTIONNEMENT DU CERVEAU ET DANS LES SOINS Dossier réalisé avec l aide des : Professeur Yehezkel BEN ARI Directeur de l INMED
1ST2S Biophysiopathologie : Motricité et système nerveux. 2012-2013. La physiologie neuro-musculaire :
 La physiologie neuro-musculaire : Introduction : Le neurone possède 3 propriétés indispensables pour assurer sa fonction au sein de l organisme : -l excitabilité : capacité à répondre à un stimulus -la
La physiologie neuro-musculaire : Introduction : Le neurone possède 3 propriétés indispensables pour assurer sa fonction au sein de l organisme : -l excitabilité : capacité à répondre à un stimulus -la
Form of Deeds Relating to Certain Successions of Cree and Naskapi Beneficiaries Regulations
 CANADA CONSOLIDATION CODIFICATION Form of Deeds Relating to Certain Successions of Cree and Naskapi Beneficiaries Regulations Règlement sur la forme des actes relatifs à certaines successions de bénéficiaires
CANADA CONSOLIDATION CODIFICATION Form of Deeds Relating to Certain Successions of Cree and Naskapi Beneficiaries Regulations Règlement sur la forme des actes relatifs à certaines successions de bénéficiaires
Quick Start Guide This guide is intended to get you started with Rational ClearCase or Rational ClearCase MultiSite.
 Rational ClearCase or ClearCase MultiSite Version 7.0.1 Quick Start Guide This guide is intended to get you started with Rational ClearCase or Rational ClearCase MultiSite. Product Overview IBM Rational
Rational ClearCase or ClearCase MultiSite Version 7.0.1 Quick Start Guide This guide is intended to get you started with Rational ClearCase or Rational ClearCase MultiSite. Product Overview IBM Rational
Cheque Holding Policy Disclosure (Banks) Regulations. Règlement sur la communication de la politique de retenue de chèques (banques) CONSOLIDATION
 Regulations. Règlement sur la communication de la politique de retenue de chèques (banques) CONSOLIDATION") CANADA CONSOLIDATION CODIFICATION Cheque Holding Policy Disclosure (Banks) Regulations Règlement sur la communication de la politique de retenue de chèques (banques) SOR/2002-39 DORS/2002-39 Current to
CANADA CONSOLIDATION CODIFICATION Cheque Holding Policy Disclosure (Banks) Regulations Règlement sur la communication de la politique de retenue de chèques (banques) SOR/2002-39 DORS/2002-39 Current to
PHOTO ROYAUME DE BELGIQUE /KINDOM OF BELGIUM /KONINKRIJK BELGIE. Données personnelles / personal data
 1 ROYAUME DE BELGIQUE /KINDOM OF BELGIUM /KONINKRIJK BELGIE Service Public Fédéral Affaires Etrangères, Commerce et Coopération au développement Federal Public Service Foreign Affairs, External Trade and
1 ROYAUME DE BELGIQUE /KINDOM OF BELGIUM /KONINKRIJK BELGIE Service Public Fédéral Affaires Etrangères, Commerce et Coopération au développement Federal Public Service Foreign Affairs, External Trade and
Programme d entrainement aux processus temporels chez des enfants présentant un trouble du déficit de l attention avec ou sans hyperactivité (TDA/H)
") UNIVERSITÉ PAUL SABATIER FACULTÉ DE MÉDECINE TOULOUSE RANGUEIL INSTITUT DE FORMATION EN PSYCHOMOTRICITÉ Programme d entrainement aux processus temporels chez des enfants présentant un trouble du déficit
UNIVERSITÉ PAUL SABATIER FACULTÉ DE MÉDECINE TOULOUSE RANGUEIL INSTITUT DE FORMATION EN PSYCHOMOTRICITÉ Programme d entrainement aux processus temporels chez des enfants présentant un trouble du déficit
RETHINKING JACQUES ELLUL AND THE TECHNOLOGICAL SOCIETY IN THE 21ST CENTURY REPENSER JACQUES ELLUL ET LA SOCIETE TECHNICIENNE AU 21EME SIECLE
 CALL FOR PAPERS / APPEL A COMMUNICATIONS RETHINKING JACQUES ELLUL AND THE TECHNOLOGICAL SOCIETY IN THE 21ST CENTURY REPENSER JACQUES ELLUL ET LA SOCIETE TECHNICIENNE AU 21EME SIECLE The Conference Rethinking
CALL FOR PAPERS / APPEL A COMMUNICATIONS RETHINKING JACQUES ELLUL AND THE TECHNOLOGICAL SOCIETY IN THE 21ST CENTURY REPENSER JACQUES ELLUL ET LA SOCIETE TECHNICIENNE AU 21EME SIECLE The Conference Rethinking
Photoactivatable Probes for Protein Labeling
 Photoactivatable Probes for Protein Labeling THÈSE N O 4660 (2010) PRÉSENTÉE LE 26 MARS 2010 À LA FACULTÉ SCIENCES DE BASE LABORATOIRE D'INGÉNIERIE DES PROTÉINES PROGRAMME DOCTORAL EN CHIMIE ET GÉNIE CHIMIQUE
Photoactivatable Probes for Protein Labeling THÈSE N O 4660 (2010) PRÉSENTÉE LE 26 MARS 2010 À LA FACULTÉ SCIENCES DE BASE LABORATOIRE D'INGÉNIERIE DES PROTÉINES PROGRAMME DOCTORAL EN CHIMIE ET GÉNIE CHIMIQUE
APPENDIX 2. Provisions to be included in the contract between the Provider and the. Holder
 Page 1 APPENDIX 2 Provisions to be included in the contract between the Provider and the Obligations and rights of the Applicant / Holder Holder 1. The Applicant or Licensee acknowledges that it has read
Page 1 APPENDIX 2 Provisions to be included in the contract between the Provider and the Obligations and rights of the Applicant / Holder Holder 1. The Applicant or Licensee acknowledges that it has read
