Etude de la régulation du canal CFTR impliqué dans la mucoviscidose par un analogue de la GnRHet le Mg2+
|
|
|
- Bénédicte Boulet
- il y a 6 ans
- Total affichages :
Transcription



1 Etude de la régulation du canal CFTR impliqué dans la mucoviscidose par un analogue de la GnRHet le Mg2+ Marie-Laure Calvez To cite this version: Marie-Laure Calvez. Etude de la régulation du canal CFTR impliqué dans la mucoviscidose par un analogue de la GnRHet le Mg2+. Médecine humaine et pathologie. Université de Bretagne occidentale - Brest, Français. <NNT : 2017BRES0048>. <tel > HAL Id: tel Submitted on 4 Oct 2017 HAL is a multi-disciplinary open access archive for the deposit and dissemination of scientific research documents, whether they are published or not. The documents may come from teaching and research institutions in France or abroad, or from public or private research centers. L archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des établissements d enseignement et de recherche français ou étrangers, des laboratoires publics ou privés.
 Etude de la régulation du canal CFTR, impliqué dans la mucoviscidose, par un analogue de la GnRH et le Mg 2+ présentée par Marie-Laure Calvez Prépa la recherche médicale, UMR 1078, Génétique,")
2 THÈSE / UNIVERSITÉ DE BRETAGNE OCCIDENTALE Bretagne Loire pour obtenir le titre de D Mention : Biologie-Santé École Doctorale "Santé, Information, Communication, Mathématiques, Matière" (ED SICMA 373) Etude de la régulation du canal CFTR, impliqué dans la mucoviscidose, par un analogue de la GnRH et le Mg 2+ présentée par Marie-Laure Calvez Prépa la recherche médicale, UMR 1078, Génétique, Génomique Fonctionnelle et Biotechnologies Thèse soutenue le 13 juin 2017 devant le jury composé de : Christelle Coraux Docteur, Université de Reims Champagne-Ardenne / Rapporteur (Inserm UMR-S 903) Valérie Urbach Docteur, Université Paris Descartes / Rapporteur (Inserm U1151) Pascale Fanen Professeur, Université Créteil Paris-Est Val de Marne / Examinateur (Inserm U955) Tristan Montier Professeur, Université Bretagne Occidentale / Examinateur (Inserm U1078) Claude Férec Professeur, Université Bretagne Occidentale / Examinateur (Inserm U1078) Mathieu Kerbiriou Docteur, Université Bretagne Occidentale / Invité (Inserm U1078) Pascal Trouvé Docteur, Université Bretagne Occidentale / Directeur de thèse (Inserm U1078)
3 1
4 2
5 La mucoviscidose est la maladie héréditaire autosomique récessive, rare, létale, la plus fréquente dans la population caucasienne. Cette maladie est causée par des mutations du gène CFTR (Cystic Fibrosis Transmembrane Conductance Regulator) codant la protéine CFTR. Cette protéine est principalement un canal chlorure (Cl - ) AMPc-dépendant localisé dans la membrane apicale des cellules épithéliales. La mutation F508del entraîne un défaut de maturation de la protéine qui est dégradée. Cependant, une faible quantité de protéines malformées échappe à ce système de contrôle et parvient à la membrane plasmique. Des travaux de notre équipe ont montré une augmentation des efflux ioniques dépendants du CFTR dans des lignées cellulaires épithéliales bronchiques (CFBE41o - ), exprimant le CFTR sauvage ou le CFTR muté F508del, après un traitement par une hormone: la gonadolibérine (GnRH, Gonadotropin releasing hormone, 1h, 10-9 M). Cette augmentation est vraisemblablement due à un nombre plus important de canaux CFTR à la membrane plasmique. objectif de cette thèse a été de tester un analogue de la GnRH comme modulateur, sur des cultures primaires de cellules épithéliales nasales humaines homozygotes pour la mutation F508del. Dans un premier temps, nous avons vérifié la présence du récepteur à la GnRH (R-GnRH) dans notre modèle cellulaire. Puis, nous avons étudié l fonction du CFTR par électrophysiologie. Nous avons observé une augmentation des efflux Cl - médiés par le canal CFTR après un traitement à l (2h, M). Enfin, une étude protéomique protéines différentiellement exprimées après traitement. Certaines protéines mises en évidence pourraient appartenir à des voies de signalisation intracellulaires ayant un rôle dans la régulation de la protéine CFTR et être des cibles thérapeutiques. Par ailleurs, le canal CFTR est régulé par le Mg 2+ intracellulaire ([Mg 2+ ] i). Le canal TRPM7 est le principal régulateur du [Mg 2+ ] i. La [Mg 2+ ] i TRPM7 vérifiée dans des cellules Hela transfectées avec le CFTR sauvage (wt) ou mutés (G551D et F508del). Nous avons étudié la localisation, la fonction et la régulation de TRPM7 dans nos modèles cellulaires avant de rechercher un possible lien fonctionnel entre le CFTR et TRPM7. rmédiaire de la diminution du [Mg 2+ ] i impliquant TRPM7 dans la physiopathologie de la mucoviscidose. CFTR, Mutation F508del, GnRH, cellules nasales humaines, analogue, R-GnRH, Mg 2+, TRPM7 3
6 Cystic fibrosis is the most common lethal autosomal recessive disease in the Caucasian population. This disease is caused by mutations in the gene encoding the CFTR (Cystic Fibrosis Transmembrane conductance Regulator) protein. This protein is a campregulated chloride channel expressed at the apical membrane of epithelial cells. The F508del mutation causes a defect in CFTR protein folding preventing its maturation. Some misfolded proteins escape the control system and reach the plasma membrane. We previously showed a rise of CFTR-dependent ion efflux in wt- and F508del-CFTR human bronchial epithelial expressing cells (CFBE41o - ) after incubation with GnRH (Gonadotropin releasing hormone; 1h, 10-9 M). This increase was probably due to an increased cell surface expression of CFTR. The aim of the present study was to test a GnRH analog as a modulator of CFTR delivery to the plasma membrane and/or activity of CFTR on primary culture of human nasal epithelial cells (F508del/F508del). We checked the GnRH receptor expression in our model. Then, we studied the GnRH effect on CFTR function by electrophysiology. We found a significant increase of CFTR dependent chloride efflux in cells pretreated with analogue (2h, M). Proteomic study enabled us to identify differentially expressed proteins after treatment. Some highlighted proteins could be part of signalling pathways regulating CFTR and could be therapeutic targets. Moreover, CFTR is regulated by intracellular Mg 2+ ([Mg 2+ ] i). TRPM7 (Transient Receptor Potential Melastatin 7) is the main channel regulating [Mg 2+ ] i. [Mg 2+ ] i and TRPM7 expression were measured in Hela cells stably expressing wildtype and two CFTR mutants (F508del-CFTR and G551D-CFTR). We studied TRPM7 expression, function and regulation in our cell models before examining a functional link between TRPM7 and CFTR. In CF models, TRPM7 expression, localization and function are altered. There should be a functional link between TRPM7 and CFTR through the decreased [Mg 2+ ] i involving TRPM7 in cystic fibrosis physiopathology. CFTR, F508del mutation, GnRH, human nasal epithelial cells, analogue, GnRHR, Mg 2+, TRPM7 4
7 I. Généralités...20 II A. épithélium trachéo-bronchique Epithélium de surface...22 a)les cellules ciliées...22 b)les cellules caliciformes...23 c)les cellules basales...23 d)les cellules intermédiaires ou parabasales B C III A. Barrière mécanique...27 B. Transports ioniques transépithéliaux des voies aériennes...29 C. La clairance mucociliaire...30 D. -bactériens...31 E IV. Modèles de culture in vitro...34 A. Cultures primaires de cellules épithéliales respiratoires...35 B. Cultures de lignées cellulaires...37 I. Le canal CFTR : du gène à la protéine...39 A. Le gène CFTR...39 B. Les principaux domaines de la protéine CFTR Les domaines transmembranaires (MSDs) Les domaines de liaison aux nucléotides (NBDs) Le domaine régulateur R
8 II. Voie de biosynthèse de la protéine CFTR...43 A. Biogénèse de la protéine CFTR Assemblage des domaines et glycosylation du CFTR...44 a).. Synthèse et insertion des domaines transmembranaires dans la membrane du RE...44 b)n-glycosylation Contrôle qualité de la conformation des protéines CFTR (ERQC)...45 a)le système de chaperons moléculaires...46 b) 47 B. Transport du CFTR du RE à la membrane plasmique...48 C. Ancrage du CFTR à la membrane plasmique...49 D. Endocytose, recyclage et dégradation...49 III. Activation du CFTR...50 IV. Fonction du canal CFTR...52 A. Canal anionique...52 B. Régulation de canaux ioniques et transporteurs Le canal CFTR et les canaux Cl Le canal CFTR et le canal potassique ROMK Le canal CFTR et le canal ENaC /HCO Le canal CFTR et les aquaporines...55 C. Activateurs/potentiateurs et inhibiteurs du CFTR Activateurs/potentiateurs du CFTR...56 a)liaison directe sur le canal CFTR...56 b)modulation du système de régulation de la protéine CFTR Inhibiteurs du CFTR...57 I. Généralités...59 II. Anomalies moléculaires...60 A. Classes de mutations...60 B. Impact de la mutation F508del sur la protéine CFTR Défaut de repliement Modification des interactions avec les chaperons moléculaires
9 3. La protéine CFTR-F508del à la membrane...64 a)défaut fonctionnel...64 b)système périphérique de contrôle qualité (PQC, peripherical quality control)..64 c)thérapie envisageable pour la mutation F508del...65 III. Manifestations cliniques de la maladie...66 A. Relation génotype/phénotype...66 B. Les différentes atteintes Atteintes respiratoires Atteintes digestives Autres atteintes...69 C. Diagnostic...70 D. CFTR-related disorder (CFTR-RD)...70 IV. Stratégies thérapeutiques...71 A. Traitements disponibles...71 B. La recherche thérapeutique La thérapie génique La thérapie protéique...72 a)suppresseur de codon stop...73 b) Approches alternatives...75 a)activation des canaux Cl b)la cystéamine...75 I. La GnRH...76 A. -hypophysaire...76 B. La GnRH Isoforme de type I (GnRH-I) Isoforme de type II (GnRH-II) Isoforme de type III (GnRH-III)...78 C. Neurones à GnRH...78 D. Régulation de la sécrétion de la GnRH...79 II. Le récepteur de la GnRH (R-GnRH)
10 A. R-GnRH Hypophysaire...79 B. Expression du R-GnRH dans des tissus extra-hypophysaires...81 III. Utilisation de la GnRH synthétique et de ces analogues en thérapeutique humaine..83 A. La GnRH synthétiques et ses analogues...83 B. Les agonistes de la GnRH...83 C. Les antagonistes de la GnRH I. Description...85 II. Fonction et régulation du canal TRPM A. Fonction...86 B. Régulation...87 III. Pathologies associées...88 Article 1.Stimulation de cellules épithéliales bronchiques humaines par la GnRH : CFTR Article 2. Etude de la régulation du canal CFTR impliqué dans la mucoviscidose par un analogue de la GnRH
11 Article 3. La fonction et la régulation de TRPM7 ainsi que la concentration intracellulaire en Mg2+ sont altérées dans les cellules exprimant les mutations CFTR-F508del ou CFTR-G551D
12 Abréviations et sigles A ABC : ATP-binding cassette AC : adénylate cyclase ACTH : Adenocorticotrophic Hormone ADN : acide déoxyribonucléique ADNc : ADN complémentaire ADP : adénosine diphosphate Aha1 : activator of Hsp90 ATPase homologue 1 ALI : interface air-liquide AMPc : adénosine monophosphate cyclique AMPK : -activated protein kinase) AMP-PNP : adenylyl imidodiphosphate ANO1 : anoctamine-1 Anx : annexine (AnxA5 : annexine A5 ; AnxA1 : annexine A1) AP-2 : adaptor protein complex 2 AQP : aquaporines ARN : acide ribonucléique ARNm : ARN messager ASL : liquide de surface des voies aériennes (airway surface liquid) ATF6 : activating transcriptor factor 6 ATP : adénosine triphosphate ATPA : ATP synthase subunit alpha B Bag1 : Bcl2-associated athanogene 1 Br - : ion bromure C Ca 2+ : ion calcium CaCC : calcium (Ca 2+ )-activated chloride channel CAL : CFTR-associated ligand CAP70 : CFTR-associated protein of 70 kda CC10 : Club Cell protein 10 kda CCSP : Club Cell Secretory Protein CCV : clathrin-coated vesicle CCT : chaperoning-containing T-complex 1 CF : cystic fibrosis CFTR : cystic fibrosis transmembrane conductance regulator CFTRinh172 : inhibiteur du CFTR CFTR-RD : CFTR-related disorder CGRP : calcitonin gene-related peptide CHIP : C-terminus of Hsc70-interacting protein CK2 : caseine kinase 2 10
13 Cl - : ion chlorure CNX : calnexine COPI/II : coat protein complex I, II CPX : 8-cyclopentyl-1,3-dipropylxanthine CREB : camp response element binding C-ter : C-terminale Cxs : connexines ChaK1 : channel-kinase 1 DAG : diacyl glycérol ddp : différence de potentiel DPC : Acide diphenylamine-2-carboxylique 2D/3D : bidimensionnel, tridimensionnel E E : Glu (acide glutamique) EC : extracellulaire ECLs : boucles extracellulaires (extra-cellular loops) EDEM : ER degradation-enhancing alpha-mannosidase-like protein ENaC : epithelial sodium (Na+) channel ER (, ) : récepteurs aux oestrogènes (alpha, bêta) (estrogen receptors) ER : endoplasmic reticulum ERQC : contrôle de qualité du RE (endoplasmic reticulum quality control) ERAD : ER-associated degradation ERCs : endosomal recycling compartements ERES : ER exit sites ERM : erzine-radixine-moésine ERK : extracellular signal-regulated kinase E2 : F F - : ion fluorure FSH : follicle-stimulating hormone FSH : sous-unité spécifique à la FSH Fsk : forskoline G Gi,q,s : protéine G (sous-unités i, q, s) GAP : GnRH-associated peptide GH : Growth Hormone Glc : glucose GlcNAc : N-acétylglucosamines GnRH : gonadotropin-releasing hormone GRASP : Golgi reassembly stacking protein GRP78/94 : glucose-regulated protein, 78 or 94 kda GSH : gluthation Gst : génistéine 11
14 GSU : alpha-glycoprotein hormone subunit H hcg : hormone chorionique gonadotrope HCO 3 - : ion bicarbonate Hdj-1/-2 : Human DnaJ homologue 1 & 2 HHG : hypothalamo-hypophyso-gonadique Hsc70 : protéine de choc thermique (heat shock cognate protein 70) Hsp70/90 : protéine de choc thermique (heat shock protein 70 or 90) I I : Ile (Isoleucine) I - : ion iodure IBMX : 3-isobutyl-1-methylxanthine IC : intracellulaire ICLs : boucles intracytoplasmiques (intra-cellular loops) IFN- : interféron-gamma IgA : Immunoglobuline A IP3 : inositol 1,4,5-triphosphate IP3R : IRE- : inositol- Isc : courant de court-circuit J/K/L JAM : Junctionnal Adhesion Molecule JNK : c-jun N-terminal kinase K : Lys (Lysine) K + ion potassium kb : kilobase kda : kilodalton LH : luteinizing hormone LH : sous-unité spécifique à la LH LHRH : luteinizing hormone-releasing hormone M Man : mannose MAPKs : mitogen-activated protein kinases MBL2 : Mannose-Binding Lectin 2 MDR : Multi Drug Resistance Mg 2+ : ion magnésium Mn 2+ : ion manganèse MPB : benzo[c]quinolizinium MSDs : domaines transmembranaires (membrane spanning domains) MSH : Melanocyte Stimulating Hormone MUC : mucines 12
15 MHC II : myosine II N/O Na + : ion sodium NBDs : domaines de liaison aux nucléotides (nucleotide binding domains) NDUV1 : NADH dehydrogenase [ubiquinone] flavoprotein I NEBs : corps neuro-épithéliaux (neuroepithelial bodies) NF-B : nuclear factor kappa-b NHERFs : Na+/H+ exchanger regulatory factors NKCC : cotransporteur Na+/K+/2Cl- NOS : Nitric Oxide Synthases NPPB : acide 5 -nitro- 2 -(3-phénylpropylamino) benzoïque N-ter : N-terminale N-WASP : neural Wiskott-Aldrich syndrome protein ORCC : outwardly rectifying chloride channel P P : Pro (Proline) PCL : liquide périciliaire (periciliary liquid) PCR : polymerase chain reaction PDEs : inhibiteur des phosphodiestérases (phosphodiesterase inhibitors) PDI : protéine disulfure isomérase (protein disulfide isomerase) PERK PKR : like endoplasmic reticulum kinase Pi : phosphate inorganique PIP2 : phosphatidylinositol 4,5-bisphosphate PI3K : phosphatidylinositol-3-kinase PKA/PKC : protéine kinase A / C PLA2 : phospholipase A2 PLC : phospholipase C PLC : phospholipase bêta PNECs : cellules neuroendocrines pulmonaires (pulmonary neuroendocrine cells) Po : PPQs : pyrimido-pyrrolo-quinaxolinediones PQC : peripheral quality control PP2A/2C : protéines phosphatases 2, groupe A et C PRL : Prolactine ps : picosiemens PTC : codon stop prématuré (premature termination codons) P2Y 2 : récepteurs purinergiques R R : Arg (Arginine) RCPG : récepteur couplé aux protéines G RD : domaine régulateur (regulatory domain) RE : réticulum endoplasmique (endoplasmic reticulum) R-GnRH : récepteur de la GnRH 13
16 ROMK : renal outer medullary potassium ene species) RTC ribosome translocon complex S SCN - : ion thiocyanate S : Ser (Sérine) SLC26 : solute carrier 26 SLPI : secretory leukocyte proteinase inhibitor SM : spectrométrie de masse SNAP-23 : synaptosome-associated protein, 23 kda SNARE : Soluble N-ethylmaleimide-sensitive-factor Attachement protein REceptor t/v-snare : target/vesicule associated-snare SRP : signal recognition particle SUR : récepteurs aux sulfonylurés T TCP1 : T-complex protein 1 TEER : résistance transépithéliale TGF : transforming growth factor beta TGN : réseau trans-golgien (trans-golgi network) TLR : toll like receptor TMs : transmembranaires TMEM16A : transmembrane protein 16A TNF : tumor necrosis factor alpha TRP : transient receptor potentiel TRPA : transient receptor potentiel ankyrin TRPC : transient receptor potentiel canonical TRPM : transient receptor potentiel melastatin TRPM7 : transient receptor potential melastatin 7 TRPML : transient receptor potential mucolipin TRPP : transient receptor potential polycistic TRP-PLIK : transient receptor potential-phospholipase c-interacting kinase TRPV : transient receptor potential vanilloid TSH : thyroid stimulating hormone U/V/W/X/Z UPR : unfolded protein response V : Val (Valine) VEMS : volume expiratoire maximal par seconde VEF-1 : volume expiratoire forcé Wt : wild-type X-33 : 3,7-dimethyl-1-isobutylxanthine ZO :zonula occludens 14
17 Les figures sont retrouvées aux pages mentionnées ci-dessous -bronchique () : Représentation schématique des épithéliums de surface et glandulaire recouvrant les voies aériennes () () Morphologie des cellules caliciformes en microscopie électronique à transmission. () () Cellule neuroendocrine en microscopie électronique à transmission () () Cellule de Clara en microscopie électronique à transmission () Barrière mécanique () Représentation schématique du transport des ions Na+, Cl-, et K+ retrouvé au niveau des cellules épithéliales des voies aériennes () :Clairance mucociliaire () : () : Représentation schématique des conditions de culture pour les cultures en conditions immergées ou en interface air-liquide (ALI) () Du gène à la protéine CFTR () Les transporteurs ABC () Modèle pour le pore du canal CFTR : Topologie en deux dimensions () Localisation des acides aminés arginine et lysine importants pour la perméabilité du canal aux ions Cl- (A). Représentation schématique des hélices transmembranaires (TMs) et des boucles intracellulaires (ICLs) importantes pour la fonction du canal CFTR (B) ( ) Représe«tête-bêche» (ou head-to-tail) des NBDs () Biogénèse du canal CFTR () Modèle co- et post-traductionnel du repliement de la protéine CFTR dans la membrane du réticulum endoplasmique (RE) () : Partenaires de la protéine CFTR à la membrane plasmique () () 15
18 () Carte représentant les diffé France () Modèle des systèmes de défense défaillants chez les patients CF () Classes de mutations () Physiopathologie des atteintes pulmonaires () () Thérapie de la mucoviscidose () Schéma du système hypothalamo-hypophysaire () -hypophyso-gonadique (HHG) des mammifères () Représentation schématique de la GnRH mammalienne dans la conformation dans laquelle elle se fixe sur le récepteur hypophysaire () : Représentation en deux dimensions du récepteur à la GnRH (R-GnRH) humain () Voies de signalisations activées après fixation de la GnRH sur le récepteur R-GnRH des cellules gonadotropes () Représentation des configurations du R-GnRH (R1-R4) suivant les ligands (L1- L4) () Structure de TRPM7 () TRPM7 : domaines et fonctions () :Caractéristiques physiologiques de lignées cellulaires ou de cultures primaires de cellules épithéliales des voies aériennes () :Séquence en acides aminés des isoformes I, II et III de la GnRH () : Séquence en acides aminés des agonistes et des antagonistes de la GnRH ( ) : Composés inhibiteurs de TRPM7 () :Composés activateurs de TRPM7 () 16
19 La prise en charge des atteintes respiratoires, digestives et nutritionnelles a permis la fonction pulmonaire, le potentiateur Vx-770. Il est proposé aux patients porteurs de la pourquoi la recherche de modulateurs de la protéine CFTR est un axe essentiel dans la recherche sur la mucoviscidose. Ces composés comprennent les correcteurs et les potentiateurs. Les correcteurs à la membrane plasmique via une interaction directe avec le CFTR ou du système de contrôle qualité des protéines. Les CFTR muté. Dans ce manuscrit trois travaux, qui inscrive protéine CFTR, vous seront présentés. r de nombreuses interactions protéine- de la protéine CFTR est donc un axe important dans le contexte de la mucoviscidose. Une sain ou muté F508del a été mis en évidence au laboratoire (Le Drévo et al., 2008) dans les cellules exprimant le mutant CFTR-F508delhlorure (Cl - n membranaire de la protéine ont été observées. L défauts causés par la mutation F508del. Des recherches bibliographiques ont permis AnxA5. La GnRH pourrait être une piste thérapeutique pour la mucoviscidose. La première étude de ce manuscrit a mis en évidence une augmentation significative - via le canal CFTR après une incubation des lignées cellulaires épithéliales bronchiques exprimant un CFTR sain ou muté (CFBE41o-/wt et CFBE41o-/F508del) avec la GnRH (1h, 10-9 M). Cette amélioration serait due à une Pour mettre en évidence ces résultats, western-blot du récepteur à la GnRH, de biotinylation du CFTR La deuxième étude,, a été réalisée sur des cultures primaires de cellules épithéliales nasales exprimant le CFTR-F508del. Les cultures primaires Pour ces travaux buséréline. Cet analogue est déjà prescrit en 17
20 thérapeutique et existe sous forme de spray nasal. Cette étude a nécessité la mise en place de la culture en interface air- permettant de mesurer les transports vectoriels de Cl -. Les résultats confirment sur la sortie ns Cl - traitement de la mucoviscidose. divalents tels que le magnésium (Mg 2+ ). Le principal régulateur de la concentration intracellulaire en Mg 2+ est un canal ubiquitaire appelé TRPM7. La troisième étude détaille les de fonction et de régulation du canal TRPM7 ainsi que sa, dans des cellules exprimant un CFTR normal ou muté (F508del et G551D). Pour cela nous avons utilisé des lignées cellulaires Hela transfectées stablement avec un CFTR sauvage ou muté. étudié la localisation du canal TRPM7 18
21 19