Support Orders and Support Provisions (Banks and Authorized Foreign Banks) Regulations
 Regulations") CANADA CONSOLIDATION CODIFICATION Support Orders and Support Provisions (Banks and Authorized Foreign Banks) Regulations Règlement sur les ordonnances alimentaires et les dispositions alimentaires (banques
CANADA CONSOLIDATION CODIFICATION Support Orders and Support Provisions (Banks and Authorized Foreign Banks) Regulations Règlement sur les ordonnances alimentaires et les dispositions alimentaires (banques
UNIVERSITE LYON 3 (JEAN MOULIN) Référence GALAXIE : 4140
 Référence GALAXIE : 4140") UNIVERSITE LYON 3 (JEAN MOULIN) Référence GALAXIE : 4140 Numéro dans le SI local : Référence GESUP : 0202 Corps : Professeur des universités Article : 51 Chaire : Non Section 1 : 01-Droit privé et sciences
UNIVERSITE LYON 3 (JEAN MOULIN) Référence GALAXIE : 4140 Numéro dans le SI local : Référence GESUP : 0202 Corps : Professeur des universités Article : 51 Chaire : Non Section 1 : 01-Droit privé et sciences
Règlement relatif à l examen fait conformément à la Déclaration canadienne des droits. Canadian Bill of Rights Examination Regulations CODIFICATION
 CANADA CONSOLIDATION CODIFICATION Canadian Bill of Rights Examination Regulations Règlement relatif à l examen fait conformément à la Déclaration canadienne des droits C.R.C., c. 394 C.R.C., ch. 394 Current
CANADA CONSOLIDATION CODIFICATION Canadian Bill of Rights Examination Regulations Règlement relatif à l examen fait conformément à la Déclaration canadienne des droits C.R.C., c. 394 C.R.C., ch. 394 Current
MEMORANDUM POUR UNE DEMANDE DE BOURSE DE RECHERCHE DOCTORALE DE LA FONDATION MARTINE AUBLET
 MEMORANDUM POUR UNE DEMANDE DE BOURSE DE RECHERCHE DOCTORALE DE LA FONDATION MARTINE AUBLET ATTENTION : Tout formulaire de candidature qui présentera des erreurs ou anomalies dans le remplissage des champs
MEMORANDUM POUR UNE DEMANDE DE BOURSE DE RECHERCHE DOCTORALE DE LA FONDATION MARTINE AUBLET ATTENTION : Tout formulaire de candidature qui présentera des erreurs ou anomalies dans le remplissage des champs
Bourses d excellence pour les masters orientés vers la recherche
 Masters de Mathématiques à l'université Lille 1 Mathématiques Ingénierie Mathématique Mathématiques et Finances Bourses d excellence pour les masters orientés vers la recherche Mathématiques appliquées
Masters de Mathématiques à l'université Lille 1 Mathématiques Ingénierie Mathématique Mathématiques et Finances Bourses d excellence pour les masters orientés vers la recherche Mathématiques appliquées
DOCUMENTATION MODULE BLOCKCATEGORIESCUSTOM Module crée par Prestacrea - Version : 2.0
 DOCUMENTATION MODULE BLOCKCATEGORIESCUSTOM Module crée par Prestacrea - Version : 2.0 INDEX : DOCUMENTATION - FRANCAIS... 2 1. INSTALLATION... 2 2. CONFIGURATION... 2 3. LICENCE ET COPYRIGHT... 3 4. MISES
DOCUMENTATION MODULE BLOCKCATEGORIESCUSTOM Module crée par Prestacrea - Version : 2.0 INDEX : DOCUMENTATION - FRANCAIS... 2 1. INSTALLATION... 2 2. CONFIGURATION... 2 3. LICENCE ET COPYRIGHT... 3 4. MISES
F1 Security Requirement Check List (SRCL)
") F1 Security Requirement Check List (SRCL) Liste de vérification des exigences relatives à la sécurité (LVERS) Cyber Protection Supply Arrangement (CPSA) Arrangement en matière d approvisionnement en cyberprotection
F1 Security Requirement Check List (SRCL) Liste de vérification des exigences relatives à la sécurité (LVERS) Cyber Protection Supply Arrangement (CPSA) Arrangement en matière d approvisionnement en cyberprotection
7- Les Antiépileptiques
 7- Les Antiépileptiques 1 Définition L épilepsie est un trouble neurologique chronique caractérisé par la survenue périodique et imprévisible de crises convulsives dues à l émission de décharges électriques
7- Les Antiépileptiques 1 Définition L épilepsie est un trouble neurologique chronique caractérisé par la survenue périodique et imprévisible de crises convulsives dues à l émission de décharges électriques
IPSAS 32 «Service concession arrangements» (SCA) Marie-Pierre Cordier Baudouin Griton, IPSAS Board
 Marie-Pierre Cordier Baudouin Griton, IPSAS Board") IPSAS 32 «Service concession arrangements» (SCA) Marie-Pierre Cordier Baudouin Griton, IPSAS Board 1 L élaboration de la norme IPSAS 32 Objectif : traitement comptable des «service concession arrangements»
IPSAS 32 «Service concession arrangements» (SCA) Marie-Pierre Cordier Baudouin Griton, IPSAS Board 1 L élaboration de la norme IPSAS 32 Objectif : traitement comptable des «service concession arrangements»
RULE 5 - SERVICE OF DOCUMENTS RÈGLE 5 SIGNIFICATION DE DOCUMENTS. Rule 5 / Règle 5
 RULE 5 - SERVICE OF DOCUMENTS General Rules for Manner of Service Notices of Application and Other Documents 5.01 (1) A notice of application or other document may be served personally, or by an alternative
RULE 5 - SERVICE OF DOCUMENTS General Rules for Manner of Service Notices of Application and Other Documents 5.01 (1) A notice of application or other document may be served personally, or by an alternative
Neurologiques gq Centrales EMPR LE NORMANDY GRANVILLE
 La Marche dans les Affections Neurologiques gq Centrales S Injeyan JL Isambert Y Bebin S Le Doze M Cano P Fages W Loisel La Marche Fonction complexe Organisée hiérarchiquement au niveau spinal Contrôlée
La Marche dans les Affections Neurologiques gq Centrales S Injeyan JL Isambert Y Bebin S Le Doze M Cano P Fages W Loisel La Marche Fonction complexe Organisée hiérarchiquement au niveau spinal Contrôlée
MANUEL MARKETING ET SURVIE PDF
 MANUEL MARKETING ET SURVIE PDF ==> Download: MANUEL MARKETING ET SURVIE PDF MANUEL MARKETING ET SURVIE PDF - Are you searching for Manuel Marketing Et Survie Books? Now, you will be happy that at this
MANUEL MARKETING ET SURVIE PDF ==> Download: MANUEL MARKETING ET SURVIE PDF MANUEL MARKETING ET SURVIE PDF - Are you searching for Manuel Marketing Et Survie Books? Now, you will be happy that at this
Notice Technique / Technical Manual
 Contrôle d accès Access control Encodeur USB Mifare ENCOD-USB-AI Notice Technique / Technical Manual SOMMAIRE p.2/10 Sommaire Remerciements... 3 Informations et recommandations... 4 Caractéristiques techniques...
Contrôle d accès Access control Encodeur USB Mifare ENCOD-USB-AI Notice Technique / Technical Manual SOMMAIRE p.2/10 Sommaire Remerciements... 3 Informations et recommandations... 4 Caractéristiques techniques...
English Q&A #1 Braille Services Requirement PPTC 144918. Q1. Would you like our proposal to be shipped or do you prefer an electronic submission?
 English Q&A #1 Braille Services Requirement PPTC 144918 Q1. Would you like our proposal to be shipped or do you prefer an electronic submission? A1. Passport Canada requests that bidders provide their
English Q&A #1 Braille Services Requirement PPTC 144918 Q1. Would you like our proposal to be shipped or do you prefer an electronic submission? A1. Passport Canada requests that bidders provide their
Short-term Pooled Investment Fund Regulations. Règlement sur le fonds commun de placement à court terme CONSOLIDATION CODIFICATION
 CANADA CONSOLIDATION CODIFICATION Short-term Pooled Investment Fund Regulations Règlement sur le fonds commun de placement à court terme SOR/2006-245 DORS/2006-245 Current to September 27, 2015 À jour
CANADA CONSOLIDATION CODIFICATION Short-term Pooled Investment Fund Regulations Règlement sur le fonds commun de placement à court terme SOR/2006-245 DORS/2006-245 Current to September 27, 2015 À jour
Thierry DELZESCAUX. «biopicsel» group, URA CNRS-CEA 2210 Service MIRCen, I²BM, CEA Fontenay-aux-Roses, France. Thierry.Delzescaux@cea.
 Thierry DELZESCAUX «biopicsel» group, URA CNRS-CEA 2210 Service MIRCen, I²BM, CEA Fontenay-aux-Roses, France Thierry.Delzescaux@cea.fr Thierry DELZESCAUX Ingénieur chercheur CEA Experience Since 2008:
Thierry DELZESCAUX «biopicsel» group, URA CNRS-CEA 2210 Service MIRCen, I²BM, CEA Fontenay-aux-Roses, France Thierry.Delzescaux@cea.fr Thierry DELZESCAUX Ingénieur chercheur CEA Experience Since 2008:
8. Cours virtuel Enjeux nordiques / Online Class Northern Issues Formulaire de demande de bourse / Fellowship Application Form
 F-8a-v1 1 / 7 8. Cours virtuel Enjeux nordiques / Online Class Northern Issues Formulaire de demande de bourse / Fellowship Application Form Nom de famille du candidat Langue de correspondance Français
F-8a-v1 1 / 7 8. Cours virtuel Enjeux nordiques / Online Class Northern Issues Formulaire de demande de bourse / Fellowship Application Form Nom de famille du candidat Langue de correspondance Français
Life Companies Borrowing Regulations. Règlement sur les emprunts des sociétés d assurance-vie CONSOLIDATION CODIFICATION
 CANADA CONSOLIDATION CODIFICATION Life Companies Borrowing Regulations Règlement sur les emprunts des sociétés d assurance-vie SOR/92-277 DORS/92-277 Current to August 4, 2015 À jour au 4 août 2015 Published
CANADA CONSOLIDATION CODIFICATION Life Companies Borrowing Regulations Règlement sur les emprunts des sociétés d assurance-vie SOR/92-277 DORS/92-277 Current to August 4, 2015 À jour au 4 août 2015 Published
Présentation Générale
 Mars 2009 Présentation Générale 1- Le Master Recherche en Sciences de la Vie et de la Santé à Nice Objectifs, environnement scientifique, organisation Enseignements, les spécialités, les cours et les stages
Mars 2009 Présentation Générale 1- Le Master Recherche en Sciences de la Vie et de la Santé à Nice Objectifs, environnement scientifique, organisation Enseignements, les spécialités, les cours et les stages
Loi sur l aide financière à la Banque Commerciale du Canada. Canadian Commercial Bank Financial Assistance Act CODIFICATION CONSOLIDATION
 CANADA CONSOLIDATION CODIFICATION Canadian Commercial Bank Financial Assistance Act Loi sur l aide financière à la Banque Commerciale du Canada S.C. 1985, c. 9 S.C. 1985, ch. 9 Current to September 10,