22 Zone de conduction Trachée Bronches Générations Diamètre (cm) Longueur (cm) Nombre Aire de la section (cm 2 ) Bronchioles Bronchioles terminales Zone de transition et respiratoire Bronchioles respiratoires Canaux alvéolaires. Adaptée de Sporty et al., 2008 Tapis muqueux Cils Cellules ciliées Cellules caliciformes Lame basale Vaisseau sanguin Tissu conjonctif Tissu musculaire lisse Glande sous-muqueuse. La paroi des voies aériennes est composée muqueuse (constituée épithélium reposant sur une lame basale), tissu musculaire lisse et tissu conjonctif contenant du cartilage. trachéo-bronchique humain est composé de 4 types cellulaires: les cellules ciliées, caliciformes, basales et intermédiaires. Adaptée de la Thèse de J. Roux, 2010
23 Dans la mucoviscidose, les atteinte des voies aériennes sont les plus graves et contraignantes pour les patients. Au cours du travail qui va vous être présenté nous avons utilisé des modèles cellulaires première partie. Le système respiratoire humain est composé de deux parties : les voies aériennes supérieures comprenant les cavités nasales, les sinus, le pharynx et le larynx et les voies aériennes inférieures formées par la trachée, les bronches et les bronchioles. Le rôle de ce système La partie supérieure a po le dessè. La présence du mucus et des cils permet aussi une première pour éliminer une partie des particules et des pathogènes inhalés (Hollenhorst et al., 2011). De son côté, la partie inférieure a un rôle de conduction. Elle permet le transport de chée vers les sacs alvéolaires et de dioxyde de carbone entre les poumons et le sang). Par la suite, 23 séries de divisions successives dichotomiques et asymétriques. La trachée se divise en bronches souches, puis en bronches segmentaires qui à leurs tours se divisent en petites bronches pour enfin terminer aux bronchioles. Contrairement aux bronchioles, les bronches possèdent des glandes sous-muqueuses jouant un rôle dans la clairance mucociliaire qui permet limination des particules respirées piégées dans le mucus. Les alvéoles naissent des bronchioles terminales (). La paroi des voies aériennes est composée tissu musculaire lisse conjonctif contenant du cartilage (). La totalité comprend. Il est recou constituée un épithélium de surface (ou de 20
24 - ium diffèrent selon sa localisation au sein des voies aériennes (Breeze et Wheldon, 1977). Un épithélium est composé de cellu système de jonctions cellulaires, reposant sur une lame basale qui le sépare du tissus conjonctif sous-jacent. Les cellules épithéliales présentent un pôle apical orienté vers la lumière et un pôle basa et des surfaces du corps humain ce qui en fait le premier acteur luttant contre les attaques extérieures. La peau est le meilleur exemple mais les épithéliums de la lumière intestinale et des voies respiratoires permettent aussi de protéger le milieu intérieur contre les pathogènes et les substances toxiques (Papazian et al., 2016). re la lumière et le milieu intérieur permettant de maintenir un gradient de concentration électrochimique et un transfert sélectif de molécules. Il constitue une barrière physique sécréter des Trois critères sont utilisés pour qualifier un épithélium : le nombre de couches cellulaires (simple, pseudo-stratifié ou stratifié), la forme des cellules superficielles (cubiques, pavimenteuses, cylindriques ou prismatiques) et la différenciation des structures superficielles : modification de la membrane plasmique au niveau apical (cils, stéréocils, une accumulation de substances dans les cellules superficielles (comme Dans - fonction du diamètre des voies aériennes. Ils sont composés de différents types cellulaires avec des propriétés spécifiques. -bronchique est dit pseudos le composant est en contact avec la lame basale. Cet épithélium est principalement composé lium bronchiolaire proximal est aussi pseudostratifié et comprend trois types cellulaires plus ou moins représentés : les 21
.")
25 . trachéo-bronchique de surface est un épithélium pseudostratifié, composé de quatre types cellulaires (cellules ciliées, caliciformes, intermédiaires et basales). glandulaire est constitué de trois types cellulaires (les cellules à mucus, séreuses et myoépithéliales). bordant les voies bronchiolaires humaines est de type monostratifié et composé des cellules basales, ciliées, neuroépithéliales et de Clara. alvéolaire est de type monostratifié et constitué des pneumocytes de type I (PN I) et pneumocytes de type II (PN II). la thèse de T. Joly, 2010 :. Morphologie de respiratoire au microscope électronique à transmission, C: cellule ciliée (A). Jeffery et Li, 1997
26 cellules basales qui sont rares, des cellules ciliées et des cellules sécrétrices non ciliées de tal est monostratifié et ne présente que des cellules Clara (ou clubs : les pneumocytes de type I et les pneumocytes de type II. Par conséquent au moins huit types de cellules épith respiratoire (). 1. Epithélium de surface a) Les cellules ciliées surface trachéo-bronchique. Elles représentent plus de 50% de la population cellulaire et possèdent à leur surface apicale entre 200 et 300 cils par cellules (Rhodin, 1966). Une grande quantité de battement ciliaire jouant un rôle essentiel dans la clairance mucociliaire. Ce battement ciliaire est synchronisé contribuant au déplacement du mucus. Les cellules ciliées ont une forme cylindrique et mesurent environ 20 µm de longueur, 7 µm de diamètre au niveau apical et 2 µm à la base (Breeze et Wheeldon, 1977). La longueur des cils ainsi que leur densité varient en fonction de la zone du tractus respiratoire étudiée : elle est de 5 à 7 µm dans la trachée contre 3 à 4 µm dans les bronches (Greenwhood et al.,1973) avec un diamètre moyen de 0.25 µm environ (). Les cellules ciliées sont considérées comme ayant atteintes leurs différenciations terminales et incapables de se diviser pour proliférer à nouveau (Breeze et Wheeldon, 1977). Cependant cette règle est remise en cause avec la découverte de la transdifférenciation des cellules ciliées en cellules sécrétrices (Park et al., 2006). Les cellules sécrétrices peuvent donner de nouveau des cellules ciliées étant une e surface respiratoire (Bals et Hiemstra, 2004). Les cellules ciliées Elles contribuent à la régulation du contenu en eau et en ions du mucus et du liquide périciliaire tels que le canal CFTR (Cystic Fibrosis Transmembrane conductance Regulator), ou ENaC (epithelial sodium (Na+) channel) (Barbry et Lazdunski, 1996). Des mutations au niveau du canal CFTR entraînent une pathologie appelée mucoviscidose. 22

 illustre la libération des vésicules sécrétoires dans le liquide de")
 (B) Epithélium trachéobronchique en microscopie électronique")
 sont aussi presents (A).")
27 (A) (B). Au niveau de trachéo-bronchique observé sont retrouvés les cellules ciliées (C) et les cellules caliciformes (G) (A). (B) illustre la libération des vésicules sécrétoires dans le liquide de surface (flèche). Mercer et al., 1993 (A) (B) Epithélium trachéobronchique en microscopie électronique à transmission: cellules mucosécrétantes (G), ciliées (C), et basal (B). Un vaisseau et un mastocyte (M) sont aussi presents (A). (B) met en évidence les desmosomes (d) et les filaments de kératines (f) retrouvés dans une cellule basales. (A) et (B) Jeffery et Li, 1997
28 b) Les cellules caliciformes revêtement tout au long des voies aériennes ainsi que dans 7000 cellules/mm 2 sont dénombrées dans la trachée mais leur nombre diminue dans la partie profonde des voies respiratoires (Ellefsen et Tos, 1972). Elles sont retrouvées entre les cellules ciliées et sont environ cinq fois moins nombreuses que ce type cellulaire (Wanner et al., 1996). Leur cytoplasme contient des granules sécrétoires contenant principalement des mucines, protéines principales qui composent le mucus dans lequel elles sont excrétées (Verdugo, 1990) (). Les cellules caliciformes produisent aussi des protéines à activités anti-bactériennes telles que les IgA sécrétoires (Goodman et al., 1981) et la peroxydase (Christensen et Haye, essen défensive. -renouvèlement et de remplacer les de pathologies respiratoires inflammatoires chroniques (mucoviscidose ou broncho-pneumopathie chronique obstructive, BPCO) conduisant à une hyperplasie des cellules sécrétoires maintenue par des molécules anti-apoptotiques telles que Bcl-2 (Turner et al., 2011 ; Gao et al., 2015). c) Les cellules basales Les cellules basales représentent entre 6 et 31% de la population des cellules épithéliales suivant leur localisation tout au long des voies aériennes. Ainsi, dans les conduits (supérieur à 4 mm) elles représentent 31% contre 6% dans ceux de petits diamètres (inférieur à 0.5mm) situé de surface. Ces cellules tiennent leur nom de leur proximité avec la lame basale avec laquelle relativement peu différenciées avec un rapport nucléo-cytoplasmique très important (). Les cellules basales -stratifié à la membrane basale par des hémidesmosomes et aux cellules adjacentes grâce à des desmosomes (Rock et al., 2010). Un cytosquelette abondant, les protéines de jonctions et expriment particulièrement des cytokératines (CK) telles que la CK13 (Hajj et al., 2007). 23
29 Les cellules basales sont une population de cellules souches multipotentes qui maintien es voies respiratoires peuvent proliférer et se différencier en cellules ciliées, en cellules à mucus ou en cellules Clubs -liquide (ALI) ou en culture tridimensionnel de bronchosphères ex-vivo (Smirnova, 2016). I des différences significatives de la capacité proliférative des cellules basales suivant leurs provenances (trachéales, des bronchiques ou des bronchiolaires) (Rock et al., 2010). Les cellules basales peuvent aussi contribuer à la susceptibilité et à la progression de pathologies respiratoires telles que la BPCOhronique ou la mucoviscidose (Rock et al., -renouvellement des cellules basales et la génération des cellules ciliées ou caliciformes (Rock et al., 2010). Cependant si un déséquilibre dans cette balance entre la prolifération et la différenciation des cellules basales apparait cela peut entraîner une hyperplasie des cellules basales. Dans le cas de la mucoviscidose, les changements histologiques incluent une hyperplasie des cellules dilatations des voies respiratoires ou bronchiectasies (Rock et al., 2010). d) Les cellules intermédiaires ou parabasales Les cellules intermédiaires ou parabasales forment une couche de cellules fusiformes cylindriques. En microscopie optique, les cellules intermédiaires ont été caractérisées par leur absence de caractéristiques morphologiques de cellules ciliées, neuroendocrines et sécrétrices (Boers et al., 1998). Mais en microscopie électronique à transmission elles ressemblent morphologiquement aux cellules basales sur lesquelles elles reposent, même si elles présentent un noyau supra-basal (Boers et al., établie mais elles contribueraient à la différenciation des cellules épithéliales en cellules (Donnelly et al., 1982). 2. Les glandes - 24
, les cellules séreuses (contenant des vésicules avec de nombreuses")
 contenant des vésicules denses aux")
30 Les glandes sous-muqueuses exocrines comprennent trois types cellulaires : les cellules muqueuses (responsables de la sécrétion de mucus et sont retrouvées dans la partie proximale de la glande), les cellules séreuses (contenant des vésicules avec de nombreuses macromolécules) qui sont prédominantes ainsi que des cellules myoépithéliales. Lee et Foskett, 2014 : Cellules sécrétoires non ciliées bronchiolaires (cellules de Clara) contenant des vésicules denses aux électrons entourée de cellules ciliées. Jeffery et Li 1997
31 -muqueux (Meyrick et Reid, 1970 cellulaires responsables de la sécrétion de facteurs anti-bactériens, de mucus et de fluide dans la lumière épithéliale (Jeffery, 1983 ; Basbaum et al., 1990 ; Bals et al., 1998a et b)(). Les cellules muqueuses (ou cellules à mucus), représentant 40% de la population cellulaire. Elles contiennent des vésicules remplies de mucines comme les cellules sécrétoires -bronchique. Ces cellules sont donc responsables de la sécrétion de mucus et sont retrouvées dans la partie proximale de la glande. Les cellules séreuses, quant à elles, représentent 60% de la population cellulaire et sont retrouvées dans la partie plus distale de la glande. Elles contiennent dans leurs pôles vésicules contiennent de nombreuses macromolécules telles que le lysosyme, la lactoferrine, les IgA (immunoglobuline A) sé ; Basbaum et al., 1990). Les cellules myoépithéliales lisses entourent les acini glandulaires. Ce sont des cellules le. Leur et ainsi la libération des granules sécrétoires dans la lumière (Ballard et Inglis, 2004). s. Il est constitué de cellules basales, de cellules ciliées et des cellules sécrétoires non ciliées (cellules clubs) au niveau proximal (Winkelmann et Noack, 2010) et uniquement de cellules clubs au niveau des bronchioles distales (Plopper et al., 1983). Les cellules club sont des cellules cuboïdales (). respiratoire contre des agents extérieurs (Stripp et Reynolds, 2008) en synthétisant et sécrétant la protéine CC10 (Club cells protein 10 kda) ayant un rôle anti-inflammatoire. Elles jouent des pathogènes des voies respiratoires. Dans les cellules clubs sont des cellules souches pour remplacer les cellules ciliées ou les cellules à mucus en cas de lésion (Gao et al., 2015). 25

32 Son cytoplasme contient des petites granules denses aux électrons. (L, lymphocyte) Jeffery et Li, 1997
33 Les cellules neuro-endocrines (PNEC, Pulmonary Endocrine Cells) sont retrouvées -bronch bronchiolaire appelés corps neuro-épithéliaux (NEB, Neuroepithelial Bodies) (Linnoila, 2006). Leurs cytoplasmes contiennent de nombreuses granules denses aux électrons riches en amines et neuropeptides transmetteurs ou médiateurs (Kummer et al., 2008) (). Il existe de nombreux marqueurs moléculaires des cellules neuro-endocrines. La principale neuroendocriniens tels que la calcitonine, la CGRP (Calcitonin gene-related peptide), la al., 2013). Lors de la morphogénèse des poumons, les PNECs ou les NEBs jouent un rôle dans la croissance et la différenciation pulmonaire. A la naissance, ils peuvent fonctionner comme senseur 2 -né au monde extérieur. Ensuite, les NEBs proposeraient une niche de cellules progénitrices pour permettre la régénération de al., 2000). -bronchique adulte est trois à quantité de PNECs/NEBs est augmentée dans de nombreuses pathologies pulmonaires périnatales ou post-natales (Cutz, 2015). La concentration de ces cellules est plus importante diaphragmatique ou des pneumonies, chez les enfants prématurés souffrant de dysplasie broncho-pulmonaire (Cutz et al. de la petite enfance (Young et al., 2011). Cette augmentation est aussi retrouvée par exemple dans des tumeurs endocrines telles que le cancer du poumon à petites cellules (Semenova et al., 2015). Le dysfonctionnement des PNECs/NEBs peut être impliqué dans de nombreuses n artériel (Cutz et al., 2007). recouvrant est constitué de deux types cellulaires : les pneumocytes de type I et les pneumocytes de types II. Les pneumocytes de type I sont des cellules minces et allongées recouvrant 96% de (Matthay et al., illaires 26