CANADA CONSOLIDATION CODIFICATION Canadian Commercial Bank Financial Assistance Act Loi sur l aide financière à la Banque Commerciale du Canada S.C. 1985, c. 9 S.C. 1985, ch. 9 Current to September 10,
Physiopathologie : de la Molécule à l'homme
 Mention Sciences du Vivant Spécialité de Master : Physiopathologie : de la Molécule à l'homme Pourquoi, comment, combien, contourner, greffer, restaurer, inhiber, suppléer Responsables : Dr Gilles Prévost
Mention Sciences du Vivant Spécialité de Master : Physiopathologie : de la Molécule à l'homme Pourquoi, comment, combien, contourner, greffer, restaurer, inhiber, suppléer Responsables : Dr Gilles Prévost
SCHOLARSHIP ANSTO FRENCH EMBASSY (SAFE) PROGRAM 2015-2 APPLICATION FORM
 PROGRAM 2015-2 APPLICATION FORM") SCHOLARSHIP ANSTO FRENCH EMBASSY (SAFE) PROGRAM 2015-2 APPLICATION FORM APPLICATION FORM / FORMULAIRE DE CANDIDATURE Note: If there is insufficient space to answer a question, please attach additional
SCHOLARSHIP ANSTO FRENCH EMBASSY (SAFE) PROGRAM 2015-2 APPLICATION FORM APPLICATION FORM / FORMULAIRE DE CANDIDATURE Note: If there is insufficient space to answer a question, please attach additional
Material Banking Group Percentage Regulations. Règlement fixant le pourcentage (groupe bancaire important) CONSOLIDATION CODIFICATION
 CONSOLIDATION CODIFICATION") CANADA CONSOLIDATION CODIFICATION Material Banking Group Percentage Regulations Règlement fixant le pourcentage (groupe bancaire important) SOR/2008-163 DORS/2008-163 Current to August 30, 2015 À jour
CANADA CONSOLIDATION CODIFICATION Material Banking Group Percentage Regulations Règlement fixant le pourcentage (groupe bancaire important) SOR/2008-163 DORS/2008-163 Current to August 30, 2015 À jour
SERVEUR DÉDIÉ DOCUMENTATION
 SERVEUR DÉDIÉ DOCUMENTATION Release 5.0.6.0 19 Juillet 2013 Copyright 2013 GIANTS Software GmbH, All Rights Reserved. 1/9 CHANGE LOG Correction de bug divers (5.0.6.0) Ajout d une option de relance automatique
SERVEUR DÉDIÉ DOCUMENTATION Release 5.0.6.0 19 Juillet 2013 Copyright 2013 GIANTS Software GmbH, All Rights Reserved. 1/9 CHANGE LOG Correction de bug divers (5.0.6.0) Ajout d une option de relance automatique
Comment Accéder à des Bases de Données MySQL avec Windows lorqu'elles sont sur un Serveur Linux
 Comment Accéder à des Bases de Données MySQL avec Windows lorqu'elles sont sur un Serveur Linux Distribué par Le Projet Documentation d'openoffice.org Table of Contents 1) Préambule...3 2) Télécharger
Comment Accéder à des Bases de Données MySQL avec Windows lorqu'elles sont sur un Serveur Linux Distribué par Le Projet Documentation d'openoffice.org Table of Contents 1) Préambule...3 2) Télécharger
Université de XY University of XY. Faculté XY Faculty of XY
 Université de XY University of XY Faculté XY Faculty of XY Le présent supplément au diplôme suit le modèle élaboré par la Commission européenne, le Conseil d'europe et l'unesco/cepes. Le supplément vise
Université de XY University of XY Faculté XY Faculty of XY Le présent supplément au diplôme suit le modèle élaboré par la Commission européenne, le Conseil d'europe et l'unesco/cepes. Le supplément vise
CHAPITRE 3 LA SYNTHESE DES PROTEINES
 CHAITRE 3 LA SYNTHESE DES ROTEINES On sait qu un gène détient dans sa séquence nucléotidique, l information permettant la synthèse d un polypeptide. Ce dernier caractérisé par sa séquence d acides aminés
CHAITRE 3 LA SYNTHESE DES ROTEINES On sait qu un gène détient dans sa séquence nucléotidique, l information permettant la synthèse d un polypeptide. Ce dernier caractérisé par sa séquence d acides aminés
ICA Congress, Brisbane 2012 Thème général : Les temps qui changent. La confiance et les archives*
 ICA Congress, Brisbane 2012 Thème général : Les temps qui changent. La confiance et les archives* Titre : Un résumé de la pratique archivistique internationale adaptée au niveau local : manuel pratique
ICA Congress, Brisbane 2012 Thème général : Les temps qui changent. La confiance et les archives* Titre : Un résumé de la pratique archivistique internationale adaptée au niveau local : manuel pratique
Sommeil et sport Dr. Arnaud PRIGENT (Pneumologue à St LAURENT) sport et sommeil 01/06/2010
 sport et sommeil 01/06/2010") Sommeil et sport Structure du sommeil Au cours du sommeil, on repère 2 principaux types de sommeil : Le sommeil lent. Le sommeil paradoxal. Processus de régulation 3 processus: - circadienne (processus
Sommeil et sport Structure du sommeil Au cours du sommeil, on repère 2 principaux types de sommeil : Le sommeil lent. Le sommeil paradoxal. Processus de régulation 3 processus: - circadienne (processus
Paxton. ins-20605. Net2 desktop reader USB
 Paxton ins-20605 Net2 desktop reader USB 1 3 2 4 1 2 Desktop Reader The desktop reader is designed to sit next to the PC. It is used for adding tokens to a Net2 system and also for identifying lost cards.