. sont composées de claudines, de JAM et sur lesquelles les protéines ZO-1, 2 et 3 et lient les cytosquellette aux jonctions.")
34 (A) Cellules épithéliales adjacentes Microscopie optique Microscopie électronique Jonction serrée Jonction adhérente Jonction communicante Desmosome Hémidesmosome (B) (C) Cytoplasme de la cellule 1 Jonctions communicantes Espace intercellulaire Cytoplasme de la cellule 2 2 connexons = 1 gap jonction Espace intercellulaire 6 connexines = 1 connexon Membrane cytoplasmique Cytoplasme Canal ouvert Canal fermé Complexes jonctionnels retrouvés entre deux cellules adjacentes composés par les jonctions serrées, les jonctions adhérentes et les desmosomes qui fonctionnent avec les jonctions communicantes pour permettre les fonctions physiologiques des cellules épithéliales polarisées. de microspie optique montre un marquage en rouge des tonofilaments de kératine liant les cellules adjacentes au niveau des desmosomeset marqués en vert par la desmoplakine. La microscopie électronique permet de visualiser desmosome (A). sont composées de claudines, de JAM et sur lesquelles les protéines ZO-1, 2 et 3 et lient les cytosquellette aux jonctions. Pour les, les cadhérines sont associées à caténines qui aussi au cytosquellette (B) (Papazian et al., 2016). Représentation schématique des jonctions communicantes (C) (A) Green et Gaudry, 2000, (B) Papazian et al., 2016, (C) thèse J. Roux
35 Les pneumocytes de type II sécrètent le surfactant (Castranova et al., 1988) contenant des phospholipides, des lipides neutres et des apoprotéines spécifiques (Van Golde et al., 1988 ; Jobe et al., 1987). Le surfactant permet entre autre de diminuer la tension du mucus type I (incapable de se diviser) (Weibel et al., 1971). une barrière mécanique contre les pathogènes, les allergènes et les particules inhalées, en formant une barrière physique assurée par tout un système de jonctions. Les transports ioniques au travers de lrégule le recouvrant. Le mucus, impliqué dans la clairance mucociliaire, antibactériens si nécessaire. rend-contaminants () : - Les jonctions serrées (zonula occludens ou tight junction) sont localisées au niveau apical de la membrane basolatérale des cellules épithéliales cylindriques où elles établissent la polarité de la cellule en séparant les domaines basolatéral et apical de la cellule (Gumbiner, 1993). Les jonctions serrées sont des complexes multiprotéiques formés par les protéines cytoplasmiques zonula occludens-1 (ZO-1), ZO-2 et ZO-3 associées aux protéines Junctionnal Adhesion Molecule). Elles permettent le passage sélectif de certaines molécules de petites tailles par transport paracellulaire et transcellulaire (Powell, 1981). La perméabilité des jonctions serrées peut être régulée par les cytokines (TNF-, interféron- et interleukines) ainsi que des facteurs de Les différents membres de la famille des claudines confèrent une perméabilité ionique distincte déterminée par la boucle extracellulaire de la protéine formant un pore sélectif pour les ions et la charge. Par exemple, la présence de claudine 2 dans la jonction serrée entraîne une 27
36 augmentation de la perméabilité ionique alors que la claudine 4 augmente la résistance de la barrière (pour revue : M. Loxham et al., 2014). - Les jonctions adhérentes (jonctions intermédiaires ou zonula adherens) se situent juste en dessous des jonctions serrées et assurent le contact intercellulaire. Elles sont composées de plaques intracellulaires et de protéines transmembranaires (Farquhar et Palade, 1963). Les protéines les plus importantes sont les E-cadhérines qui sont des protéines transmembranaires localisées au niveau de la membrane basolatérale des cellules épithéliales assurant le contact intercellulaire avec les cellules adjacentes par des interactions homotypiques dépendantes du calcium (Ca 2+ ) changements conformationnels des cellules (Perez-Moreno et al., aux E-cadhérines grâce à un complexe protéique composé de différentes protéines de -actinine. Les E- via les protéines adaptatrices - et -caténines (pour revue : Loxham et al., 2014). - Les desmosomes (ou macula adherens) sont retrouvés en dessous des zonula adherens de manière ponctiforme sur toutes les cellules épithéliales assurant le contact intercellulaire par chaque cellules adjacentes, composées de desmoplakines 1,2,3 et 4 (Green et Gaudry, 2000). - Les hémidesmosomes sont des petits complexes protéiques situés au niveau basal des Les jonctions communicantes,ou Gap junctions, sont constituées de connexons de 2 cellules petites molécules hydrosolubles inférieures à 1kDa par diffusion passive entre deux cellules (adénosine monophosphate cyclique), le Ca 2+ ou IP 3 (inositol 1,4,5-tris-phosphate) (Saez et al., 1986 ; Kumar et Gilula, 1996). et permet les échanges ioniques actifs entre les membranes apicale et basolatérale des 28
37 éostasie cellulaire. Ils se font majoritairement par des canaux, des échangeurs ou des co-transporteurs., adénosine triphosphate) pour son transport, ou «actifs» si le épithéliums dépend de de ces transpo s est -muqueuse respiratoire est quant à lui sécréteur (pour revue : Kunzelmann, 2001). de surface (ASL) composé du liquide périciliaire (PCL) et de mucus. Le mucus piège les particules inhalées et les évacue en dehors de voies respiratoires grâce au battement ciliaire. La composition du PCL est régulée par transports mis en jeu dans cette régulation concernent les ions sodium (Na + ), chlorure (Cl - ) et potassium (K + ) qui seront détaillés dans la suite du chapitre. + dans les cellules épithéliales suit le gradient électrochimique. Elle ENaC) retrouvé à la surface apicale. Ce canal travaille en coordination avec la pompe Na + /K + ATPase, située à la membrane basolatérale, permettant la sortie de trois ions Na + +. Ce + +, qui ion de la cellule. Le canal ENaC,, est composé de 3 sous-unités () formant le pore du canal. Il joue + dans le milieu extracellulaire par son absorption dans les cellules épithéliales (Boucher, 1994). Plusieurs types de canaux transportant le Cl - ont été mis en évidence dans les membranes apicales et basolatérales des cellules épithéliales. La sécrétion apicale de Cl- est principalement régulée par le canal CFTR et dans une moindre mesure par les canaux Cl - calcium (Ca 2+ )-dépendant (CaCC) ou les canaux Cl - à rectification sortantante ORCC (outwardly rectifying chloride channel). Cl - CaCC tels que le TMEM16A (Transmembrane member 16A) et le canal SLC26A9 a été proposée comme pouvant être des cibles thérapeutiques dans le cas de la mucoviscidose où 29
 et de mucus. Le mucus piège les particules inhalées et les évacue en dehors des voies respiratoires grâce au battement ciliaire.")
38 Na + Cl - Cl - Cl - transport des ions Na + par voie paracellulaire K + 2Cl - Na + 2K + Na + voie paracellulaire K + 3Na + : Au niveau apical, de surface est recouvert liquide de surface (ASL) compose du liquide périciliaire (PCL) et de mucus. Le mucus piège les particules inhalées et les évacue en dehors des voies respiratoires grâce au battement ciliaire. La composition du PCL est régulée par les transports ioniques principalement apicaux de Na + (absorption) et Cl - (sécrétion). L quant à elle suit le gradient osmotique passivement. Na + dans les cellules épithéliales suit le gradient électrochimique et est régulée majoritairement par le canal sodique sensible à amiloride (ENac) retrouvé à la surface apicale de ces cellules. Ce canal travaille en coordination avec la pompe Na + / K + ATPase, située sur la membrane basolatérale. de Na + est essentielle pour la sécrétion apicale de Cl - qui est principalement régulée par le canal CFTR chez et dans une moindre mesure par les canaux CaCC et ORCC. Cette sécrétion est maintenue par le co-transporteur basolatéral Na + / K + /2Cl - ). Les canaux K + identifiés dans les cellules des voies aériennes modulent le sécrétion apicale de Cl -. adapté de la revue de Edelman et Fritsch, 2005
39 la mutation du gène CFTR entraîne un défaut du transport des ions Cl - (Mall et Galietta, 2015) (). La sécrétion apicale de Cl - est maintenue par le co-transporteur basolatéral Na + / K + /2Cl - 3- /Cl - qui permettent une accumulation de Cl - au niveau basolatéral entraînant un gradient électroch - (Edelman et Fritsch, 2005). - au niveau ap K+ par des canaux identifiés dans la membrane basolatérale. Ils sont importants pour réguler le potentiel membranaire et le gradient électrochimique du Cl -. Il existe une grande variété de canaux K + dépendants du voltage, du Ca 2+ Bardou et al., 2009). En plus des canaux basolatéraux, des canaux K +, BK Ca et SK4 (contribuant à la sécrétion de Cl - Ca 2+ -dépendante), ont été retrouvés dans la membrane apicale des cellules al., 2011). + par les canaux ENaC entraînent un passage Cl - passif par la voie paracellulaire dite de «shunt» essentielle pour assurer entraînetarran et al., 2001 ; Mall et Galietta, 2015). De même, la sortie des ions Cl - par les canaux Cl - Le transport simultané des ions Na + et Cl - u suit le gradient osmotique) soit par la voie paracellulaire soit en traversant les aquaporines par la voie transcellulaire. ociliaire (Boucher, 1994 ; Tarran et al., 2001). contre les agressions extérieures. Elle est composée de deux éléments essentiels à son fonctionnement : le liquide de surface («ASL» sol» appelée liquide périciliaire (PCL) et la phase «gel» composé par le mucus (Davis et Lazarowski, 2008) (). La phase «sol» (ou le liquide périciliaire) transépithéliaux de Na + (absorption) et de Cl - 30
 épaisseur 7 µm et le battement ciliaire.")
40 La clairance mucociliaire est un système de défense innée de respiratoire contre les agressions extérieures. Elle est composée de deux éléments essentiels à son fonctionnement : le liquide de surface («ASL» : Airway Surface Liquid) épaisseur 7 µm et le battement ciliaire. Le liquide de surface empêche aussi la dessiccation de grâce à un équilibre des échanges ioniques. ASL : airway surface liquid (liquide de surface des voies aériennes), PCL : periciliary liquid (liquide périciliaire) adaptée de la revue de Frizzell et Hanrahan, 2012
41 a un rôle de lubrifiant facilitant le mouvement du mucus le long des voies aériennes (Zahm et al., 1989). Le liquide périciliaire contient aussi des protéines solubles sécrétées par les cellules séreuses et muqueuses des glandes sous-muqueuses (Kaliner, 1991) participant à la pithélium grâce à leur propriétés antibactériennes, antiprotéasiques ou antioxydantes (pour revue : Bals et Hiemstra, 2004). La phase «gel» ou le mucus i piège les particules et qui les évacue en dehors des voies respiratoires grâce au battement ciliaire. Il est produit par les cellules caliciformes et les cellules glandulaires. Les mucines (MUC) bronchiques sont des glycoprotéines de haut poids moléculaire soit sécrétées soit membranaires. Seules les mucines sécrétées constituent le réseau tridimensionnel du gel de mucus. Elles comprennent la MUC5AC, la MUC5B, la MUC2 et la MUC6 (Cone, 2009). Elles sont composées de nombreux domaines cystéine leur permettant de former des oligomères et de se polymériser pour donner la configuration tridimensionnelle al., 2000; Van Klinken et al.,1998). La MUC5AC est sécrétée par les cellules caliciformes alors que la MUC5B par les cellules glandulaires (Hovenberg et al., 1996 ; Wickström et al., 1998) et sont prédominantes dans la formation du gel de mucus (Hovenberg et al., 1996 ; Sheehan et al., 1999). Les mucines ont un rôle important dans les propriétés rhéologiques du mucus des voies respiratoires. transcellulaire, battement ciliaire jouent un rôle important dans la clairance mucociliaire. Si la clairance mucociliaire diminue cela altère ce système de défense et favorise la sécrétion de mucines est exacerbée ce qui entraîne une hyperviscosité du mucus limitant son élimination hors des voies aériennes. prenant quelques heures, les cellules épithéliales (surtout les cellules des glandes sous-muqueuses) sont capables de produire et de sécréter dans le liquide de surface de nombreuses molécules possédant des activités antimicrobiennes, antifongiques et antivirales. Ces molécules sécrétées peuvent aussi être tion et la réaction immunitaire. En sécrétant ces 31
42 lium respirat innée et adaptative pour immobiliser et tuer les micro-organismes (pour revue : Bals et Hiemstra, 2004). Parmi ces substances nous retrouvons des protéines antimicrobiennes telles que le lysozyme, la lactoferrine ou le SLPI (secretory leukocyte proteinase inhibitor). Le lysosyme est et son rôle est de lyser les parois bactériennes. La lactoferrine est une glycoprotéine avec une forte affinité pour le fer. Elle possède une activité bactériostatique et bactéricide indirecte (le fer étant nécessaire au développement bactérien) et des propriétés anti-inflammatoires. Elle est présente dans les granules de sécrétion des cellules glandulaires séreuses (Finkbeiner et al., 2010). Le SLPI est une protéine retrouvée dans antibactérienne et antivirale (Melrose et al., 1992 ; pour revue : Bals et Hiemstra, 2004). Des petits peptides antimicrobiens cationiques sécrétés jouent aussi un rôle dans la ies respiratoires. Par exemple, l -défensines sont synthétisées et sécrétées des voies aériennes et glandulaire (Singh et al., 1998) afin de lyser les bactéries Gram positives et négatives. La cathélicidine LL-37 est retrouvée dans les cellules épithéliales respiratoires (Bals et al., 1998) et cellules épithéliales des voies respiratoires peuvent sécréter la lactoperoxidase (Christensen et al., 1982) ction de radicaux libres. Elle possède des propriétés antibactériennes contre Pseudomonas aeruginosa, Burkholderia cepacia et Haemophilus influenza (Wijkstrom-Frei et al., 2003) joue aussi un rôle dans le système immunitaire en produisant de nombreuses cytokines. Les cytokines regroupent les interleukines (IL-1, IL-6, IL-8 : cytokines proinflammatoires), les interférons ou divers facteurs de croissance (TNF- TGF-vation des «Toll-like» récepteurs (TLR). Par exemple : le TLR2 reconnaît un large panel de bactéries gram+ et gram- ainsi que de champignons (pour revue viral double brin, le TLR4 les lipolysaccharides (LPS) des gram + et le TLR7 est activé par les ARN simple brin. kinases et la translocation nucléaire de NF-B sauf le TLR3 (pour revue : Loxham et al., 2014). Les cellules clubs synthétisent et sécrètent des protéines appelées CC10 (Club Cell protein 10 kda) ou CCSP (Club Cell Secretory Protein) (pour revue : Adam et al., 2014). Ces 32
43 protéines ont un rôle anti- oire alvéolaire. les cellules épithéliales respiratoires libèrent des tachykinines (la substance P et la neurokinine A). Ce sont des neuropeptide contribuant au reflex de la toux (Maggi, 1993). Les cellules épithéliales respiratoires peuvent aussi cellulaires avec des propriétés antimicrobiennes incluant les «natural killer» (NK), les cellules lymphoïdes, les neutrophiles, les cellules T et les macrophages (M2) (McLellan et al., 2015). sa fonction. processus physiologique qui Dans des conditions pathologiques telles que la BPCO, la mucoviscidose anomalies dans la structure bronchique comme par exemple des lésions, une métaplasie malpighienne ou un épaississement de la lame basale et du muscle lisse. Ces altérations perturbent la fonction épithéliale et bronchique (Jeffery, 2000 ; pour revue : Adam et al., 2014). in vitro et in vivo ont été développés afin de reproduire les conditions de l différentes étapes de ce mécanisme. Lin vitro regroupent les cultures primaires de cellules épithéliales respiratoires (trachée, bronches, bronchioles ou nasales) et les cultures de lignées cellulaires. Ces différents types de cultures ont été étudiés en deux dimensions (cultures submergées, en explant ou en air-liquide) ou en trois dimensions (sphéroïdes) (Herard et al., 1996 ; Dorscheid et al., 2001 ; Castillon et al., 2002). in vivo comprend, par exemple, les modèles animaux chez le petit mammifère le modèle chimérique de xénogreffe dans la souris nude qui la régénération espiratoire humain (Hajj et al., 2007 ; Adam et al., 2015). 33