Paxton ins-20605 Net2 desktop reader USB 1 3 2 4 1 2 Desktop Reader The desktop reader is designed to sit next to the PC. It is used for adding tokens to a Net2 system and also for identifying lost cards.
Cedric Dumoulin (C) The Java EE 7 Tutorial http://docs.oracle.com/javaee/7/tutorial/doc/
 The Java EE 7 Tutorial http://docs.oracle.com/javaee/7/tutorial/doc/") Cedric Dumoulin (C) The Java EE 7 Tutorial http://docs.oracle.com/javaee/7/tutorial/doc/ Webographie The Java EE 7 Tutorial http://docs.oracle.com/javaee/7/tutorial/doc/ Les slides de cette présentation
Cedric Dumoulin (C) The Java EE 7 Tutorial http://docs.oracle.com/javaee/7/tutorial/doc/ Webographie The Java EE 7 Tutorial http://docs.oracle.com/javaee/7/tutorial/doc/ Les slides de cette présentation
Air Transportation Tax Order, 1995. Décret de 1995 sur la taxe de transport aérien CONSOLIDATION CODIFICATION
 CANADA CONSOLIDATION CODIFICATION Air Transportation Tax Order, 1995 Décret de 1995 sur la taxe de transport aérien SOR/95-206 DORS/95-206 Current to August 30, 2015 À jour au 30 août 2015 Published by
CANADA CONSOLIDATION CODIFICATION Air Transportation Tax Order, 1995 Décret de 1995 sur la taxe de transport aérien SOR/95-206 DORS/95-206 Current to August 30, 2015 À jour au 30 août 2015 Published by
Hospices cantonaux Centre Hospitalier Universitaire Vaudois DOSSIER DE PRESSE. Création du Centre romand hospitalo-universitaire de neurochirurgie
 Hospices cantonaux Centre Hospitalier Universitaire Vaudois DOSSIER DE PRESSE Création du Centre romand hospitalo-universitaire de neurochirurgie 1. Le Centre romand hospitalo-universitaire de neurochirurgie
Hospices cantonaux Centre Hospitalier Universitaire Vaudois DOSSIER DE PRESSE Création du Centre romand hospitalo-universitaire de neurochirurgie 1. Le Centre romand hospitalo-universitaire de neurochirurgie
FCM 2015 ANNUAL CONFERENCE AND TRADE SHOW Terms and Conditions for Delegates and Companions Shaw Convention Centre, Edmonton, AB June 5 8, 2015
 FCM 2015 ANNUAL CONFERENCE AND TRADE SHOW Terms and Conditions for Delegates and Companions Shaw Convention Centre, Edmonton, AB June 5 8, 2015 Early-bird registration Early-bird registration ends April
FCM 2015 ANNUAL CONFERENCE AND TRADE SHOW Terms and Conditions for Delegates and Companions Shaw Convention Centre, Edmonton, AB June 5 8, 2015 Early-bird registration Early-bird registration ends April
Sujets présentés par le Professeur Olivier CUSSENOT
 ANAMACaP Association Nationale des Malades du Cancer de la Prostate 17, bis Avenue Poincaré. 57400 SARREBOURG Sujets présentés par le Professeur Olivier CUSSENOT Place des nouvelles techniques d imagerie
ANAMACaP Association Nationale des Malades du Cancer de la Prostate 17, bis Avenue Poincaré. 57400 SARREBOURG Sujets présentés par le Professeur Olivier CUSSENOT Place des nouvelles techniques d imagerie
CEST POUR MIEUX PLACER MES PDF
 CEST POUR MIEUX PLACER MES PDF ==> Download: CEST POUR MIEUX PLACER MES PDF CEST POUR MIEUX PLACER MES PDF - Are you searching for Cest Pour Mieux Placer Mes Books? Now, you will be happy that at this
CEST POUR MIEUX PLACER MES PDF ==> Download: CEST POUR MIEUX PLACER MES PDF CEST POUR MIEUX PLACER MES PDF - Are you searching for Cest Pour Mieux Placer Mes Books? Now, you will be happy that at this
This is a preview - click here to buy the full publication NORME INTERNATIONALE INTERNATIONAL STAN DARD. Telecontrol equipment and systems
 NORME INTERNATIONALE INTERNATIONAL STAN DARD CEI IEC 870-3 Première édition First edition 1989-03 Matériels et systèmes de téléconduite Troisième partie: Interfaces (caractéristiques électriques) Telecontrol
NORME INTERNATIONALE INTERNATIONAL STAN DARD CEI IEC 870-3 Première édition First edition 1989-03 Matériels et systèmes de téléconduite Troisième partie: Interfaces (caractéristiques électriques) Telecontrol
Resident Canadian (Insurance Companies) Regulations. Règlement sur les résidents canadiens (sociétés d assurances) CONSOLIDATION CODIFICATION
 Regulations. Règlement sur les résidents canadiens (sociétés d assurances) CONSOLIDATION CODIFICATION") CANADA CONSOLIDATION CODIFICATION Resident Canadian (Insurance Companies) Regulations Règlement sur les résidents canadiens (sociétés d assurances) SOR/92-284 DORS/92-284 Current to August 4, 2015 À jour
CANADA CONSOLIDATION CODIFICATION Resident Canadian (Insurance Companies) Regulations Règlement sur les résidents canadiens (sociétés d assurances) SOR/92-284 DORS/92-284 Current to August 4, 2015 À jour
Grandes tendances et leurs impacts sur l acquisition de produits et services TI.