44 Différentiation :. Les cellules entourant la lésion se différencient,, migrent puis prolifèrent en formant une métaplasie malpighienne transitoire avant de se différencier petit à petit en épithélium mature. Adam et al., 2014
45 Le mécanisme de régénération comprend quatre étapes : prolifération et la différenciation cellulaire (). Il fait intervenir de nombreux facteurs comme par exemple des composants de la matrice extracellulaire, des protéases, des facteurs de croissance et des cytokines. : ique de développer des thérapies cellulaires et régénératrices pour les atteintes pulmonaires (Navarre et al., 2015 ; Akram et al., 2016). Dans le but de comprendre polluants et des pathogènes sur le système respiratoire, des expériences in vivo (dans ex vivo (sur des tissus isolés de poumons, des biopsies de bronches ou de trachée) et des cultures in vitro ont été mis en place (Bukowy et al., 2011). peuvent être en théorie prélevé tout au long des voies aériennes. Cependant certains tissus ne sont pas faciles cornets nasaux, de la trachée, des bronches ou des bronchioles. Les biopsies bronchiques, bronchiolaires ou trachéales sont difficiles à obtenir et nécessitent obligatoirement une chirurgie. Les cellules épithéliales des polypes nasaux sont souvent utilisées, cependant elles ont des fonctionnalités différentes par rapport aux cellules des autres parties de la muqueuse nasale (Bernstein et Yankaskas, 1994). En effet, elles présentent une absorption du Na + et une perméabilité aux ions Cl - plus forte, ce qui empêche leur utilisation pour les études de pharmacologie et de métabolisme (Merkle et al., 1998). Le prélèvement des cellules nasales au niveau du cornet moyen permet de surmonter les restrictions mentionnées ci-dessus. Les cellules épithéliales nasales peuvent être prélevées post mortem chez des individus ne présentant pas de cancer et par des méthodes traumatiques ou non-traumatiques. Les méthodes traumatiques concernent les prélèvements des polypes et des cornets nasaux par chirurgie. Elles permettent de récupérer un nombre beaucoup plus important de 34
46 Modèle de culture submergée Cellules basales indifférenciées Milieu de culture Modèle de culture en interface air-liquide Cellules caliciformes différenciées Cellules ciliées de voies aériennes différenciées Membrane Milieu de culture :. Mc Lellan et al., 2015
47 cellules cependant elles nécessitent une anesthésie et cela ne peut pas être répété plusieurs fois chez un même donneur. Les méthodes non-traumatiques permettent de récolter les cellules par grattage ou brossage sont moins invasif, moins traumatisants et peuvent être répétés plusieurs fois chez un même donneur. Cependant, très peu de cellules sont collectées par rapport aux méthodes traumatiques (Bridges et al., 1991a ; Bridges et al., 1991b ; Lopez-Souza et al., 2003). Différents systèmes de cultures primaires de cellules épithéliales respiratoires humaines ont été développés A partir de biopsie, sur des supports coatés ou non pour amélio peut une forte répétabilité pour les expériences (Wiesel et al., 1983). Cette technique permet la utilisé gration ou de la prolifération cellulaire ainsi que dans les études de la réponse immunitaire aux allergènes et aux pathogènes (Ooi et al., 2007 ; Liu et al., 2007). Cette technique est moins utilisée La culture de cellules dissociées protéase (ou pronase) à basse température (Dimova et al., 2005), soit à partir de cellules collectées par brossage ou grattage (Bridges et al.,1991b ; Neugebauer et al., 2003). Les cellules dissociées peuvent être cultivées de manière adhérente avec les cultures de cellules submergées et les cultures en air-liquide, en suspension ou de manière séquentielle. La culture des cellules adhérentes submergées présente le désavantage de ne pas permettre la différenciation mucociliaire terminale contrairement à la culture en air-liquide (ALI). Pour les cultures ALI les cellules poussent sur des inserts poreux avec du milieu nutritif ( ). Ce type de culture est souvent utilisé pour les mesures de transport et de métabolisme des drogues (Bukowy et al., 2010). Dans le cas de la culture en suspension, les cellules collectées par grattages nasaux sont cultivées sous forme de sphéroïdes multicellulaires polarisés avec une faible adhérence pour leur support. Les cellules conservent les cils pendant 14 jours (Bridges et al., 1991b). La culture séquentielle comporte deux phases. Dans un premier temps, les cellules sont obtenir des cellules en suspension. Puis, pendant les semaines suivantes les cellules sont mises en rotation pour permettre la formation de sphéroïdes contenant des cellules 35
48 différenciées. Ce modèle de culture présente de nombreux avantages : la phase de prolifération permet de sélectionner les cellu cellules ciliées et sécrétrices. De plus les cellules peuvent être maintenues en culture pendant des semaines (Jorissen et Bessems, 1995) et des marqueurs de différenciation tels que les mucines peuvent être suivis (Lamblin et al., 1992). Les méthodes de cultures primaires ont été développées dans le but de reconstituer un épithélium ressemblant au tissu initial et possédant les mêmes propriétés telles que la fonction de barrière, la capacité métabolique, le transport vectoriel de molécules, la production de al., 2005). Cependant, elles présentent certains désavantages tels que le nombre limité de générations ou des différences importantes de résistance transépithéliale (TEER). Au-delà de quatre générations les cellules perdent leurs capacités à se polariser (Galietta et al., 1998). Les résistances transépithéliales montrent une variabilité interindividuelle et intra-individuelles pour un même prélèvement suivant les cultures (Papazian et al., 2016 ; Tosoni et al., 2016). Pour pallier à la difficulté de la disponibilité des échantillons de tissus des voies respiratoires, des «cultures-prêtes-à-», appelées Epiairway, ont été développées par -Unis). Ces cultures correspondent à un épithélium différencié à partir de cellules trachéo-bronchiques utilisées pour les études pharmacologiques. De plus, Lonza (Basel, Suisse) propose un panel de cultures primaires bronchiques offrant la garantie de former des cils, de produire des mucines, et avec une résistance transépithéliale élevée provenant de donneurs sains ou malades (BPCO, mucoviscidose). Pour notre étude nous avons choisi de travailler avec des cultures immergées ou en interface air-liquide (ALI) de cellules nasales obtenues par grattage. En effet, les prélèvements de ces cellules sont possibles chez des patients atteints de mucoviscidose même enfants (Mosler et al., 2008), et peuvent être utilisées pour étudier la fonction du canal CFTR (De Courcey et al., 2012). Les lignées cellulaires présentent de nombreux avantages par rapport à la culture primaire. Leur utilisation est moins coûteuse, elles sont plus homogènes et présentent moins de différences phénotypiques et les expériences sont plus reproductibles que pour les cultures 36
49 Adaptée de Bur et Lehr, 2008 et Papazian et al., 2016 RPMI 2650 Cultures primaires de cellules bronchiques 16HBE Carcinome pavimenteux de la cloison nasale Tissu sain Tissu sain immortalisé couche mince oui oui (ALI) cellules ciliées, mucosécrétantes, intermédiaires, basales cellules arrondies, cuboïdales, non-ciliées oui ~750 Forte (localisée) Transport non n.a Forte (localisée) Transport Calu-3 Adénocarcinome non-ciliées, pavimenteuses oui 1200 ~600 Forte (localisée) CFBE 41o- Beas-2B Cultures primaires de cellules alvéolaires A549 NH441 Cellules trachéales/bronchiques transformées Tissus sain bronchique immortalisé Tissu sain Adénocarcinome alvéolaire Adénocarcinome alvéolaire cellules pavimenteuses non n.a Pneumocytes de type I et II, corps lamellaires Pneumocytes de type II, inclusions liées à la membrane Pneumocytes de type II, cellules de Clara Transport, métabolisme, modèle de blessure non 1200 n.a Etude de la mucoviscidose Faibles à modérées (localisées) oui >1000 >1000 Forte (localisée) non n.a Faibles (non localisées) non 300 n.a Réparation, toxicologiques, infections Réparation, toxicologiques, infections
50 primaires. Cependant, elles sont incapables de différenciation terminale ciliée. Elles dérivent soit de carcinomes respiratoires, soit de cellules épithéliales des voies aériennes transformées. Parmi les lignées cellulaires respiratoires dérivées de carcinome nous retrouvons par exemple () : - Calu-3 : de 25 ans, exprimant de manière endogène le CFTR, des mucines (Kreft et al., 2015). Elles présentent une excellente polarisation avec un nombre important de protéines de jonctions (occludines et Ecadhérine) au niveau des jonctions serrées et une résistance transépithéliale élevée, particulièrement en culture en air-liquide (Stewart et al., 2012). La lignée cellulaire Calu-3 possède des propriétés séreuses et forment une monocouche cellulaires composée de cellules présentant un phénotype cilié et sécrétoire produisant du mucus. Elles sont capables de se différencier en interface air-liquide et de présenter des microvillosités apicales et un complexe de jonctions serrées (Papazian et al., 2016 ; Bur et Lehr, 2008). Elles sont souvent de bl- Fahlenkamp, 2016), ainsi que pour les études de transport (Mathia et al., 2002) et les processus métaboliques (Borchard et al., 2002). - RPMI 2650 : seule lignée cellulaire nasale utilisée pour les études in vitro. Elles proviennent études ont montré la présence de joncions serrées et un doublement de la résistance permettrait une utilisation plus fréquente dans les études in vitro (Bai et al., 2008 ; Werner et Kissel, 1996 ; De Fraissinette et al., 1995). - A549 : adénocarcinome pulmonaire humain la plus utilisée pour étudier les voies aériennes inférieures (alvéoles). Les cellules A549 possèdent des caractéristiques similaires aux pneumocytes de types II: des corps lamellaires, des microvillosités et expriment le CFTR, les protéines du surfactant A, B et C, Muc1 et de nombreux récepteurs. Cependant, contrairement aux pneumocytes de types II, elles ne présentent pas de résistance transépithéliale. Ces cultures sont très utilisées pour la transfection, les études pharmacologiques et pour les infections (Bhowmick et Gappa- Fahlenkamp, 2016). Elles peuvent être cultivées en culture 2-D ou 3-D. 37
51 Dans le cas des lignées épithéliales transformées il y a entre autres : - BEAS-2B : lignée cellulaire dérivant de cellules épithéliales bronchiques humaines, (Sporty et al., 2008). Cette lignée cellulaire est adaptée pour des études de réparation, -Canton et al., 2013). - 16HBE14o - : cellules épithéliales bronchiques humaines, -, transformées et immortalisées, exprimant de manière endogène le CFTR sauvage (Cozens et al., 1994). Ces cellules expriment un taux important de protéines des jonctions serrées (occludines et E-cadhérines) et une résistance épithéliale élevée (Papazian et al., 2016, Bur et Lehr, 2008), mais sont incapables de se polariser en condition air-liquide. En condition submergée les cellules 16HBE14o - forment une couche cellulaire polarisée et de nombreuses protéines de transport sont exprimées (Ehrhardt et al., 2002, Ehrhardt et al., 2003). - CFBE41o - : lignée cellulaire générée par la transformation de cellules trachéo-bronchiques cellulaire sert pour les études de transfert de gènes, des traitements alternatifs par de petites molécules et pour permettre une meilleure connaissance de la maladie au niveau cellulaire sans faire de la culture primaire (Bur et Lehr, 2008). Au laboratoire, une lignée cellulaire dérivée d'un adénocarcinome utérin humain (HeLa) a été transformée. Les cellules HeLa ont été transfectées stablement avec des plasmides contenant un ADNc codant une protéine CFTR normale (WT) ou mutée (F508del ou G551D). Ces mutations sont retrouvées chez de nombreux patients atteints de la mucoviscidose. Ces lignées ont été produites dans le but durs sur la fonction de la protéine CFTR. 38
, de 2 domaines de liaisons aux nucléotides (NBD1, NBD2) et domaine régulateur R")
52 (A) (B) CFTR Localisation du gène CFTR sur le bras long du chromosome 7 (A). Ce gène comprend 27 exons et sur environ 250 kb. Il code pour une protéine transmembranaire de 1480 acides aminés composée de 2 domaines transmembranaires (MSD1, MSD2), de 2 domaines de liaisons aux nucléotides (NBD1, NBD2) et domaine régulateur R spécifique du CFTR (B). (A) Moisan, 2015 (tel ) (B) Adaptée de Nissim-Rafinia et al., 2006
53 CFTR Le gène CFTR (Cystic Fibrosis Transmembrane conductance Regulator) a été cloné pour la première fois en 1989 par trois équipes (Kerem et al., 1989 ; Riordan et al., 1989 ; Rommens et al., le bras long du chromosome 7 au locus 7q31.2 (Zengerling et al., 1987). Il est transcrit en un ). La protéine CFTR (aussi appelée ABCC7) appartient à la famille des transporteurs adenosine-triphosphate (ATP)-binding cassette AMPc-dépendant présent dans les cellules épithéliales polarisées (Gadsby et al., 2006). 7 sous-familles (ABCA-ABCG) (Dean et al.,2001) comprenant des protéines telles que les MDR (Multi Drug Resistance), le canal CFTR et les récepteurs aux sulfonylurés (SUR1 et Nucleotide Binding Domains) ou domaines de passage actif de nombreuses molécules au travers des membranes cellulaires telles que des acides aminés, des peptides, des protéines, des ions organiques ou inorganiques, quelques toxines ou antibiotiques (Higgins, 1992). Les NBDs contiennent des motifs très conservés dont les principaux sont (ter Beek et al., 2014): - acide aminé et h est un aci co-facteur de la catalyse, le magnésium (Mg 2+ ). Ces séquences sont caractéristiques mais pas spécifiques aux transporteurs ABC. Elles constituent le noyau catalytique. - La séquence C (L-S-X-G-X-R) ou séquence signature spécifique aux transporteurs ABC est située entre les sites Walker. 39
, deux domaines de liaison aux nucléotides (NBDs). un exporteur ABC qui est schématisé ici.")

 La fixation de deux molécules au niveau des domaines RecA-like et hélice induit la dimérisation des NBDs")
54 (A) (B) (C) (A) Topologie générale transporteur ABC (vue du dessus) comprenant deux domaines transmembranaire (MSDs), deux domaines de liaison aux nucléotides (NBDs). un exporteur ABC qui est schématisé ici. En absence il est en position fermée avec les NBDs éloignés. Lorsque les molécules ATPs se fixent, il passe en position ouverte nécessaire à du substrat dans extracellulaire. (B) La fixation de deux molécules au niveau des domaines RecA-like et hélice induit la dimérisation des NBDs selon une configuration tête-bêche, laquelle changement conformationnel des TMDs. (C) Positions relatives des séquences des motifs dans les NBDs. Adaptée de ter Beek et al., 2014
55 Les transporteurs ABC comprennent aussi deux domaines transmembranaires hydrophobes (MSDs, Membrane-Spanning Domains) constitués dans la majorité des cas de 6 segments transmembranaires (). Contrairement aux NBDs, les MSDs sont plus variés et permettent de distinguer les différentes familles (Locher, 2016). Certains transporteurs possèdent des domaines «accessoires» qui peuvent être présents et influencer le transport des molécules. Par exemple, la protéine CFTR est le seul transporteur à présenter un domaine globulaire cytoplasmique appelé domaine régulateur ou domaine R. Ce domaine possède de nombreux sites de phosphorylation et est impliqué dans la régulation de la fonction du canal CFTR. fonctionner comme un canal ionique (McCarty, 2000 ; Linsdell, 2006). Elle appartient à la famille C des exporteurs ABC et de manière similaire à la majorité des transporteurs ABC elle comprend deux domaines transmembranaires (MSD1 et MSD2), deux domaines de liaisons aux nucléotides (NBD1 et NBD2) et un domaine régulateur R particulier au CFTR. 1. Les domaines transmembranaires (MSDs) La protéine CFTR comprend deux MSDs formant le pore du canal, composés de 6 segments transmembranaires chacun (TM1 à TM12) (Sheppard et Welsh, 1999). s (TM1- pore de sélectivité anionique et de faible conductance (6-9 picosiemens, ps). Les segments insertions dans la bicouche membranaire lipidique ( leurs extrémités extracellulaires par 6 courtes boucles extracellulaires (ECL1-ECL6) et à leurs extrémités cytoplasmiques par 4 longues boucles intracellulaires (ICL1-ICL4). Celles-ci sont maintenant considérées comme étant des extensions des segments transmembranaires dans le cytoplasme (pour revue : Linsdell, 2016). Les ICLs vont avoir un rôle important dans la stabilité de la protéine et dans son activité. Elles possèdent une forte interaction avec les domaines NBDs par l informations pour les changements conformationnels entre les NBDs et les MSDs permettant al., 2009 ; pour revue Hwang et Sheppard, 2009). protéine CFTR à la membrane apicale. Une mutation dans ICL2 (N287) déclenche al., 2003). Pour les boucles extracellulaires, une étude menée par Billet et collaborateurs (Billet et al., 40
 comprenant six hélices transmembranaires (TMs, en rose) connectées par de courtes boucles extracellulaires (ECLs, en bleu) et de longues boucles intracellulaires (ICLs, en")
.")
.")
56 (A) ECL 1 ECL 2 ECL 3 ECL 4 ECL 5 ECL 6 ICL 1 ICL 2 ICL 3 ICL 4 (B) NBD1 NBD1 (C) Vestibule externe Pore du canal Vestibule interne Voie cytoplasmique La protéine CFTR est composée de deux domaines transmembranaires (MSDs) comprenant six hélices transmembranaires (TMs, en rose) connectées par de courtes boucles extracellulaires (ECLs, en bleu) et de longues boucles intracellulaires (ICLs, en turquoise). Chaque MSDs est suivi par un domaine de liaison aux nucleotides (NBD1, en orange et NBD2 en vert). Les 2 ailes du CFTR MSD-NBD sont reliées par le domaine régulateur (R, en gris) (A). Modèle structural de la protéine CFTR entière proposé par homologie suite à de la structure complète du transporteur ABC bactérien Sav1866 et McjD (B). Représentation schématique du pore basé sur des analyses fonctionnelles. Le pore possède une région plus étroite comprise entre un vestibule interne et externe. Le vestibule interne est connecté au cytoplasme par une porte latérale (C). Les mêmes couleurs sont utilisées dans les schémas pour représenter les ECLs, les TMs, les ICLs et les NBDs et indiquent que les ECLs, TMs et ICLs contribuent à la formation du pore. Cependant, le domaine R pas été représenté pour les images B et C. Linsdell, 2016
57 -K1060 et S263-V1056), proche dans u couple E267-K1060 diminuent les courants Cl - dépendants du CFTR alors que celles du couple S263-V1056 les augmentent. ECL4 présente aussi deux sites de N-glycosylation sur les résidus asparagines en position 894 et 900 de la chaîne en acides aminés, déterminant le stade de maturation de la protéine CFTR (pour revue : Riordan, 1993 ; Sheppard et Welsh, 1999). La protéine CFTR est constituée de deux ailes segments TM TM et 8 (Linsdell, 2014). Le domaine de liaison aux nucléotides NBD1 et le domaine R sont retrouvés entre les deux MSDs. Les extrémités N-terminales et C-terminales des MSDs sont dans le cytosol. Le changement conformationnel de ces ailes lorsque ou non le pore aux différents ions. Le pore du canal CFTR a un diamètre de 5,3 angströms dans sa partie la plus étroite (Linsdell et al., 1997). Il a un rôle de filtre de sélectivité liant les substrats suivant leurs tailles et leurs propriétés physico-chimiques. Le pore du canal est entouré par des parties plus larges appelées vestibules intra et extracellulaire (Linsdell, 2005 homologues bactériens (Mornon et al., 2015 ; Corradi et al., 2015) et des analyses structurefonction (El Hiani et Linsdell, 2015 ; El Hiani et al., 2016) suggèrent que le vestibule intracellulaire est relié au cytoplasme par une porte latérale et non par une voie centrale. Elle serait composée par les extensions cytoplasmiques des hélices transmembranaires formant les longues boucles intracellulaires (pour revue : Linsdell, 2016) (). Certains acides aminés formant le pore ont un rôle essentiel dans la sélectivité, la sécrétion de Cl - ou les propriétés électrophysiologiques du canal. Quatre acides aminés apparaissent indispensables pour la fonction du pore niveau du vestibule externe attirerait les anions du milieu extracellulaire vers le pore (Smith et al., 2001), alors que la lysine K95 (TM1), les arginines R303 (TM5) et R352 (TM6) du vestibule interne entraîneraient les anions du milieu intracellulaire vers le pore (Aubin et Linsdell, 2006 ; pour revue : Linsdell, 2006) (). Il e dans la formation et le fonctionnement du pore (McCarty, 2000 ; Smith et al., 2001). Les segments TMs 1, 11 et 12, quant à eux, participent uniquement à la formation du pore (Ge et al., 2004 ; Fatehi et Linsdell, 2009) (). 41

 violet; R104 (TM1) orange; K190 (extension cytoplasmique de transmembranaire TM3) turquoise; R248 (extension de")
 violet; K1041(extension de transmembranaire TM10) gris; and R1048 (extension")
, 5 (rouge), 6 (violet), 11 (vert) and 12 (bleu).")

58 (A) (B). Les mêmes informations sont représentées soit sur un schéma de la topologie générale du CFTR, soit sur un modèle structural de la protéine CFTR entière proposée par homologie du transporteur ABC bactérien McjD. K95 (TM1) violet; R104 (TM1) orange; K190 (extension cytoplasmique de transmembranaire TM3) turquoise; R248 (extension de transmembranaire TM4) bleu; R303 (extension de transmembranaire TM5) rose; R334 (TM6) rouge; K335 (TM6) jaune; R352 (TM6) marron; K370 (extension de transmembranaire TM6) violet; K1041(extension de transmembranaire TM10) gris; and R1048 (extension de transmembranaire TM10) doré (A). Pour plus de clarté les NBDs ont été retirés du modèle structural. TMs 1 (orange), 5 (rouge), 6 (violet), 11 (vert) and 12 (bleu). La porte latérale est composée des extensions cytoplasmiques des hélices transmembranaire TMs 3, 4, 5, et 6 (turquoise) (B). Daprès Linsdell, 2016 I II WA et WB réprésentent les sites Walker A et B, et S.S la séquence signature C. Les sites de liaisons et de sont entourés par des pointillés : le Site I ou site«non canonique»: formé par les motifs WA et WB du domaine NBD1 et la séquence C de NBD2 et le Site II ou site«canonique»: comprend les motifs WA et WB du domaine NBD2 et la séquence C de NBD1. lathèse Billet
59 2. Les domaines de liaison aux nucléotides (NBDs) al., 2002 ; Basso et al., 2003). Tout comme les autres transporteurs ABC, les NBDs de la protéine CFTR comprennent les séquences hautement conservées : les motifs Walker A et B et le motif signature C (LSGGQ), modifié au niveau de NBD2 en LSHGH (Moran, 2014). central (60 à 70 % du (10 % du domaine) (Bianchet et al., 1997). Lewis et collaborateurs ont révélé la présence de trois régions spécifiques au NBD1 du -terminale (résidus ) appelée «insertion régulatrice» et une autre en C-terminale (résidus ) nommée «extension régulatrice». Ces régions sont très flexibles et présentent des sites de phosphorylation pour la PKA (S422, S660 et S670) (Lewis et al., 2004). NBDs. Cette dimérisation se fait de manière à ce que les NBDs se retrouvent «tête-bêche» (Lewis et al., 2004 ; pour revue : Hwang et Sheppard, 2009) (): - Site I ou site «non canonique» formé par les motifs Walker A et B du domaine NBD1 et la séquence C signature de NBD2 - Site II ou site «canonique» comprend les motifs Walker A et B du domaine NBD2 et la séquence C signature de NBD1 Les deux NBDs du canal CFTR ne présentent pas la même capacité hydrolytique. En (Aleksandrov et al., la chaîne en acides aminés, comme par exemple la p uniquement dans la partie distale du motif Walker B du site II (Tombline et al., 2004). La délétion du NBD2 de la protéine CFTR ne perturbe ni sa maturation ni son adressage à la membrane plasmique, où les protéines mutées présentent une stabilité similaire aux protéines CFTR sauvages (CFTR wt, wild type). Le domaine NBD2 influencerait al., 2007). 42
60 3. Le domaine régulateur R 195 acides aminés appelé domaine régulateur (ou domaine R) unique chez les transporteurs ABC. Le domaine R fait la liaison entre les deux parties de la protéine CFTR et contient de nombreux sites de phosphorylations. Ces sites sont les cibles des protéine fermeture du canal et le transport du Cl - (Winter et Welsh 1997 ; Liang et al., 2012 ; pour revue: Bozoky et al., 2013). Le domaine R présente de nombreuses séquences consensus phosphorylables par la PKA. Ce sont huit séquences dibasiques (R-R/K-X-S/T) en position S686, S700, S712, S737, S768, T788, S795 et S813 de la chaîne en acides aminés et quatre séquences monobasiques (R/K-X-S/T) localisées en position S690, S753, T787 et S790 (pour revue : Seibert et al., 1999). La majorité des phosphorylations des sérines du domaine R entre les S686 et 813 sont activatrices sauf les phosphorylations des Ser737 et 768 qui ont un effet inhibiteur sur la fonction du canal CFTR (Csanady et al., 2005 ; Chang et al., 1993 ; pour revue : Gadsby et Nairn, 1999). Les phosphorylations du domaine R sont importantes pour son association avec les autres domaines du canal CFTR, notamment le NBD1 (Chappe et al., 2005 ; Baker et al., 2007), mais aussi avec des protéines intervenant dans sa biogénèse et sa stabilité à la membrane plasmique telles que la protéine (Liang et al., 2012) et NHERF-1 (Alshafie et al., 2014). (Chappe et al., AMP-activated kinase) (pour revue : Bozoky et al., 2013). endoplasmique (RE). La protéine de reconnaissance du peptide signal (SRP, signal recognition particle vers le translocon Sec61. Le canal CFTR est une protéine complexe avec de nombreux domaines. Sa synthèse et son trafic intracellulaire sont hautement régulés afin que la protéine nouvellement 43
 en jaune et le domaine R en bleu.")
61 : fixation de domaine R AMPc dépendante : Le canal CFTR est une protéine complexe avec de nombreux domaines : les domaines transmembranaires (MSDs) sont représentés en rose, les domaines de liaison au nucléotides (NBDs) en jaune et le domaine R en bleu. Sa synthèse et son trafic intracellulaire sont hautement régulés afin que la protéine nouvellement synthétisée ait une bonne conformation et soit correctement insérée dans la membrane plasmique. La synthèse de la protéine CFTR commence par des mécanismes co-traductionnels de la protéine naissante dans la membrane du réticulum endoplasmique (RE) et de glycosylation. Une fois que la protéine possède une conformation correcte, elle sort du RE et rejoint de Golgi où les domaines transmembranaires glycosylés sont vérifiés avant que la protéine CFTR mature soit chargée dans des vésicules depuis trans-golgien (TGN) la membrane plasmique. La quantité de protéine CFTR à la membrane apicale dépend du nombre ARMm transcrits, de de de la fraction de protéines correctement repliées et de leurs stabilités à la membrane. La quantité de protéine CFTR à la membrane résulte balance étroite entre les protéines délivrées à la membrane (trafic antérograde), et le recyclage des protéines insérées dans la membrane. Les protéines malformées sont reconnues par le système de contrôle qualité du RE (ERQC, endoplasmic reticulum quality control) qui prévient de protéines anormales et non fonctionnelles en les retenant dans le RE avant de les rétrotransloquer et finalement de les dégrader par le système ERAD. du canal dépend de la phosphorylation du domaine R et de la fixation de deux molécules sur les NBDs. Cutting, 2015
62 synthétisée acquiert une bonne conformation et soit correctement insérée dans la membrane plasmique. Son assemblage repose sur un repliement coordonné des domaines ainsi que sur la création de contacts interdomaines pour obtenir une protéine stable et bien repliée. Cette étape est surveillée par le système de contrôle qualité du RE (ERQC) (endoplasmic reticulum quality control) (Wang et al., 2006 ; Skach, 2006) qui comprend des protéines chaperons et co- elle est dirigée dirigé vers la voie ubiquitine-protéasome, qui correspond au système de dégradation associé au RE (ERAD, endoplasmic reticulum-associated degradation) (pour revue : Cutting, 2015) (). 1. Assemblage des domaines et glycosylation du CFTR a) Synthèse et insertion des domaines transmembranaires dans la membrane du RE synthétisé puis la séquence signal de TM2 est reconnue par SRP ce qui initie la translocation du peptide néosynthétisé (Lu et al., -traductionnelle de TM1 crée la -TM4 et TM5- TM6 fonctionne aussi par paire. Ces insertions créent les boucles ECL2 (TM3-4) et ECL3 (TM5-6) (Carveth et al., 2002). Les domaines NBD1 et R sont ensuite synthétisés dans le cytosol (Kleizen et al., 2005). Une fois leurs synthèses terminées, le domaine MSD2 est synthétisé et inséré dans la gment TM8 dépend du segment TM7 via leur interaction physique par al., 2009 ; Carveth et al., séquence stop. Pour les segments TM9-10 et TM11-12, le mécanisme est similaire à celui observé pour TM3-4 et TM5-6 dans MSD1 (Carveth et al., 2002 ; Enquist et al., 2009). Le NBD2 est le dernier domaine traduit mettant fin à la traduction de la protéine par les ribosomes (Kleizen et al., 2005) (). 44
.")

63 Co-Traductionnel Post-Traductionnel : Lorsque la protéine néosythétisée émerge du ribosome, les séquences signales des hélices transmembranaires sont reconnues par la protéine de reconnaissance du signal (SRP). Les domaines de la protéine CFTR-wt sont insérés dans la membrane du RE par translocon (Sec61) puis rétrotransloqués dans le cytosol une fois la traduction achevée. Les interactions post-traductionnelles impliquant les boucles intracellulaires (ICLs) sont essentielles pour une conformation correcte de la protéine. Farinha et Canato, 2017
64 Les domaines transmembranaires sont donc insérés dans la membrane du RE alors que les ECLs se retrouvent dans la lumière du RE et les domaines R, NBDs et les ICLs dans le cytosol. b) N-glycosylation La synthèse suivre un processus de maturation débutant par sa glycosylation. La glycosylation de la protéine CFTR, co- et post-traductionnelle, joue un rôle dans (Cheng et al., 1990). Il existe 3 formes de la protéine CFTR (A, B et C) suivant son degré de glycosylation al., 1990 ; Lukacs et al., 1994). La chaîne polypeptidique nouvellement synthétisée n'est pas glycosylée et correspond à la forme A manière co- oligosaccharyltransférase retrouvée dans la membrane du RE : 2 N-acetylglucosamine (GlcNAc), 9 mannoses (Man) et 3 glucoses (Glc), additionnés aux acides aminés Asn 894 et Asn 900 retrouvés dans les séquences consensus (Asn-X-Ser/Thr) de la boucle ECL4 (Patrick et al., 2011 ; Enquist et al., 2009). La protéine CFTR présente alors un poids moléculaire Les glucosidases I sialique via des glycosyltransférases. La protéine CFTR est alors entièrement glycosylée ce 2. Contrôle qualité de la conformation des protéines CFTR (ERQC) Lors de sa biosynthèse la protéine CFTR est insérée dans la membrane du RE. Le composé de protéines co-chaperons et chaperons. Ces protéines interagissent avec le CFTR immature pour favoriser son repliement (Wang et al., 2006). 45
65 De nombreuses protéines ont été identifiées comme chaperons de la protéine CFTR dont les protéines de choc thermique tels que Hsp/c70 et Hsp90 (Hsp : Heat shock protein ; Hsc : Heat shock cognate protein) et leurs co-chaperons respectifs Hdj-1, Hdj-2 (Human DnaJ homologue 1 et 2) et Aha1 (activator of Hsp90 ATPase homologue 1) retrouvés dans le cytosol (Loo et al., 1998 ; Meacham et al., 1999). Les lectines calnexine (CNX) et calréticuline sont des chaperons décrits dans la lumière du RE (Pind et al., 1994 ; Harada et al., 2006). a) Le système de chaperons moléculaires Le peptide émerge du ribosome et interagit avec les chaperons cytosoliques de la famille des Hsp70/Hsc70 et leurs co-chaperons Hdj-1 et Hdj-2 (Farinha et al., 2002 ; Meacham et al., 1999). Les couples Hsc70/Hdj-2 et Hsp70/Hdj-1 sont impliqués dans les premières étapes de la biogénèse du CFTR en facilitant son processus de repliement et sa stabilisation (Meacham et al., 1999). Hdj-2/Hsc70 interagissent avec le NBD1 de la protéine néosynthétisée, dès sa synthèse, pour stabiliser ce domaine et son interaction avec R (Meacham et al., 1999). Hsp90 est recrutée par les protéines Hsp70/Hsc70. Le complexe Aha1/Hsp90 est essentiel pour un repliement correct du CFTR à des étapes plus tardives de la biogénèse (Loo et al., 1998, Wang et al., demandent à être précisé (Wang et al., 2006). Dans le RE les principales protéines chaperons caractérisées sont la CNX et la calréticuline. Ce sont des protéines sensibles au Ca 2+ qui jouent un rôle important dans le contrôle qualité du RE. La calréticuline est localisée dans la lumière du RE alors que la calnexine se retrouve fixée à la membrane. Ce sont des lectines qui reconnaissent et se fixent sur les chaînes glycosylées additionnées aux acides aminés Asn 894 et Asn 900. Initialement composée(s) de 14 unités (2 GlcNAc, 9 Man et 3 Glc), les glucosidases I et II clivent rapidement 2 glucoses permettant au complexe calréticuline/calnexine de reconnaître cette chaîne monoglycosylée. Ensuite, la glucosidase II retire le dernier glucose ce qui diminue repliée de quitter le RE (pour revue : Helenius et Aebi, 2004 ; Amaral, 2005). La protéine rejoint Outre son rôle de chaperon moléculaire, la calnexine liée aux chaînes oligosaccharidiques al., 2008). 46
66 b) Dégradation des précurseurs mal conformés dépendante de Les protéines chaperons jouent un rôle dans la stabilisation de la protéine CFTR lors de son repliement mais aussi dans la reconnaissance des protéines mal conformées. Le système de dégradation associé au RE (ERAD) consiste en un marquage des protéines mal repliées par plusieurs résidus ubiquitines (ubiquitylation). Ce processus consiste à attacher de manière covalente des ubiquitines en les polymérisant sur le substrat au niveau de résidus sérines (S660, S686, S700, S712, S737, S795, S1444 ou S1456) (McClure et al., 2012 ; pour revue : Pranke et Sermet- ubiquitylation de la protéine peut se faire de manière co- ou post-traductionnelle tout au long de la biosynthèse ou lors de son intégration cytosolique de la membrane du RE (Xiong et al., 1999 ; pour revue : Pranke et Sermet- Gaudelus, 2014). Les protéines marquées sont ensuite dirigées vers le protéasome. Le complexe du protéasome 26S forme un tonneau et contient deux sous-unités : la sous-unité catalytique 20S dans la partie centrale et la sous-unité régulatrice 19S qui comprend de nombreuses protéines ATPases et des composés fixant les substrats ayant subi al., 1996). Le polypeptide marqué pénètre dans le cylindre où il est coupé en peptides puis dégradé par des enzymes cytosoliques. Trois enzymes sont indispensables pour cette modification : l'enzyme d'activation de l'ubiquitine (E1), l'enzyme de conjugaison de l'ubiquitine (E2) et l'ubiquitine-ligase (E3) (Hershko et al., 2000). Les protéines chaperons Hsp70/Hsc70 déclenchent le système ERAD suite à leur -ligase CHIP (C-terminus of Hsc70-interacting protein). Cette ubiquitine- UbcH5 (Meacham et al., 2001 ; Younger et al., 2004). calnexine/calréticuline empêchant les protéines néo- Cependant, si les protéines restent mal conformées, la chaîne oligosaccharidique est réduite à 8 mannoses pour être reconnue par la lectine EDEM (ER-degradation enhancing 1,2- mannosidase-like protein) qui va les diriger vers les voies de dégradation ERAD (pour revue : Farinha et Amaral, 2005 ; Farinha et Canato, 2017) 47
67 Le système ERAD possède un rôle central dans la dégradation des protéines - agrégats de protéines vers la voie de dégradation dans des vacuoles lysosomales. Cette voie comprend quatre étapes autophagosomes, le ciblage et la fusion des autophagosomes avec les lysosomes puis : Pranke et Sermet-Gaudelus, 2014). Les protéines CFTR sont regroupées au niveau des sites de sortie du RE appelés ERES (ER exit sites) COPII (coat protein complex II). Ces vésicules bourgeonnent à partir de la membrane du RE et sont recouvertes des complexes hétérodimériques Sec23/Sec24 et Sec 13/Sec31 (Yoo et al., 2002). Le transport de ces protéines cargos implique des protéines de transport telles que la GTPase Sar1 (Yoo et al., coat protein complex I), depuis le compartiment cis-golgien puis médian et enfin trans-golgien (Yu et al., 2007). La voie de transport dite conventionnelle est dépendante des protéines adaptatrices telles que la GTPase et al., 2002). Une étude en microscopie électronique montre un faible taux de protéines CFTR dans ERES (Bannykh et al., conventionnel de transport permet le trafic de la protéine CFTR entre le RE et la membrane Golgi, engagerait la protéine t-snare (target membrane associated SNARE), syntaxine 13, al., 2002). Golgi, GRASPs (Golgi reassembly stacking protein) dans le transport non conventionnel de la protéine CFTR. Ce chemin amenant la protéine partiellement glycosylée à la membrane plasmique (Gee et al., 2011) requiert la monomérisation, la phosphorylation et la relocalisation al., 2016). La phosphorylation de GRASPs est induite par le déclencheme Unfolded protein 48

68 : Guggino et Stanton, 2006
69 response assure sa fonction de canal ionique (Gee et al., 2011). La stabilité du CFTR à la membrane dépend de son interaction avec de nombreuses protéine-protéine de acides aminés cytosquelette (Cushing et al., 2008 ; Ranganathan et Ross, 1997). Pour la protéine CFTR -terminale qui permet cette interaction. Les partenaires du CFTR à domaines PDZ sont par exemple NHERF-1 (Na+/H+ exchanger regulatory factor isoform-1), NHERF-2, NHERF-3 (ou CFTR-associated protein 70 KDa, CAP70), NHERF-4 et CAL (CFTR-associated ligand) (Guggino et Stanton, 2006) (). Le domaine ERM (ezrine/radixine/moésine) de NHERF- liaison du complexe NHERF--1 fixe le CFTR via son domaine PDZ (Cantiello, 1996 ; Naren et al., 2003 ; Li et al., 2004). Le rôle de NHERF- 1 dans la stabilisation de la protéine CFTR à la membrane implique son interaction avec des GTPases de la famille des protéines Rho (Farinha et al., 2013). Les protéines PDZ régulent aussi le destin de la protéine CFTR : NHERF-1 et NHERF- à la membrane apicale, la surexpression de CAL réduit la demi-vie de la protéine CFTR mature en promouvant son endocytose et sa dégradation lysosomale (Cheng et al., 2004). Le taux de canaux CFTR dans la membrane plasmique dépend de 3 processus : le des protéines du manteau et adaptatrices avec des partenaires du réseau intracellulaire incluant les protéines du cytosquelette, des protéines motrices et des protéines kinases (Kravtsov et Ameen, 2013). Lukacs, 2007) Clathrin Coated Vesicles 49
70 CFTR (au niveau de la queue C- -2 (Assembly Polypeptide-2) (Madden et Swiatecka-Urban, 2012). -Urban et al., 2004). La protéine N-WASP (neural Wiskott-Aldrich syndrome protein) est aussi très importante dans le pr lorsque la protéine N-WASP est inhibée (Ganeshan et al., 2007). u CFTR sont également régulées par des GTPases de la et al., 2004). AP-2 et les clathrines se dissocient et sont recyclées vers la membrane plasmique (Swiatecka- Urban et al., 2004). Les canaux CFTR internalisés sont orientés vers la voie de dégradation lysosomale ou recyclés à la membrane plasmique via des endosomes de recylage (Guggino et Stanton, 2006). Le recyclage des protéines CFTR internalisées est considéré comme étant le mécanisme principal pour maintenir une quantité de protéines CFTR fonctionnelles à la es CFTR à la (actine-f) (Haggie et al., 2006). Plusieurs protéines sont engagées dans le recyclage du CFTR, notamment Rme-1. Cette protéine régule la sortie des canaux CFTR du compartiment de recyclage (ERC, endosomal recycling compartements) (Okiyoneda et Lukacs, 2007). La GTPase TC10 participe aussi au recyclage du CFTR. Elle inhibe la protéine CAL qui favorise la dégradation lysosomale de la protéine CFTR de Golgi et celle insérée dans la membrane plasmique (Cheng et al., 2004, Cheng et al., 2005). domaine régulateur par des protéines kinases. Deux sites de phosphorylation possibles ont aussi été mis en évidence dans le domaine NBD1 et correspondent aux sérines 660 et 670. al., 1991 al., 2004), la CK2 (caseine kinase 2) (Cesaro et al., 50

71 :. du CFTR nécessite la phosphorylation de nombreux sites du domaine régulateur (R) par les protéines kinase A et C (PKA, PKC) et la fixation sur les domaines NBDs. Meng et al., 2017 Ouvert :. La fixation des molécules induit la dimérisation des domaines NBDs entraînant un changement conformationnel au niveau des MSDs permettant du canal CFTR. de par enzymatique des NBDs, termine le cycle relâchant. Ce cycle pas réversible, ne peut pas redonner de et libérée par de pas utilisée pour le transport de Cl -. Moran, 2017
72 ents conformationnels déclenchés suite à la phosphorylation par la PKC (Chappe et al., 2003 ; Seavilleklein et al., 2008). phosphorylation du CFTR par la PKA. Dans les épithéliums in vivo par les agonistes -a (Choi et al., 2007 ; Tarran et al., 2006). al., 2005) déclenchant leurs dimérisations responsables de changements conformationnels ). Le mécanisme moléculaire de dimérisation des protéines ABC procaryotes montrent clairement une interaction entre les NBDs et les boucles intracellulaires (ICL) (Mornon et al., 2008 ; Mornon et al., 2015 ; Serohijos et al., 2008 ; Belmonte et Moran, 2015). Des mutations au niveau de la boucle ICL2 et ICL4 entraînent une tivation du CFTR. Deux acides aminés retrouvés dans ICL2 semblent être importants: la S263 qui augmente le courant dépendant du CFTR et le E267 qui le diminue (Billet et al., 2013). Le mutant ICL4 quant à lui affecte la synthèse de la protéine CFTR (Cotten et al., 1996). Ces deux boucles intracellulaires joueraient un rôle dans la transduction du modifications conformationnelles majeures des domaines transmembranaires influencent transmembranaires pour ouvrir le canal ionique (Mornon et al., 2015). et al., 2005 ; Csanády et al., 2010 ; pour revue : Moran, 2017) (). - (Zeltwanger et al., 1999 ; Csanády et al., 2010). Ceci est illustré par le fait que le canal CFTR peut être ouvert par des -hydrolysable c -PNP (Aleksandrov et al., 2001 ; Weinreich et al., 1999). 51
73 (PP2A et PP2C) qui déphosphorylent le domaine R de la protéine CFTR (Becq et al., 1994). La protéine CFTR est principalement un canal anionique formant un pore qui permet le passage rapide au travers de la membrane plasmique des ions Cl -, bicarbonates (HCO 3- ) et nions. La perméabilité relative aux différents ions du canal CFTR est la suivante : SCN - - > NO 3 > Br - > Cl - > I - > formate > F - (Hwang et Kirk, 2013). Outre son rôle de canal anionique, le canal CFTR est un régulateur essentiel de nombreux canaux ioniques tels que les canaux Cl - (ORCC et CaCC), le canal potassique ROMK, le canal sodique ENaC ou - /HCO3 -. Ces deux points seront développés dans ce chapitre. Mais, la protéine CFTR participe aussi à de nombreux processus biologiques. En effet, elle possède une activité anti-inflammatoire (Bodas et Vij, 2010) et participe à la réponse immunitaire (Schroeder et al., 2002). Enfin, le canal CFTR serait retrouvé à la surface des lysosomes et des endosomes es (Swanson, 2006). Les propriétés fonctionnelles du canal CFTR ont été mises en évidence par des techniques biophysiques. Le canal CFTR possède une faible conductance comprise entre 6 et 9 ps et fonctionne comme un canal unique avec une faible sélectivité pour les anions (Linsdell et Hanrahan, 1998 ; McCarty et Zhang, 2001 ; Hwang et Kirk, 2013). Le rôle physiologique du canal CFTR est de réguler la conductance anionique au niveau de la membrane apicale de nombreuses cellules épithéliales (Frizzell et Hanrahan, 2012). En plus des ions Cl - polyatomiques tels que les ions HCO 3-. Ces ions sont importants dans la régulation du ph des solutions en les alcalinisant. Un ph alcalin est nécessaire pour le bon fonctionnement des al., 1968 ; Kopelman et al., al., 1999 ; Coakley et al., 2003). Le CFTR est aussi perméable aux ions Thiocyanates (SCN - ). Ces ions participent à la réponse immunitaire. Ils possèdent un effet antimicrobien, en générant Staphyloccocus aureus (S. 52
74 aureus) et Pseudomonas aeruginosa (P. aeruginosa) (Moskwa et al., 2007), et antiinflammatoire (Chandler et al., 2015). Le canal CFTR est aussi perméable au gluthation (GSH). Ce tripeptide possède un fort pouvoir antioxydant. Son rôle est de protéger les cellules du stress oxydatif causé par les Reactive Oxygene Species) lors de la réponse inflammatoire (pour revue : Hudson, 2001). 1. Le canal CFTR et les canaux Cl - Les canaux rectifiant sortant ou ORCC sont présents dans les membranes apicale et - et sont régulés par et al., 1993, Jovov et al., milieu extracellulaire par le CFTR (Schwiebert et al., 1995, Hwang et al., 1996). Des études utilisant des protéines tronquées ont permis de montrer la nécessité des domaines NBD1 et R du canal CFTR dans la régulation des canaux ORCC (Schwiebert et al., 1998). Les canaux CaCC sont des canaux Cl - activés par le Ca 2+. Ils sont présents à la membrane apicale des cellules épithéliales où ils participent, avec le canal CFTR, au transport al., 2002). Ces canaux semblent aussi avoir une relation fonctionnelle avec le CFTR. La signalisation calcique, activant les CaCCs, -1 a été montrée (pour revue : Kunzelmann et al., 2012). 2. Le canal CFTR et le canal potassique ROMK Les canaux ROMK (renal outer medullary potassium) sont situés principalement à la membrane apicale des cellules rénales (Ho et al., 1993 ; Lee et Hebert, 1995) où ils participent s la membrane (Mennitt et al., 1997). Leur expression et leur activité sont modulées par de nombreux facteurs tels que le ph sulfonylurées comme le glibenclamide (inhibiteur du CFTR) (pour revue : Hebert et al., 2005). Une relation fonctionnelle entre les canaux ROMK et les canaux CFTR se fait au niveau apical 53
75 NHERF2. Ce complexe interagit avec les motifs PDZ des deux canaux (Yoo et al., 2004). 3. Le canal CFTR et le canal ENaC Le canal ENaC est un canal Na + transmembranaire composée de 3 sous-unités (, et ) qui forment un pore. Dans + par ENaC crée une différence de potentielle - par la voie paracellulaire dite de shunt (Boucher, 1994 ; pour revue canaux ENaCs est augmentée ce qui entraîne une hyperabsorption de Na+ contribuant à la déshydratation de liquide de surface recouvrant les voies aériennes (Stutts et al., 1995 ; Donaldson et Boucher, 2007). Le canal ENaC + et dans la clairance mucociliaire (Stutts et al., 1995 ; Donaldson et Boucher, 2007). Des études montrent que le canal CFTR inhibe le canal ENaC (Stutts et al., 1995 ; Mall et al., 1996 ; Hopf et al., 1999). Cependant, les mécanismes moléculaires mis en jeu ne sont pas encore totalemen ENaC e aux récepteurs purinergiques P2Y 2 (pour revue : Schwiebert et al., ENaC semble être régulée par la concentration en Cl - intracellulaire (König et al., 2001). Une interaction physique ENaC nécessaire à son activation (Gentzsch et al., 2010) /HCO est un ion majeur sécrété dans les liquides de surface recouvrant la majorité des voies respiratoire, digestive et reproductrice où il joue un rôle de tampon. Cette sécrétion est majoritairement due au canal CFTR et aux échangeurs Cl - - /HCO 3 tels que les transporteurs anioniques de la famille SLC26 (solute carrier 26) (Ishiguro et al., 2007, pour revue : Lee et al., 2001). Certains échangeurs de la famille SCL26 tels que SLC26A3 (DRA) et SLC26A6 (PAT1) (Singh et al., canal CFTR est augmentée en présence de SLC26A3 (Ko et al., 2004). Une interaction physique entre le 54
76 ions Cl - et HCO 3 - a été mis en évidence lors de la capacitation des spermatozoïdes (Rode et al., 2012). 5. Le canal CFTR et les aquaporines se (AQPs). smique (pour revue : initialement été décrite dans les cellules respiratoires humaines et dans les oocytes de al., 1999 ; Schreiber et al., 2000, Pietrement et al., al., 2014a) et les cellules de Sertoli (Jesus et al., ession du canal CFTR dans les oocytes de Xénope augmente la perméabilité de ces protéines ce qui suggère un rôle du canal CFTR dans la régulation des AQs (Schreiber et al., 1999 ; Schreiber et al., al., 2008). De nombreux modulateurs du CFTR ont été mis en évidence, par des similarités de ge à haut débit (pour revue : Zegarra- Moran et Galietta, 2017). Seuls quelques exemples de modulateurs de CFTR utilisés en recherche seront décrits dans le chapitre suivant. 1. Activateurs/potentiateurs du CFTR La modulation de la fonction du canal CFTR par des activateurs ou des potentiateurs soit par son action sur des éléments régulateurs de la protéine CFTR. 55
77 a) Liaison directe sur le canal CFTR Il existe de nombreuses molécules activatrices du CFTR telles que : - Les xanthines -isobutyl-1-methylxanthine) (Drumm et al., 1991), lecpx (8- cyclopentyl-1,3-diphenylxanthine) (Arispe et al., 1998), X-33 (3,7-dimethyl-1-isobutylxanthine) (Bulteau et al., 2000) ou la théophyline (Chappe et al., 1998). Ces molécules inhibent des phosphodiestérases et agiraient directement sur le domaine NBD1 du CFTR. - Les flavonoïdes : flavonnes (apigénine), isoflavonnes (génistéine), des dérivés de flavonnes concentration (Wang et al., 1998). Le site de fixation de la molécule se trouve au niveau de la glycine G1349 (Melin et al., 2005) dans le site I non catalytique de NBD2 ce qui favorise la - Les benzimidazolones tels que NS004 et NS1619 agissent uniquement sur les protéines phosphorylées (Al-Nakkash et al., 2001). La génistéine et les dérivés be phloxine B et les benzo(c)quinolizinium (MPB) ont été identifiés comme potentiateurs. MPB- 07 ne modifie ni la con suggère un effet potentiateur direct de cette molécule sur la protéine CFTR (Cai et Sheppard, 2002 ; Becq et al., 1999). b) Modulation du système de régulation de la protéine CFTR Pour notre étude nous avons utilisé la forskoline (Fsk). Cette molécule est un activateur Coleus forskohlii. -wt à des AMPc active les PKA qui vont alors phosphoryler le canal CFTR. La phosphorylation du domaine R activation de la conductance de CFTR. -unité phosphodiestérases (PDE) le cpt-amp. 56
78 Les PDE empêche la phosphorylation dépendante des PKA en dégradant les permet tion du canal CFTR en empêchant la déphosphorylation du domaine R (Cobb et al., 2003). Enfin, inhiber les phosphatases permet de maintenir une activation du canal CFTR (Becq et al., qui inhibe la PP de type 2A (Luo et al., 1998). Les activateurs et potentiateurs du CFTR sont très nombreux et appartiennent à CFTR il est fort possible que de nouveaux modulateurs soient découverts et la découverte de nouveaux composés pharmacologiques pourrait faire progresser la recherche fondamentale sur le canal CFTR. 2. Inhibiteurs du CFTR Les inhibiteurs du canal CFTR peuvent interagir directement avec la protéine en bloquant le canal en position ouverte tels que le DPC (Acide diphenylamine-2-carboxylique), NPPB (acide 5 -nitro- 2 -(3-phénylpropylamino) benzoïque) glibenclamide. sur le blocage direct du pore du côté intracellulaire (Scott-Ward et al., 2004 ; Sheppard et Robinson, 1997 ; Walsh et al., 1999 ; Zhang et al., 2000 ; Zhou et al., 2002). La glycine hydrazyde ou GlyH-101 est aussi une molécule inhibitrice bloquant le canal entre depuis la partie extracellulaire du pore pour bl et actif à faible concentration (Muanprasat et al., 2004). le cas par exemple du thiazolidinone ou CFTR inh-172 qui affecte les mécanismes de «gating» du CFTR. Son effet passe par une stabilisation de son état fermé (Caci et al., 2008). Cet inhibiteur est souvent utilisé en recherche pour étudier la fonction du canal CFTR (Ma et al., 2002 ; Yang et al., 2003). Le CFTR inh-172 a une solubilité maximale de 20 µm dans une solution saline. Pour les études sur certains tissus ou modèles animaux, la solubilité est un facteur limitant dans certaines conditions expérimentales. Par conséquent, des dérivés avec une solubilité plus importante et une efficacité satisfaisante ont été développés : tetrazolo
79 et oxo-172 (Sonawane et Verkman, 2008). Ils sont utilisés pour les études où le CFTR inh-172 semble avoir une efficacité plus faible. Les PPQs (pyrimido-pyrrolo-quinaxolinedi composé le plus intéressant dans cette famille est une petite molécule non chargée qui inhibe complètement le CFTR appelée, PPQ-102 (Tradtrantip et al., une modification du «gating» de la protéine CFTR. 58
 présentent deux sytèmes de défense contre les pathogènes non")