 Grandes tendances et leurs impacts sur l acquisition de produits et services TI. François Banville Partenaire exécutif chez Gartner Gartner is a registered trademark of Gartner, Inc. or its affiliates.
Grandes tendances et leurs impacts sur l acquisition de produits et services TI. François Banville Partenaire exécutif chez Gartner Gartner is a registered trademark of Gartner, Inc. or its affiliates.
Insulinothérapie et diabète de type 1
 Insulinothérapie et diabète de type 1 Introduction: la molécule d insuline L instauration de l insulinothérapie Dispositif d administration de l insuline Les propriétés de l insuline Insuline et schémas
Insulinothérapie et diabète de type 1 Introduction: la molécule d insuline L instauration de l insulinothérapie Dispositif d administration de l insuline Les propriétés de l insuline Insuline et schémas
TD de Biochimie 4 : Coloration.
 TD de Biochimie 4 : Coloration. Synthèse de l expérience 2 Les questions posées durant l expérience 2 Exposé sur les méthodes de coloration des molécules : Générique Spécifique Autres Questions Pourquoi
TD de Biochimie 4 : Coloration. Synthèse de l expérience 2 Les questions posées durant l expérience 2 Exposé sur les méthodes de coloration des molécules : Générique Spécifique Autres Questions Pourquoi
Rôle des acides biliaires dans la régulation de l homéostasie du glucose : implication de FXR dans la cellule bêta-pancréatique
 Rôle des acides biliaires dans la régulation de l homéostasie du glucose : implication de FXR dans la cellule bêta-pancréatique Tuteur : Anne Muhr-Tailleux cardiovasculaires et diabète (Equipe 1) Institut
Rôle des acides biliaires dans la régulation de l homéostasie du glucose : implication de FXR dans la cellule bêta-pancréatique Tuteur : Anne Muhr-Tailleux cardiovasculaires et diabète (Equipe 1) Institut
Insérer des images dans Base
 Insérer des images dans Base Version 1.0 du 30.05.2006 Réalisé avec : OOo 2.0.2 Plate-forme / Os : Toutes Distribué par le projet fr.openoffice.org Sommaire 1 Présentation...3 2 Quelques notions initiales...3
Insérer des images dans Base Version 1.0 du 30.05.2006 Réalisé avec : OOo 2.0.2 Plate-forme / Os : Toutes Distribué par le projet fr.openoffice.org Sommaire 1 Présentation...3 2 Quelques notions initiales...3
Formulaire de candidature pour les bourses de mobilité internationale niveau Master/ Application Form for International Master Scholarship Program
 Formulaire de candidature pour les bourses de mobilité internationale niveau Master/ Application Form for International Master Scholarship Program Année universitaire / Academic Year: 2013 2014 INTITULE
Formulaire de candidature pour les bourses de mobilité internationale niveau Master/ Application Form for International Master Scholarship Program Année universitaire / Academic Year: 2013 2014 INTITULE
NORME INTERNATIONALE INTERNATIONAL STANDARD. Dispositifs à semiconducteurs Dispositifs discrets. Semiconductor devices Discrete devices
 NORME INTERNATIONALE INTERNATIONAL STANDARD CEI IEC 747-6-3 QC 750113 Première édition First edition 1993-11 Dispositifs à semiconducteurs Dispositifs discrets Partie 6: Thyristors Section trois Spécification
NORME INTERNATIONALE INTERNATIONAL STANDARD CEI IEC 747-6-3 QC 750113 Première édition First edition 1993-11 Dispositifs à semiconducteurs Dispositifs discrets Partie 6: Thyristors Section trois Spécification
Disclosure on Account Opening by Telephone Request (Trust and Loan Companies) Regulations
 Regulations") CANADA CONSOLIDATION CODIFICATION Disclosure on Account Opening by Telephone Request (Trust and Loan Companies) Regulations Règlement sur la communication en cas de demande téléphonique d ouverture de
CANADA CONSOLIDATION CODIFICATION Disclosure on Account Opening by Telephone Request (Trust and Loan Companies) Regulations Règlement sur la communication en cas de demande téléphonique d ouverture de
e-science : perspectives et opportunités pour de nouvelles pratiques de la recherche en informatique et mathématiques appliquées
 Emilie MANON, Joanna JANIK, Gabrielle FELTIN e-science : perspectives et opportunités pour de nouvelles pratiques de la recherche en informatique et mathématiques appliquées 1 Introduction : La recherche
Emilie MANON, Joanna JANIK, Gabrielle FELTIN e-science : perspectives et opportunités pour de nouvelles pratiques de la recherche en informatique et mathématiques appliquées 1 Introduction : La recherche
en SCÈNE RATIONAL Rational Démonstration SDP : automatisation de la chaîne de développement Samira BATAOUCHE sbataouche@fr.ibm.com