80 : Audrezet et al., 2015 Les voies aériennes des patients atteints de mucoviscidose (CF) présentent deux sytèmes de défense contre les pathogènes non fonctionnels. La partie gauche du schéma montre que la perte des canaux CFTR cause une diminution du ph inhibant antimicrobiennes des molécules sécrétées par respiratoire. A droite du schéma est présenté la perte du canal CFTR dans les glandes sous-muqueuses. Le mucus déshydraté stagne et devient un site préférentiel pour le développement bactérien. Adaptée de Stoltz et al., 2015
81 La mucoviscidose ou fibrose kystique du pancréas est la maladie héréditaire autosomique récessive la plus fréquente dans la population caucasienne. Elle affecte approximativement personnes dans le monde (Farell, 2008) avec une distribution très hétérogène suivant les origines ethniques (O'Sullivan et Freedman, 2009). En effet, elle est très fréquente dans les populations de type caucasiennes alors que les populations asiatiques et africaines sont les moins touchées. La France compte environ 6500 patients et al., 2015) mais la densité de population atteinte est différente selon les régions (). La Bretagne est une des régions les plus concernées avec une incidence de 1 naissance sur 3669 (Audrezet et al., 2015). Cette pathologie est due à des mutations du gène CFTR codant la protéine CFTR. La protéine CFTR est un canal Cl - AMPc-dépendant. La mutation du gène CFTR entraîne un dysfonctionnement du canal CFTR causant un défaut de sécrétion des ions Cl -. Cette - déclenche une hyperabsortion des ions Na +, par les canaux ENaC, par voie paracellulaire conduisant à SL. La diminution - du ph des ions HCO 3 par le CFTR, activité des molécules antimicrobiennes sécrétée et al., 2012 ; Stoltz et al., 2015). Le mucus déshydraté devient un site préférentiel pour le développement bactérien (Pezzulo et al., 2012 ; Stoltz et al., 2015). Les défauts du système immunitaire et la stagnation du mucus déclenchent une comprenant ies aériennes conduisant à la dégradation des tissus (Mall et Boucher, 2014 ; Stoltz et al., 2015) (). n diagnostic établit très tôt, ont té de vie des patients. et al., de la maladie avec une atteinte pulmonaire très importante. 59