 Rational Démonstration SDP : automatisation de la chaîne de développement Samira BATAOUCHE sbataouche@fr.ibm.com Fabrice GRELIER fabrice.grelier@fr.ibm.com RATIONAL en SCÈNE 2007 IBM Corporation Objectif
Rational Démonstration SDP : automatisation de la chaîne de développement Samira BATAOUCHE sbataouche@fr.ibm.com Fabrice GRELIER fabrice.grelier@fr.ibm.com RATIONAL en SCÈNE 2007 IBM Corporation Objectif
COMMISSION NATIONALE D EVALUATION DES DISPOSITIFS MEDICAUX ET DES TECHNOLOGIES DE SANTE. AVIS DE LA COMMISSION 08 février 2011 CONCLUSIONS
 COMMISSION NATIONALE D EVALUATION DES DISPOSITIFS MEDICAUX ET DES TECHNOLOGIES DE SANTE AVIS DE LA COMMISSION 08 février 2011 Nom : Modèles et références retenus : Fabricant : Demandeur : Données disponibles
COMMISSION NATIONALE D EVALUATION DES DISPOSITIFS MEDICAUX ET DES TECHNOLOGIES DE SANTE AVIS DE LA COMMISSION 08 février 2011 Nom : Modèles et références retenus : Fabricant : Demandeur : Données disponibles
calls.paris-neuroscience.fr Tutoriel pour Candidatures en ligne *** Online Applications Tutorial
 calls.paris-neuroscience.fr Tutoriel pour Candidatures en ligne Online Applications Tutorial 1/4 Pour postuler aux Appels d Offres de l ENP, vous devez aller sur la plateforme : calls.parisneuroscience.fr.
calls.paris-neuroscience.fr Tutoriel pour Candidatures en ligne Online Applications Tutorial 1/4 Pour postuler aux Appels d Offres de l ENP, vous devez aller sur la plateforme : calls.parisneuroscience.fr.
http://www.neur-one.fr/ Cerveau & Psycho - N 28
 1 Cerveau & Psycho - N 28 Les souvenirs sont gravés dans la mémoire sous forme de combinaisons particulières de modifications de synapses, les jonctions entre neurones. Ces modifications doivent être consolidées,
1 Cerveau & Psycho - N 28 Les souvenirs sont gravés dans la mémoire sous forme de combinaisons particulières de modifications de synapses, les jonctions entre neurones. Ces modifications doivent être consolidées,
Association les DAUPHINS. Club des Dauphins. Régate des Bancs de Flandre 2015. Dunkerque 19 & 20 septembre 2015. Avis de course
 2015 Club des Dauphins Régate des Bancs de Flandre 2015 Dunkerque 19 & 20 septembre 2015 Avis de course www.dauphinsdk.org / www.lesbancsdeflandre.free.fr AVIS DE COURSE TYPE HABITABLES 2013-2016 Nom de
2015 Club des Dauphins Régate des Bancs de Flandre 2015 Dunkerque 19 & 20 septembre 2015 Avis de course www.dauphinsdk.org / www.lesbancsdeflandre.free.fr AVIS DE COURSE TYPE HABITABLES 2013-2016 Nom de
IMMUNOLOGIE. La spécificité des immunoglobulines et des récepteurs T. Informations scientifiques
 IMMUNOLOGIE La spécificité des immunoglobulines et des récepteurs T Informations scientifiques L infection par le VIH entraîne des réactions immunitaires de l organisme qui se traduisent par la production
IMMUNOLOGIE La spécificité des immunoglobulines et des récepteurs T Informations scientifiques L infection par le VIH entraîne des réactions immunitaires de l organisme qui se traduisent par la production
COPYRIGHT Danish Standards. NOT FOR COMMERCIAL USE OR REPRODUCTION. DS/EN 61303:1997
 COPYRIGHT Danish Standards. NOT FOR COMMERCIAL USE OR REPRODUCTION. DS/EN 61303:1997 COPYRIGHT Danish Standards. NOT FOR COMMERCIAL USE OR REPRODUCTION. DS/EN 61303:1997 COPYRIGHT Danish Standards. NOT
COPYRIGHT Danish Standards. NOT FOR COMMERCIAL USE OR REPRODUCTION. DS/EN 61303:1997 COPYRIGHT Danish Standards. NOT FOR COMMERCIAL USE OR REPRODUCTION. DS/EN 61303:1997 COPYRIGHT Danish Standards. NOT
Le Tour de Bretagne à la Voile est une épreuve de catégorie 3 des RSO. En cas de traduction de cet avis de course, le texte français prévaudra
 AVIS DE COURSE Organisation : Ligue Bretagne de Voile 1 rue de Kerbriant 29200 Brest Tél : 02 98 02 83 46 Fax : 02 98 02 83 40 info@voile bretagne.com http://www.voile bretagne.com/ et http://www.tourdebretagnealavoile.com/
AVIS DE COURSE Organisation : Ligue Bretagne de Voile 1 rue de Kerbriant 29200 Brest Tél : 02 98 02 83 46 Fax : 02 98 02 83 40 info@voile bretagne.com http://www.voile bretagne.com/ et http://www.tourdebretagnealavoile.com/
Borrowing (Property and Casualty Companies and Marine Companies) Regulations
 Regulations") CANADA CONSOLIDATION CODIFICATION Borrowing (Property and Casualty Companies and Marine Companies) Regulations Règlement sur les emprunts des sociétés d assurances multirisques et des sociétés d assurance
CANADA CONSOLIDATION CODIFICATION Borrowing (Property and Casualty Companies and Marine Companies) Regulations Règlement sur les emprunts des sociétés d assurances multirisques et des sociétés d assurance