82 Adaptée de Edmondson et Davies, 2016 Protéine CFTR mature et fonctionnelle Absence de protéine CFTR fonctionnelle Absence de protéine CFTR fonctionnelle Perturbation de la régulation du canal Altération de la sélectivité et de la conductance du canal Peu de protéines fonctionnelles Instabilité à la membrane Protéine CFTR néosynthétisée Absence de synthèse de protéine Dégradation des protéines mal repliées Protéine CFTR néosynthétisée Protéine CFTR néosynthétisée Peu de protéines néosynthétisées Protéine CFTR néosynthétisée Réticulum endoplasmique Réticulum endoplasmique Réticulum endoplasmique Réticulum endoplasmique Réticulum endoplasmique Réticulum endoplasmique Réticulum endoplasmique ARNm complet ARNm Instable et tronqué ARNm complet ARNm complet ARNm complet ARNm correct ARNm incorrect ARNm complet Noyau Gène CFTR Noyau Gène CFTR Noyau Gène CFTR Noyau Gène CFTR Noyau Gène CFTR Noyau Gène CFTR Noyau Gène CFTR CFTR fonctionnelle Défaut de maturation de la protéine CFTR Perturbation de la régulation du canal Défaut de conductance et de sélectivité Altération de la stabilité de Défaut de la stabilité de la protéine mature Mutations non-sens ou aux sites Insertions/délétions Mutations faux-sens Délétions aminés Mutations faux-sens Délétions aminés Mutations faux-sens Délétions aminés Mutations faux-sens Mutations faux-sens Délétions aminés Gly542X Trp1282X Arg553X 621+1G T Asn1303Lys Ile507del Arg560Thr Gly178Arg Gly551Ser Ser549Asn Arg117His Arg347Pro Arg117Cys Arg334Trp kb C T G A G A 5T 4326delTC Gln1412X 4279insA 1
83 2009 mutations touchant le gène CFTR ont été décrites et classées suivant leurs effets sur la synthèse, la maturation et le fonctionnement de la protéine ( Il existe six classes (Welsh et Smith, 1993 ; Zielenski et Tsui, 1995 ; pour revue : Fajac et De Boeck, 2016 ; Castellani et Assael, 2017) (): -Classe I : mutations altérant. Ce type de mutation entraîne une absence totale ou partielle de la protéine. Cette classe comprend les mutations non-sens, des insertions/délétions et des mutations au niveau conduisent à un codon stop prématuré (PTC, Premature Termination Codons). Les ARNm et/ou les protéines produites sont tronqués et instables donc rapidement dégradés. Elles sont retrouvées chez à peu près 10% des patients. -Classe II : mutations perturbant de la protéine CFTR provoquant sa dégradation prématurée. Les protéines produites présentent un défaut de repliement et un défaut fonctionnel. Elles sont retenues dans le RE par le système ERQC av commune, F508del, appartient à cette classe et elle est présente sur au moins un des allèles de 90% des patients. -Classe III : mutations perturbant du canal CFTR. Le canal CFTR est exprimé à la membrane mais il est très peu fonctionnel. Elles correspondent le plus souvent à des mutations faux- est présente chez 4 à 5 % des patients dans le monde et la plus fréquente correspond à la mutation G551D observée chez à peu près 3% des patients. -Classe IV : mutations altérant du canal CFTR. Ce sont le plus souvent des mutations faux-sens retrouvées dans les domaines transmembranaires sélectivité (par exemple : R117H). -Classe V : mutations altérant du CFTR. Ces mutations produisent une protéine CFTR normale mais impactent le taux de et/ou la stabilité des transcrits. Des 60
84 mutations au niveau du promoteur, des mutations faux-sens diminuant le trafic intracellulaire synthèse protéique et donc la quantité de protéines CFTR fonctionnelles à la membrane. -Classe VI : mutations altérant -over de la protéine CFTR à la surface apicale La mutation F508del est la mutation la plus fréquente chez les patients atteints de mucoviscidose (Thibodeau et al., 2010). Elle est localisée dans le domaine NBD1 de la protéine CFTR et raîne un défaut de repliement de la protéine, ce qui empêche sa maturation (Riordan, 2005). Les protéines malformées sont reconnues par le système ERQC qui aide à la conformation et à ion des protéines anormales et non fonctionnelles en les séquestrant dans le RE et les dirige rapidement vers les voies de dégradation ubiquitine-protéasome (Vashist et Ng, 2004). 1. Défaut de repliement La mutation F508del entraîne un défaut de conformation de la protéine CFTR (Riordan et al., 1989). Des études cristallographiques du domaine NBD1 de Lewis et structure de la protéine CFTR à part au niveau du site de mutation où la topographie de surface de la protéine est modifiée (Lewis et al., 2004 ; Lewis et al., 2005). Comme la mutation F508del est présente dans une région du domaine NBD1 interagissant avec MSD1, fie des contacts interdomaines plus que le repliement du NBD1 lui-même (Lewis et al., 2004 ; Lewis et al., 2005). Pissara et son équipe en 2008 ont montré que les modifications des acides aminés (F494N/Q637R ou F429S/F494N/Q637R) dans le domaine NBD1 permettent de rétablir partiellement le trafic et la fonction du CFTR-F508del suggérant un défaut de repliement du NBD1 (Pissara et al., 2008). de mutations de second site I539T et de 4 prolines (S422P, S434P, S492P, A534P) dans le domaine NBD1 restaure le transport, la 61
85 stabilité membranaire et la fonction du CFTR (Aleksandrov et al., 2012) suggérant que la correction de structure du NBD1 est suffisante pour supprimer les défauts de conformation et de fonction du CFTR-F508del. (Mendoza et al., 2012 ; Rabeh et al., 2012) démontrent que de second sites (I539T G550E, R553M ou R555K) dans le domaine NBD1-F508del ne suffit pas pour corriger le défaut de repliement de NBD1. La mutation F508del compromet la stabilité thermodynamique du NBD1 et des interactions interdomaines (Thibodeau et al., 2010 ; Mendoza et al., 2012 ; Rabeh et al., 2012 ; Cui et al., 2007 ; Du et al., 2005). La mutation F508del déstabilise un contact hydrophobe entre NBD1 et ICL4 essentie Mornon et al., 2008 ; Serohijos et al., 2008). Les mutations de second site doivent être associées à des pour supprimer les défauts conformationnels et fonctionnels causés par la mutation F508del (Mendoza et al., 2012 ; Rabeh et al., 2012). Par exemple, cette interaction est rétablie la mutation R1070W qui favorise une amélioration de la maturation de la protéine CFTR- F508del (Thibodeau et al., 2010). La chaîne latérale du tryptophane restaure NBD1 et ICL4 (Thibodeau et al., 2010). Les défauts de repliement de la protéine CFTR-F508del sont donc attribués à une instabilité du NBD1 et à entre NBD1 et MSD2. Ces défauts doivent être corrigés pour que la biosynthèse, la conformation et la fonction de la protéine mutée se rapprochent de celles observées pour la protéine sauvage (Mendoza et al., 2012 ; Rabeh et al., 2012). Une étude plus récente de He et collaborateurs en 2015 indique que le rétablissement de la stabilité thermique du domaine NBD1 de la protéine mutée F508del est suffisant pour rétablir la maturation de la protéine CFTR (He et al., 2015). 2. Modification des interactions avec les chaperons moléculaires La mutation F508del modifie les interactions de la protéine CFTR avec certaines protéines chaperons du système de contrôle qualité du RE (ERQC) (Pind et al., 1994). Les protéines chaperons reconnaissent les défauts de repliement de la protéine mutée et la retiennent dans le RE (Vashist and Ng, 2004). Meacham et collaborateurs, en 1999 (Meacham et al., 1999), décrivent une interaction plus forte de la protéine Hsc70 avec la protéine CFTR-F508del, par rapport à la protéine CFTR-wt, pouvant expliquer la rétention de la protéine mutée dans le RE. Scott- Ward et Amaral, en 2009, ont confirmé cette observation par la technique de résonnance 62
86 plasmonique de surface (Scott-Ward et Amaral, 2009). La rétention prolongée de la protéine permettant la dégradation des protéines malformées (Meacham et al., 1999 ; Scott-Ward et Amaral, 2009). Les co-chaperons aidant au repliement de la protéine sont remplacées par les co-chaperons nécessaires pour la dégradation. Ces dernières du CFTR- F508del et ciblent les protéines malformées en déclenchant leur ubiquitylation pour les diriger vers le protéasome où elles seront dégradées (Meacham et al., 2001 ; Younger et al., 2004 ; Younger et al, 2006) UbcH5 (Meacham et al, 2001 ; Younger et al., 2004), le complexe RMA1-RNF5, la derline-1 p97 pour délivrer la protéine ubiquitinilée au protéasome (Younger et al, 2006). Le couple Hsp90/Aha-1 possède aussi un rôle crucial dans la séquestration de la protéine mutée dans le RE. En effet, tout comme la protéine Hsc70, le co-chaperon Aha-1 possède une affinité plus importante pour la protéine mutée CFTR- Aha-1 favorise la biosynthèse du canal CFTR muté alors que sa surexpression séquestre la protéine CFTR-F508del déclen entraînant sa dégradation (Wang et al., 2006 ; Koulov et al., 2010 ; Hutt et al., 2012). La calnexine (CNX) lie le CFTR sauvage et le CFTR muté F508del mais possède -F508del contribuant à une interaction prolongée entre ces deux protéines (Pind et al, 1994 ; Okiyoneda et al., exprimée, la biosynthèse de la protéine sauvage est diminuée alors que ni le transport ni la fonction du canal CFTR-F508del ne sont impactés. Cette protéine chaperon seule ne peut donc pas expliquer la rétention et la dégradation de la protéine mutée dans le RE (Farinha et Amaral, 2005 ; Okiyoneda et al., 2008). La majorité des protéines F508del sont dirigées vers la voie de dégradation dépendante d-protéasome par la machinerie Hsp/Hsc70 se trouvant en amont de la CNX (Farinha et Amaral, 2005). -F508del dans le RE déclenche un stress du RE qui active une réponse appelée UPR (Unfolded Protein Response) (Kerbiriou et al., 2007 ; Bartoszewski et al., transmembranaires du RE : IRE- (inositol- ), PERK (PKR like endoplasmic reticulum kinase) et ATF6 (activating transcription factor 6). Ces voies de signalisation sont mises en place pour diminuer la traduction des protéines malformées et éviter leur accumulation dans le RE, faciliter la conformation des protéines en synthétisant des protéines chaperons ou encore augmenter la 63
87 dégradation des protéines mal con revue : Farinha et Canato, 2016). Kerbiriou et collaborateurs, en 2007, ont observé une ion de la protéine chaperons Bi -F508del (Kerbiriou et al., 2007). Des travaux de Gomes-Alves, en 2010, ont confirmé cette observation et ont GRP94 dans les cellules exprimant le CFTR-F508del (Gomes-Alves et al., 2010). 3. La protéine CFTR-F508del à la membrane a) Défaut fonctionnel Certaines protéines malformées CFTR-F508del échappent au système de contrôle du RE et parviennent à la membrane plasmique où elles vont avoir une fonction altérée et une stabilité réduite (Dalemans et al., 1991 ; Wang et al., 2011). -F508del est thermosensible et décline rapidement à 37 C (Wang et al., 2011 ; Liu et al., 2012 le temps de fermeture du canal est augmenté (Dalemans et al., 1991 ; Haws et al., 1996). La mutation F508del réduit le taux de phosphorylation de NBD1 sur la S660 par la PKA contribuant au défaut de transport des ions Cl - et à une activation plus faible des canaux dépendant de la phosphorylation (Drumm et al., 1991 ; Hwang et al., 1997 ; Wang et al., 2000). repliement intrinsèque de NBD1 affectent le transport, la stabilité et la fonction du canal CFTR (Aleksandrov et al., 2012 ; Mendoza et al., 2012 ; Rabeh et al., 2012). b) Système périphérique de contrôle qualité (PQC, peripherical quality control) Les protéines mutées ayant atteint la membrane plasmique sont rapidement endocytées par le système périphérique de contrôle qui comprend des co-chaperons tels que Bag-1, Hsp40, HOP et Aha-1 (pour revue : Chin et al., 2017). Les protéines internalisées sont dirigées vers la dégradation lysosomale suite à leur interaction avec le système être due à son instabilité à 37 C ou aux interactions diminuées avec le complexe NHERF-1-64
88 ezrine-actine (Okiyoneda et al., 2010). Ce complexe est important pour la localisation, la Favia et al., 2010 ; Monterisi et al., 2012). -F508del en agissant sur les protéines à domaines PDZ. Ces - protéines cibles au cytosquelette (Cushing et al., 2008 ; Ranganathan et Ross, 1997). la GTPase, Rac1, déclenche la phosphorylation de -F et NHERF-1 augmentant la stabilité de la protéine CFTR-F508del à la membrane plasmique (Moniz et al., 2013). Cette -F dépendante de PIP 2 qui permet la liaison de la protéine CFTR-F508del exprimée à la surface membranaire au protéine CFTR-wt, NHERF-1 est exprimée sous la membrane plasmique alors que dans les cellules exprimant une protéine CFTR-F508del, elle est retrouvée dans le cytoplasme. NHERF-1 est donc un facteur clé pour stabiliser la protéine CFTR-F508del à la membrane (Guerra et al., 2005). c) Thérapies envisageables pour la mutation F508del De nombreuses études in vitro ont montré que la protéine CFTR mutée peut échapper au contrôle qualité du RE et continuer sa maturation, en diminuant la température (Denning et al., 1992), en créant un second site de mutation (Teem et al., 1993) ou en utilisant du glycérol (Chin et al., 1996). La restauration partielle de la fonction du canal CFTR par ces manipulations suggère des stratégies thérapeutiques envisageables pour la e la protéine CFTR. protéine CFTR-F508del. Une diminution de la température réduit les interactions de la a dégradation lysosomale (Pankow et al., indiquer de nouvelles cibles thérapeutiques. En plus de la recherche de cibles potentielles pouvant améliorer la synthèse de la protéine CFTR-F508del, de nombreux travaux -F508del. Pour cela des correcteurs et des potentiateurs ont été développés (pour revue : Farinha et Matos, 2016). 65
89 La relation entre la mutation du gène CFTR est différente suivant les organes touchés. Cette corrélation est bien établie pour les atteintes pancréatiques (Kristidis et al., 1992 ; Castellani et al., 2008). Les classes de mutations I, II et III sont associées à une insuffisance pancréatique et une maladie plus sévère. Les patients sont homozygotes ou hétérozygotes pour deux mutations dites «sévères». Les classes IV, V et VI sont associées à une fonction du pancréas normale et un phénotype moins sévère. mutation dite «modérées» (Kristidis et al., 1992). Dans le cas des atteintes respiratoires, le lien entre le génotype et le phénotype est plus difficile à mettre en évidence. Un même génotype peut aboutir à une atteinte de la fonction pulmonaire très variable selon les patients, suggérant la participation de facteurs supplémentaires (Dork et al., 1991 ; Clain et al., 2001 ; Collaco et al., 2010 ; Weiler et Drumm, 2013). Des études familiales de jumeaux (monozygotes ou dizygotes) et de fratrie ont permis de mettre en évidence un rôle des facteurs environnementaux sur les atteintes pulmonaires. En effet, les jumeaux monozygotes présentent des fonctions pulmonaires proches contrairement aux jumeaux dizygotes et aux fratries (Mekus et al., 2000 ; Vanscoy et al., 2007). La différence des atteintes respiratoires observée lorsque les jumeaux monozygotes ne vivent plus ensemble détermine la part des effets environnementaux (état nutritionnel, niveau social, infections) (Collaco et al., 2010). pulmonaire retrouvé chez les patients (Collaco et al., 2010). Dans la mucoviscidose, la perte de la fonction du canal CFTR dans les épithéliums respiratoires déclenche une réaction inflammatoire et une réaction immunitaire exacerbées causant des lésions des tissus et la fibrose pulmonaire. De nombreux gènes participant à ces processus ont été identifiés pour expliquer la sévérité des atteintes pulmonaires (Weiler et Drumm, 2013). Les gènes MBL2 (Mannose-Binding Lectin 2) et NOS (Nitric Oxide Synthases) impliqués dans la défense antimicrobienne (Buranawuti et al., 2007) ou les gènes codants pour le TNF- et IL-8 dans la réponse inflammatoire (Drumm et al., 2005 ; Hillian et al., 2008). Une part de la variabilité phénotypique peut aussi être expliquée par la présence me site de mutation ou une modification 66
90 Mutation du gène CFTR Protéine CFTR anormale : transport ionique altéré Thérapies oligonucléotidiques Modulateurs du CFTR Déshydratation du liquide de surface Baisse de la sécrétion des ions HCO 3 - Altération de la clairance mucociliaire Protection antibactérienne défectueuse Contournement du CFTR Stagnation du mucus Inflammation Infection Bronchiectasies Traitement symptomatique Antibiotiques Transplantation pulmonaire Insuffisance respiratoire :. Principales cibles des traitements symptomatiques et des thérapies émergentes. Fajac et De Boeck, 2016
91 de la séquence en cis sur un allèle du gène CFTR. Par exemple, un patient homozygote F508del portant une mutation supplémentaire R553Q sur un de ces allèles CFTR, présente un phénotype moins sévère (Dork et al., 1991 modérées sur un même allèle est associée à un phénotype sévère (Clain et al., 2001). La protéine CFTR est retrouvée à la membrane apicale des cellules ciliées des épithéliums sécréteurs des appareils respiratoire, digestifs et reproducteurs. Elle joue un rôle (Gadsby et al., 2006). Les anomalies moléculaires du gène CFTR entraînent des atteintes importantes notamment au niveau des voies respiratoires, du tube digestif et de ces annexes, du tractus génital et des glandes sudoripares. Ces manifestations cliniques seront détaillées dans la suite du chapitre. 1. Atteintes respiratoires CFTR chez les patients CF (Cystic Fibrosis) déclenche une hyper-absorption de Na +, densité du mucus. Cette hyper-absorption du Na + dépendants des canaux ENaCs par le CFTR. diminue le battement ciliaire et la clairance mucociliaire (Tang et al., 2014). Le mucus stagne et devient un terrain propice au développement bactérien (Pezzulo et al., 2012 ; Stoltz et al., 2015) (). Les infections b premières années, le mucus est surtout colonisé par Staphylococcus aureus (S. aureus) et Haemophilus influenza (H. influenza) qui sont majoritairement remplacées par Pseudomonas aeruginosa (P. aeruginosa) antibiotiques peut conduire à des infections chroniques causant des réactions inflammatoires thélium bronchique et la dégradation progressive de la fonction respiratoire et des tissus pulmonaires (Mall et Boucher, 2014 ; Stoltz et al., 2015). Les bactéries ne sont pas les uniques agents microbiologiques présents lors du développement de la pathologie. Les infections fongiques en particulier par Aspergillus fumigatus peuvent aussi être observées (Marguet et al., 2016). La perte totale ou partielle de la fonction du canal CFTR chez les patients CF induit une diminution des défenses antibactériennes conduisant au recrutement des neutrophiles (Cantin et al., 2015). Les lipopolysaccharides (LPS) bactériens et les protéines 67
92 pro-inflammatoires libérées par les neutrophiles déclenchent le TLR4 conduisant à une activation de la voie NF--8 (Cantin et al., 2015). Les neutrophiles libèrent des élastases qui dégradent petit à petit les tissus bronchiques (Cantin et al., 2015) et produisent aussi des ROS créant un stress oxydatif. Lorsque le stress oxydatif devient trop important le facteur de transcription Nrf2 (Nuclear factor erythroidderived 2-like) inhibe la voie NF-B. Chez les patients CF ce facteur est défectueux, la réponse inflammatoire est donc incontrôlée (Cantin et al., 2015). Les défauts du système immunitaire et la stagnation du mucus déclenchent une qui conduit à la dégradation des tissus et une fibrose pulmonaire (Mall et Boucher, 2014 ; Stoltz et al., 2015). Les patients peuvent aussi présenter des sinusites, des polypes nasaux et des mucocèles nécessitant parfois des interventions chirurgicales au niveau du nez ou des sinus. La mesure du volume expiratoire maximal par seconde (VEMS) chez les malades 2. Atteintes digestives Le canal CFTR est exprimé tout au long du tractus gastro-intestinal et du pancréas. Le dysfonctionnement de cette protéine entraîne la déshydratation de la bile, des sucs pancréatiques et des sécrétions intestinales. Le développement de la pathologie digestive dû à la perte de fonction de la protéine CFTR (De Lisle et Borowitz, 2013). o Avant la naissance, un iléus méconial (intestin hyperéchogène) est détecté par échographie et affecte 15% des patients nouveau-nés (Scotet et al., 2012). Il est souvent associé aux mutations les plus sévères. Les patients plus âgés peuvent présenter des constipations ou une paresse intestinale et développer une occlusion partielle ou totale de de été détectés par biopsies intestinales (Raia et al., 2000) tout comme un microbiome altéré (Manor et al., 2016). o Atteinte du pancréas e chez près de 90% des patients à la naissance (Davies et al., des sucs pancréatiques empêchent l'écoulement des enzymes pancréatiques dans le tube 68
93 digestif et provoque la dégradation progressive des tissus amenant à la fibrose du pancréas (Wilschanski et Novak, 2013 ; Gibson-Corley et al., 2016). Ces enzymes ne peuvent plus digérer les nutriments ingérés. Les protéines, les graisses, les vitamines et les minéraux sont mal absorbés et éliminés sous la forme de diarrhées graisseuse ou stéatorrhées (Gelfond et Borowitz, 2013). Cette malabsorp dénutrition (Gelfond et Borowitz, 2013) pancréas endocrine, les patients pourront déclarer un diabète insulino-dépendant appelé CFRD (Cystic Fibrosis-Related Diabetes) (Gibson-Corley et al., 2016). Le diabète rencontré pendant les exacerbations pulmonaires. 40 à 50% des patients CF présentent cette pathologie. Le diabète est associé à une atteinte respiratoire plus grave. Un bon contrôle de mortalité (pour revue : Castellani et Assael, 2017). o Les atteintes hépatobiliaires : La bile épaissie provoque une obstruction des canaux biliaires empêchant au foie de jouer son rôle dans la digestion et le métabolisme. Cette maladie obstructive débute par une cirrhose biliaire focale progressant vers une cirrhose biliaire multilobaires et une hypertension portale. Ces complications ne sont retrouvées que chez une minorité de patients mais ne sont pas rares. La fréquence actuelle des manifestations cliniques au niveau du foie est difficile à déterminer. La prévalence des atteintes sévères est élevée à n (Flass et Narkewicz, 2013). 3. Autres atteintes o Atteinte du tractus génital Le système reproducteur peut lui aussi être perturbé avec une absence bilatérale congénitale des canaux déférents détectée chez plus de 90% des patients (Yu J et al., 2012). Cette atteinte cause déférents sont obstrués par une accumulation de mucus visqueux et épais. Chez la femme une diminution de la fertilité peut être observée. La glaire cervicale épaissie peut gêner le passage des spermatozoïdes (Kopito et al., 1973). 69
94 o Atteinte osseuse Les patients montrent une densité minérale osseuse faible et présentent un risque -Gaudeluset al., 2011). Le risque de maladies osseuses est lié à la malnutrition, au faible indice de masse corporelle, à s facteurs comme par exemple. La protéine CFTR est exprimée dans les cellules osseuses et son rôle direct sur le métabolisme osseux ne peut pas être exclu (Aris et al., 2002 ; Shead et al., 2007). La forme classique de la mucoviscidose peut être distinguée des pathologies associées au dysfonctionnement de la protéine CFTR ou CFTR-RD (CFTR-related disease). Dans le cas des formes dites typiques de mucoviscidose, le test de la sueur est supérieur à 60 mmol/l, les patients présentent au moins un de signes cliniques et sont t al., 2014). Le diagnostic est posé après une démarche clinique qui débute par un test de la sueur suivi par une analyse moléculaire du gène pour établir le génotype. Les 30 mutations les plus fréquentes sont testées. Si la ou les mutations ne sont pas id (délétion et duplication) qui sont retrouvés dans plus de 2% des anomalies du gène CFTR (Audrezet et al., 2004 ; Niel et al., 2004). Si un doute persiste pour le diagnostic avec un test de la sueur intermédiaire (entre 30 et 60 mmol/l) et sans identification de la mutation du gène CFTR, un test fonctionnel peut-être effectué. Ce test consiste à mesurer la différence de potentiel (ddp) transépithélial au niveau de la muqueuse nasale en analysant le transport ionique médié par le canal CFTR. Chez un individu sain la ddp est en moyenne de -20 mv contre - atypiques de la mucoviscidose (Sermet-Gaudelus et al., 2010). La mucoviscidose dite «atypique» ou CFTR-RD correspond à une forme moins sévère de la maladie et est souvent monosymptomatique. Elle est diagnostiquée chez les patients présentant un test de la sueur normal (inférieur à 30 mmol/l) ou intermédiaire (30 à 60 mmol/l), au moins une caractéristique phénotypique de la mucoviscidose, une mutation 70
95 du gène CFTR CFTR mise en évidence par une ddp nasale élevée (Fanen et al., 2014). Elle plus modérée (Ferrari et Cremonesi, 1996). Les pathologies associées au CFTR-RD sont (CBAVD), certaines formes de pancréatites chroniques et des bronchiectasies (Bombieri et al., 2011). La majorité des traitements disponibles sont symptomatiques. Ils sont mis en place pour maintenir une bonne nutrition, une amélioration de la clairance mucociliaire et traiter les infections et les inflammations (pour revue : Fajac et De Boeck, 2016). Les traitements classiques incluent le remplacement des enzymes pancréatiques, des physiothérapies respiratoires, des trai : Castellani et Assael, 2017). Dans le cas des atteintes pulmonaires, les sécrétions épaisses sont éliminées par traitement mucolytique, antibiotiques) qui diminuent la viscosité du mucus ou permettent son élimination. La prévention et le traitement des infections respiratoires représentent le challenge le plus important dans la prise en charge de la mucoviscidose (Conway et al., 2014). Les traitements par antibiotiques nébulisés sont très utilisés pour éradiquer et contrôler les infections chroniques par Pseudomonas aeruginosa. s sont utilisés : des antiinflammatoires stéroïdiens ou non (corticostéroïdes ou ibuprofène), des antibiotiques (Azythromycine), des inhibiteurs des neutrophiles (Ibuprofène) ou de la production neutrophilique (Dornase ) (Cantin et al., 2015). antibiotiques (Lands et al., 2007 ; Spagnolo et al., 2013 ; Southern et al., 2012). Afin de diminuer la viscosité du mucus due libéré par les neutrophiles dégradés, les patients sont traités par Pulmozyme (ou Dornase Pour hydrater le mucus, des solutions hypertoniques (NaCl) permettent une sortie (Tildy et Rogers 2015). Le pas encore totalement élucidé. 71
 Composés de translecture des codons stop prématurés Probabilié de canaux CFTR internalisés de canaux CFTR à la membrane de canaux CFTR dans la cellule Activité totale du canal CFTR : (A) Les")
96 Classe VI (CFTR sauvé) Classe III Classe II, IV, V,VI CFTR totalement actif Classe I Classe II, V, VI (B) Correcteurs (C) «Stabilisateurs» (D) Potentiateurs Potentiateurs? (A) Composés de translecture des codons stop prématurés Probabilié de canaux CFTR internalisés de canaux CFTR à la membrane de canaux CFTR dans la cellule Activité totale du canal CFTR : (A) Les composés de translecture des codons prématurés dont les antibiotiques aminoglycosides (gentamicine, tobramycine) ou PTC-124 permettent à la transcription du CFTR de se poursuivre ce qui augmente la quantité de protéine produite. (B) Les correcteurs (Vx-809, Vx- 661) induisent un repliement correct des mutants du CFTR qui ne sont plus retenus dans le RE et dirigés vers la dégradation. Le nombre de canaux CFTR présent à la membrane plasmique est donc plus important. (C) Les composés stabilisateurs améliorent ou la rétention du CFTR à la surface cellulaire augmentant le nombre de canaux présent à la membrane. (D) Les potentiateurs tels que Vx-770 activent le CFTR. Pour cela ils augmentent la probabilité (Po) du canal en régulant son ou en jouant sur la conductance. Adaptée de Bell et al., 2015
97 sont généralement traitées avec tritionnel des patients (Corey et al., 1988). Depuis le clonage du gène CFTR en 1989 et les connaissances grandissantes sur la biosynthèse, la structure, et la fonction du canal CFTR, la recherche thérapeutique (pharmacothérapi défauts liés à cette pathologie. Le principal but étant de prévenir les atteintes pulmonaires. 1. La thérapie génique La thérapie génique consiste à délivrer un ADNc (ADN complémentaire) de la protéine CFTR sauvage, dans une cellule, fonctionnelle. Du fait de sa grande taille, l dans des vecteurs pour entrer dans la cellule. Ces véhicules de transfert de gènes sont soit plus étudiés sont des molécules non virales cationiques qui forment un complexe avec la (Lindberg et al., 2012). Le traitement est bien toléré par les patients mais de nombreux défis ont été mis en évidence dus au par les vecteurs, à la demi-vie courte de une faible efficacité) et au une réponse inflammatoire (Lee et Southern, 2013 ; Griesenbach et al., 2015 une administration du traitement par nébulisation du complexe gène/liposome a montré une augmentation significative statistiquement de 3,7 % du volume expiratoire forcé (VEF-1) mais avec une signification clinique modeste (Alton et al., 2015). 2. La thérapie protéique ermettant de corriger la protéine anormale suivant la classe de mutation (). 72
98 a) Suppresseur de codon stop Les mutations non-sens de classes I entraînent un arrêt prématuré de la traduction. Elles sont présentes chez environ 10 % des patients dans ou PTC-124 a des effets similaires aux antibiotiques aminoglycosides à forte concentration et induit la une protéine fonctionnelle (Welch et al., 2007). Il est déjà utilisé pour le traitement de la dystrophie musculaire de Duchenne. Des essais de phase II avec une administration orale de ce composé ont montré une amélioration de la fonction du canal dans des cellules épithéliales nasales (Kerem et al., 2008 ; Sermet-Gaudelus et al., 2010). Un essai clinique de phase III chez les patients CF a montré une efficacité du traitement seulement chez les patients non traités par nébulisation à la tobramycine (antibiotique souvent utilisé pour son activité anti-bactérienne contre P.aeruginosa) (Kerem et al., 2008 ; Kerem et al., 2014). b) canal CFTR Ces modulateurs incluent les correcteurs, les potentiateurs et les stabilisateurs du CFTR. Un correcteur est une molécule pharmacologique qui augmente le nombre de protéines CFTR matures et fonctionnelles à la membrane apicale des cellules épithéliales (Van Goor et al., 2011) Un potentiateur membrane apicale mais qui a une activité réduite (Ramsey et al.,2011) Un stabilisateur est une molécule qui stabiliserait la protéine CFTR à la membrane plasmique (Zaman et al., 2016) Les modulateurs sont mutations spécifiques. Ils ont un effet uniquement sur certaines mutations produisant des protéines défectueuses. L a développé le premier potentiateur présentant un effet chez les chez les patients porteurs de la mutation de classe III G551D. Le Vx-770 appelé Ivacaftor et commercialisé sous le nom Kalydeco accroit la fonction respiratoire et réduit les exacerbations pulmonaires. En effet, Ivacaftor améliore le transport de Cl - dépendant du canal CFTR conduisant à une hydratation plus élevée du liquide de surface des voies aériennes et à une augmentation de la fréquence de battement ciliaire permettant (Van goor et al., 2009). Pour cela, le composé se fixerait directement 73
.")
99 Thérapie génique Complexe ADN-Liposome Réparation ARN Agents PTC Correcteurs Potentiateurs : Développement clinique ou pré-clinique de modulateurs du CFTR (de: evalué en Juillet 2016). Fajac et De Boeck, 2016
100 le principal défaut des mutations de classe III (Eckford et al., 2012). tels que QBW251 (Novartis), GLPG1837 (Galapagos), ou CTP-656 (Concert Pharmaceutical) (). La mutation F508d plus fréquente chez les patients atteints de mucoviscidose. Par conséquent, de nombreuses recherches se concentrent pour trouver des molécules pharmacologiques permettant de corriger les défauts apportés par cette mutation. La majorité des protéines CFTR-F508del sont dégradées par le système de contrôle qualité du RE. Cependant, une petite quantité de protéines mal conformées atteint la surface membranaire avec un temps de demi-vie court. De nombreux composés appelés correcteurs améliorent in vitro le transport intracellulaire de la protéine CFTR-F508del ce qui permet à une plus grandes quantité de plasmique. pharmaceutique Vertex a développé le Vx-809 (Lumacaftor), un correcteur proposé dans le cas de la mutation F508del. Lumacaftor supprime le défaut de conformation en améliorant le contact entre NBD1, MSD1 et MSD2 (Ren et al., 2013). Les études in vitro montrent que Lumacaftor améliore la biosynthèse de la protéine et les secrétions de Cl - dans les cellules épithéliales bronchiques homozygotes pour la mutation F508del (Van Goor et al., 2011). Cependant, potentiateur est nécessaire pour corriger le défaut de régulation (ouverture/fermeture, «gating») de la protéine. milliers de patients, révèlent une amélioration significative de la fonction respiratoire et une baisse des exacerbations pulmonaires, chez les patients homozygotes pour la mutation F508del, lorsque Lumacaftor est combiné avec Ivacaftor (Wainwright et al., 2015). Lde la fonction respiratoire avec ce traitement est moins importante que celle observée chez les patients porteurs de la mutation G551D vacaftor seul. Le traitement Vx-809 combiné à Vx-770 est prescrit en Europe aux patients homozygotes pour la mutation Orkambi. De nombreux composés correcteurs sont en développement comme par exemple Tezacaftor (VX-661 de Vertex) ou Riociguat (Bayer) (). peuvent être combinés avec les correcteurs et les potentiateurs comme par exemple le «stabilisateur» Cavosonstat (N91115, Nivalis). Ce composé participe à la stabilisation de la protéine CFTR à la surface membranaire en 74
101 inhibant la S-nitrosoglutathione reductase qui est augmentée dans les cellules épithéliales bronchiques humaines CF. la dé membrane plasmique (Zaman et al., 2016 ; Donaldson et al., 2017). 3. Approches alternatives a) Activation des canaux Cl - De nouvelles cibles thérapeutiques telles que les canaux Ca 2+ -dépendant (TMEM16A ou ANO1) présents à la membrane apicales des cellules caliciformes sont envisagées (Mall et Galietta, 2015). Pour cela, une augmentation du Ca 2+ intracellulaire (ex : Moli-1901) ou une activation des récepteurs purinergiques P2Y 2 par des anal : Denufusol) sont testés (Tildy et Rogers, 2015). - non Ca 2+ -dépendant comme le SLC26A9 pourraient être aussi de nouvelles cibles thérapeutiques en modulant leur expression ou leur activité à la membrane à la membrane plasmique (Mall et Galietta, 2015). b) La cystéamine Une voie alternative pour permettre au CFTR-. Le système lules CF (Luciani et al., 2012) à cause de la (De Stefano et al., 2014). appelé cystéamine, déjà utilisé dans le traitement des cystinoses, avec les flavonoïdes du thé vert (Gallates d'épigallocatéchine ou EGCG). La combinaison des deux molécules diminue la concentration en sel de la sueur, améliore Cl - par les cellules nasales, re cytokines inflammatoires dans les expectorations des patients et améliore le volume expiratoire forcé de 4%. Ces résultats ont été observés chez des patients homozygotes pour la mutation F508del (De Stefano et al., 2014 ; Tosco et al., 2016). 75