An Act to Amend the Tobacco Sales Act. Loi modifiant la Loi sur les ventes de tabac CHAPTER 46 CHAPITRE 46
 2015 CHAPTER 46 CHAPITRE 46 An Act to Amend the Tobacco Sales Act Loi modifiant la Loi sur les ventes de tabac Assented to June 5, 2015 Sanctionnée le 5 juin 2015 Her Majesty, by and with the advice and
2015 CHAPTER 46 CHAPITRE 46 An Act to Amend the Tobacco Sales Act Loi modifiant la Loi sur les ventes de tabac Assented to June 5, 2015 Sanctionnée le 5 juin 2015 Her Majesty, by and with the advice and
Railway Operating Certificate Regulations. Règlement sur les certificats d exploitation de chemin de fer CODIFICATION CONSOLIDATION
 CANADA CONSOLIDATION CODIFICATION Railway Operating Certificate Regulations Règlement sur les certificats d exploitation de chemin de fer SOR/2014-258 DORS/2014-258 Current to September 10, 2015 À jour
CANADA CONSOLIDATION CODIFICATION Railway Operating Certificate Regulations Règlement sur les certificats d exploitation de chemin de fer SOR/2014-258 DORS/2014-258 Current to September 10, 2015 À jour
Public and European Business Law - Droit public et européen des affaires. Master I Law Level
 Public and European Business Law - Droit public et européen des affaires Stéphane de La Rosa Master I Law Level Delivered Lectures Jean Monnet Chair «Droit de l Union Européenne et Mutations de l intégration
Public and European Business Law - Droit public et européen des affaires Stéphane de La Rosa Master I Law Level Delivered Lectures Jean Monnet Chair «Droit de l Union Européenne et Mutations de l intégration
Les tests génétiques à des fins médicales
 Les tests génétiques à des fins médicales Les tests génétiques à des fins médicales Nous avons tous hérité d une combinaison unique de gènes de la part de nos parents. Cette constitution originale et l
Les tests génétiques à des fins médicales Les tests génétiques à des fins médicales Nous avons tous hérité d une combinaison unique de gènes de la part de nos parents. Cette constitution originale et l
INSTRUCTIONS. Comment compléter le formulaire. How to complete this form. Instructions
 Instructions INSTRUCTIONS Objet du formulaire Ce formulaire vise à documenter l équivalence de stage d un candidat au permis de géologue par l obtention de diplômes de cycle supérieur. Une déclaration
Instructions INSTRUCTIONS Objet du formulaire Ce formulaire vise à documenter l équivalence de stage d un candidat au permis de géologue par l obtention de diplômes de cycle supérieur. Une déclaration
AIDE FINANCIÈRE POUR ATHLÈTES FINANCIAL ASSISTANCE FOR ATHLETES
 AIDE FINANCIÈRE POUR ATHLÈTES FINANCIAL ASSISTANCE FOR ATHLETES FORMULAIRE DE DEMANDE D AIDE / APPLICATION FORM Espace réservé pour l utilisation de la fondation This space reserved for foundation use
AIDE FINANCIÈRE POUR ATHLÈTES FINANCIAL ASSISTANCE FOR ATHLETES FORMULAIRE DE DEMANDE D AIDE / APPLICATION FORM Espace réservé pour l utilisation de la fondation This space reserved for foundation use
Les lières. MSc in Electronics and Information Technology Engineering. Ingénieur civil. en informatique. MSc in Architectural Engineering
 Ingénieur civil Ingénieur civil Les lières MSc in Electronics and Information Technology Engineering MSc in Architectural Engineering MSc in Civil Engineering MSc in Electromechanical Engineering MSc
Ingénieur civil Ingénieur civil Les lières MSc in Electronics and Information Technology Engineering MSc in Architectural Engineering MSc in Civil Engineering MSc in Electromechanical Engineering MSc
Interest Rate for Customs Purposes Regulations. Règlement sur le taux d intérêt aux fins des douanes CONSOLIDATION CODIFICATION
 CANADA CONSOLIDATION CODIFICATION Interest Rate for Customs Purposes Regulations Règlement sur le taux d intérêt aux fins des douanes SOR/86-1121 DORS/86-1121 Current to August 4, 2015 À jour au 4 août
CANADA CONSOLIDATION CODIFICATION Interest Rate for Customs Purposes Regulations Règlement sur le taux d intérêt aux fins des douanes SOR/86-1121 DORS/86-1121 Current to August 4, 2015 À jour au 4 août
Base de données bibliographiques Pubmed-Medline
 Chapitre 1 ; Domaine 1 ; Documentation ; Champs référentiels 1.1.1, 1.1.2 et 1.1.3 Base de données bibliographiques Pubmed-Medline D r Patrick Deschamps,, 30 mai 2007 PLAN C2i métiers de la santé Introduction
Chapitre 1 ; Domaine 1 ; Documentation ; Champs référentiels 1.1.1, 1.1.2 et 1.1.3 Base de données bibliographiques Pubmed-Medline D r Patrick Deschamps,, 30 mai 2007 PLAN C2i métiers de la santé Introduction