102 Cellules mélanotropes Cellules corticotropes gonadotropes Somatotropes lactotropes thyréotrope est une glande située dans la selle turcique à la base du cerveau et directement reliée à pour former le système hypothalamo-hypophysaire. Elle est composée de deux lobes : le lobe antérieur glandulaire et le lobe postérieur nerveux. postérieure contient des terminaisons nerveuses à ocytocine et vasopressine. antérieure contient six lignées cellulaires endocrines : les cellules corticotropes, thyréotropes, somatotropes, lactotropes et mélanotropes libérant respectivement (Adenocorticotrophic Hormone), la TSH (Thyroid Stimulating Hormone), la GH (Growth Hormone), la PRL (Prolactine) et la MSH (Melanocyte Stimulating Hormone) et enfin les cellules gonadotropes produisant la LH (Luteinizing Hormone) et la FSH (Follicle-Stimulating Hormone). la thèse de A.L Shang, 2011
103 Gonadotrophin-Releasing- Hormon -hypophysaire. Elle est composée de deux lobes : le lobe antérieur glandulaire et le lobe postérieur nerveux. les pituicytes et des terminaisons nerveuses sensibles aux hormones vasopressine et es : les cellules corticotropes, thyréotropes, somatotropes, lactotropes et mélanotropes libérant Hormone), la GH (Growth Hormone), la PRL (Prolactine) et la MSH (Melanocyte Stimulating Hormone) et enfin les cellules gonadotropes produisant la LH (Luteinizing Hormone) et la FSH (Follicle-Stimulating Hormone). Des neurosécrétions hypothalamiques pulsatiles de GnRH en passant par le système porte hypothalamohypophysaire (). Le récepteur de la GnRH (R-GnRH) est exprimé à la surface des cellules gonadotropes. La GnRH se fixe sur son récepteur et relaie les informations du système nerveux central induisant la synthèse et la sécrétion des gonadotropines LH et FSH, importantes dans le contrôle de la reproduction humaine. Ces hormones sont libérées dans la circulation sanguine et stimulent la production des stéroïdes sexuels et des gamètes. Chez la femme, la LH entraîproduisant la progestérone La FSH, de son côté, favorise la croissance ui va se développer pour et testostérone. La GnRH, les gonadotropines (LH et FSH) et les gonades forment lhypophyso-gonadique (HHG) (). Chez les mammifères, la GnRH est un décapeptide sécrété de manière pulsatile dont la sécrétion est régulée par de nombreux facteurs dont des neurotransmetteurs et des neuropeptides. Son récepteur appartient à la superfamille des récepteurs couplés aux 76
104 Hypothalamus Kisspeptine Norépinéphrine Opioïdes endogènes Neurones à GnRH GnRH libérée dans la circulation porte hypothalamohypophysaire Libération pulsatile de GnRH Antéhypophyse RGnRH Cellules gonadotropes Fsh Rgnrh Lh gsu LH FSH GSU FSH Stéroïdes LH Stéroïdes sexuels Gamètes Ovocytes, spermatozoïdes : Des neurosécrétions hypothalamiques pulsatiles de GnRH régulent endocrine de en passant par le système porte hypothalamo-hypophysaire. Le récepteur de la GnRH (R-GnRH) est exprimé à la surface des cellules gonadotropes. La GnRH se fixe sur son récepteur et relaie les informations du système nerveux central induisant la synthèse et la sécrétion des gonadotropines, LH et FSH, importantes dans le contrôle de la reproduction humaine. Ces hormones sont libérées dans la circulation sanguine et stimulent la production des stéroïdes sexuels et des gamètes. De nombreux signaux des systèmes nerveux central et périphérique sont impliqués dans la modulation de et dans la sécrétion des neurones à GnRH. Certains de ces signaux sont activateurs comme la kisspeptine, la norépinéphrine ou le neuropeptide Y, inhibiteurs avec les opioïdes endogènes, -I, la progestérone ou possèdent les deux actions (activation et inhibition) tel que (E2). La GnRH elle-même exerce une boucle de régulation sur sa sécrétion les thèses de A.L Shang, 2011 et N. Benz, 2013 (tel )
105 proté C-terminale intracytoplasmique impliquée dans la désensibilisation des récepteurs (Kaiser et al., 1997 ; Hislop et al., 2000). 1. Isoforme de type I (GnRH-I) La GnRH est un décapeptide de formule pglu-his-trp-ser-tyr-gly-leu-arg-pro-gly- NH 2 dont la structure a été découverte en 1971 (Baba et al., 1971 ; Schally et al., 1971). Elle a une masse moléculaire théorique de 7,8 kda. Elle est courbée au niveau de la glycine 6 et ses extrémités C- et N-terminales ont un rôle important pour son interaction avec son récepteur (). Son rôle clé é élucidé plus tard (Conn et Crowley, 1994 ; Millar, 2005). chromosome est organisé en quatre exons et trois introns (Seeburg et al., 1987 ; Hayflick et al., 1989). Il code pour une pré-hormone de 92 acides aminés qui va être clivée puis modifiée dans des granules de sécrétions. Le précurseur contient : Un peptide signal (23 acides aminés) permettant le «packaging» et la sécrétion Le décapeptide GnRH Un site de clivage protéolytique de trois acides aminés (Gly-Lys-Arg ou GKR) reconnu par une pro-hormone convertase 56 acides aminés constituant la protéine associée à la GnRH (GAP), sécrétée avec la GnRH mais dont la fonction est pas encore élucidée -traductionnelles nécessaire à son activité hormonale : la cyclisation en pyroglutamate de la glutamine en N-terminale et amidation de la glycine C- terminale. La GnRH est synthétisée dans les neurones hypothalamiques qui la sécrètent de manière pulsatile et la libèrent dans le système porte hypophysaire vers antérieure. Elle se fixe sur son récepteur au niveau des cellules gonadotropes pour stimuler et permettre la libération des gonadotropines LH et FSH, qui régulent la gamétogénèse et la stéroidogénèse (Conn et Crowley, 1994 ; Millar, 2005). La pulsatilité de la libération de la 77



106 Acides aminés Acides aminés importants pour la fixation sur le récepteur et son activation Acides aminés importants pour la fixation sur le récepteur uniquement La molécule est courbée au niveau de aminé glycine en position 6. La substitution de cet acide aminé stabilise la conformation, augmente de pour le récepteur et diminue son catabolisme. Cette caractéristique est utilisée pour produire tous les agonistes et antagonistes de la GnRH. Les queues C- et N-terminales sont impliquées dans la fixation de la GnRH sur le récepteur mais seulement la queue C-terminale est associée à du récepteur. Millar, 2005
107 Depuis son identification en 1971, trois isoformes de la GnRH ont été caractérisées 2009). 2. Isoforme de type II (GnRH-II) la GnRH-II elle est codée par un gène localisé sur le chromosome 20 et présente acides aminés sont modifiés : His 5, Trp 7, Tyr 8 (Chen et al., 1998 ; White et al., 1998 ; Leung et al., 2003) (). La GnRH-II est distribuée dans le système nerveux central où elle semble agir comme un neuromodulateur des comportements sexuels ainsi que dans le système nerveux périphérique (Millar, 2005). Elle est aussi exprimée dans de nombreux tissus périphériques tels que les reins, la prostate ou le placenta et dans les tumeurs dérivant de ces tissus (Cheon et al., 2001 ; Millar, 2003). 3. Isoforme de type III (GnRH-III) Une autre isoforme, appelée GnRH-III, a été isolée uniquement chez la lamproie (Petromyzon marinus) où elle joue un rôle essentiel dans le contrôle des fonctions reproductrices au niveau de la gamétogénèse et de la stéroidogénèse (Deregon et Sower, 1994). avec la GnRH de type I. Quatre acides aminés sont modifiés : His 5, Asp 6, Trp 7, Lys 8 par rapport à la GnRH-I (Sower et al., 1993) (). Chez elle se fixe sur le R-GnRH de cellules cancéreuses et exerce un fort pouvoir antiprolifératif (Limonta et al., 2012). fonction reproductive chez les mammifères (Wray et Hoffman,1986) grâce à la coordination de la synthèse et de la sécrétion pulsatile de GnRH (Merchenthaler et al., 1984). souris. Cependant la conservation du processus de migration des neurones a été montrée chez mbryon précoce, les neurones à GnRH sont retrouvés dans la région nasale, au niveau les placodes olfactives, puis migrent et al., 2007). La migration des 78
108 neurones à GnRH est associée au système olfac détectés dans les placodes olfactives entre 5,5 et 6 semaines de développement embryonnaire, puis ils commencent à migrer le long des nerfs olfactifs de la région vasoméronasale pour entrer dans le prosencéphale à 6,5 semaines de développement et semaines. Leur migration est considérée complète entre la 13 ème et la 16 ème semaine de grossesse (Quinton et al., 1997). Cependant, leur maturation complète avec des connexions synaptiques et une fonction correcte est atteinte à la puberté (Kim et al., 1999). Les neurones à GnRH sont soumis à une régulation fine pour synchroniser la libération pulsatile de dans le système porte hypophysaire et induire la sécrétion correcte des gonadotropines (Tsutsumi et Webster, 2009). Une libération pulsatile de LH et de FSH semble être obligatoire pour la fonction des neurones à GnRH hypothalamiques. Les pulses de GnRH se produisent environ toutes les 2h omme adulte. Chez la femme cette libération varie de la puberté à la ménopause et pendant le cycle ovulatoire. De nombreux signaux des systèmes nerveux central et périphérique sont impliqués dans la modulation de tion des neurones à GnRH (Hrabovszky et Liposits, 2013). Certains de ces signaux sont activateurs comme la kisspeptine, la norépinéphrine ou le neuropeptide Y, inhibiteurs avec les opioïdes endogènes, - I, la proges (E2). En effet, chez, GnRH sauf à la fin de la phase folliculaire précédent le pic de LH et de FSH chez la femme où elle exerce un rétrocontrôle positif. De plus la GnRH elle-même exerce une boucle de régulation sur sa sécrétion (DePaolo et al., 1987) (). Le R-GnRH a été cloné pour la première fois en 1992 dans la lignée cellulaire murine gonadotrope -1 (Tsutsumi et al., 1992). La découverte de cette séquence a permis la (Kakar et al., 1992 ; Perrin et al., 1993). -GnRH hypophysaire est un peptide 79
, pour la structure du récepteur ou pour la poche de fixation (en vert) sont représentés.")
109 Milieu extracellulaire Glycosylation Milieu intracellulaire Le R-GnRH humains comprend sept domaines transmembranaires reliés par 3 boucles extracellulaires et 3 boucles intracellulaires. Les acides aminés indispensables pour la liaison de la GnRH (en rouge), pour la structure du récepteur ou pour la poche de fixation (en vert) sont représentés. Les résidus impliqués dans et dans le couplage aux protéines G sont notés en bleu et en orange respectivement. Les sites potentiels de phosphorylation par les protéines kinases A et C (PKA et PKC) sont aussi notés. Les interactions intermoléculaires entre les acides aminés du R-GnRH et de la GnRH sont aussi indiquées via les lignes rouges. la revue de Millar et al., 2008
110 de 328 acides aminés codé par un gène localisé sur le chromosome 4 à la position 4q13 ant sur 18,9 kilobases (kb). Il est composé de trois exons et de deux introns (Fan et al., 1994) et partage domaines transmembranaires. Il appartient à la famille des rhodopsin-like récepteurs couplés aux protéines G. Il est composé de sept domaines transmembranaires reliés par six boucles intracellulaires et un domaine extracellulaire de 35 acides aminés contenant deux sites de glycosylation (Kakar et al., terminale (1 ou 2 acides aminés) contrairement aux autres récepteurs appartenant à cette famille. La queue C-terminale joue un rôle clé pourquoi le R-GnRH est internalisé relativement lentement et ne subit pas une désensibilisation rapide. Des expériences de mutagénèses dirigées G. Ces études ont aussi permis de révéler des sites potentiellement phosphorylables par les protéines kinases A et C (PKA et PKC) dans les boucles intracytoplasmiques (pour revue Millar et al., 2008) (). Dans les cellules gonadotropes les voies de signalisation intracellulaires activées suite à la fixation de la GnRH ou de ses analogues sont bien connues. Une fois la GnRH fixée, le récepteur se couple à la sous-unité q/11 de la protéine G (G q/11) qui active la phospholipase conduisant à lulaire de diacylglycérole (DAG) et -triphosphate (IP 3) suite à la transformation de phosphatidylinositol 4,5- bisphosphate (PIP2). Le DAG active la PKC 3 induit la libération du Ca 2+ des stocks intracellulaires. La PKC déclenche la cascade de signalisation des MAP Kinases (mitogenactivated protéine kinase) initiant -unités et des gonadotropines ainsi que les MAPK telles que les protéines ERK, JNK et p38 MAPK (Naor, 2009) (). Les phospholipases D et A2 sont aussi activées par la fixation du ligand sur son récepteur entraînant une activation supplémentaire et prolongée de PKC (Naor, 1991 ; Kraus et al., 2001). La voie de signalisation activée par G s/ampc est aussi impliquée dans la réponse initiale des cellules gonadotropes à la GnRH (ou à ces analogues) et assure le lien pour la transmission du signal entre le récepteur situé à la membrane plasmique et le noyau, amenant à la synthèse et à la sécrétion des gonadotropines (Liu et al., 2002) (). 80

 est déclenchée par le couplage du récepteur avec la protéine G s ainsi que par les isoformes nouvelles de la PKC. Un couplage avec la protéine G i a aussi été suggéré.")
111 Membrane plasmique R-GnRH Cytoplasme PIP 2 q s i PLC- AC IP 3 DAG 1 AMPc Ca 2+ PKC/PKC PKA JNK MAPK/ ERK1/2 JNK1/2, p38mak, ERK1/2, c-src Autres kinases MEKK1, cdc42, c-src Egr1 Jun ELk LH Noyau La fixation de la GnRH sur son récepteur induit le couplage du récepteur avec la protéine G q11 activant la PLC qui clive le PIP 2 en DAG et IP 3. Ces seconds messagers déclenchent la PKC et la libération de Ca 2+ des réserves du RE. de cyclase (AC) est déclenchée par le couplage du récepteur avec la protéine G s ainsi que par les isoformes nouvelles de la PKC. Un couplage avec la protéine G i a aussi été suggéré. de ses voies induisent la production et le libération des gonadotropines LH et FSH. Adaptée de Kakar et al., 2004 ; Aguilar-Rojas et Huerta-Reyes, 2009
112 il a été rapporté que la GnRH-II et GnRH-III (retrouvée chez la lamproie) se fixent sur le R-GnRH dit classique de type I pour agir (Montagnani Marelli et al., 2015). Chez les mammifères, la GnRH de type III semble exercer un effet anti-tumoral sur les cellules cancéreuses exprimant le R-GnRH tout en limitant la sécrétion de gonadotropines (Kovacs et al., 2002). Outre le rôle bien établi du couple GnRH/R-GnRH au niveau pituitaire dans la régulation des gonadotropines, la détection de l'hormone et de son récepteur dans de multiples tissus normaux ou tumoraux suggère des rôles extra-hypophysaires pour la GnRH et son récepteur. Ils peuvent avoir des activités autocrine, paracrine et/ou endocrine. Ils ont été retrouvés dans les tissus normaux reproducteurs féminins (ovaire, endomètre, myomètre, placenta) (Tureck et al., 1982 ; Imai et al., 1996 ; Currie et al., 1981) et masculin (testicules, prostate) (Kakar et al., 1992) ainsi que dans des tissus non reproducteurs au niveau des systèmes immunitaire (Chen et al., 1999), du sein (Kakar et al., 1992), du foie, du pancréas, (Sion-Vardi et al., 1992 ; Kakar et Jennes, 1995). Leur présence a aussi été détectée dans des tissus et des lignées cellulaires tumorales (pour revue : Aguilar-Rojas et Huerta-Reyes, 2009). La GnRH a différentes activités suivant le tissu ion neuronale au cours du développement, la neuromodulation au niveau du cerveau pouvant affecter le comportement sexuel, une possible modulation au niveau de la vision, l'inhibition de la sécrétion de l'acide gastrique, le chimiotactisme des lymphocytes T, la libération de l'hormone chorionique gonadotrophique (hcg) dans le placenta, la stéroïdogénèse, la prolifération des cellules de mélanome et l'inhibition de la croissance des tumeurs (Kakar et Jennes, 1995 ; Hapgood et al., 2005). La présence du R-GnRH a aussi été montrée dans les tissus respiratoires de différentes espèces animales et dans des lignées cellulaires dérivant de cancers des voies aériennes humaines, mais cela reste très peu décrit pour le tissu normal humain. Le récepteur a été mis en évidence dans le tissu nasal bovin, le tissu pulmonaire et trachéal de rat (Koushik et al., 2004 ; Sundaram et al., 2009a ; Koushik et Kompella, 2004). Sundaram et collaborateurs -GnRH dans le cornet nasal inférieur et moyen chez le bovin, en 2009 (Sundaram et al., 2009a). Ces études incluraient le R-GnRH dans le transport vectoriel Koushik et collaborateurs, en 2004, a -GnRH dans la lignée Calu- adénocarcinome pulmonaire humain (Koushik et al., 2004). Enfin, a été établie dans des lignées cellulaires cancéreuses de poumons (A549 et H1299; Sundaram 81
113 Ligand Conformation du récepteur Voies de signalisations activées Protéine G monomérique des Production et libération de LH et FSH Libération pulsatile de GnRH Antiprolifération et apoptose Adhésion cellulaire et migration. Suite à la fixation du ligand, le récepteur change de conformation et déclenche des voies de signalisations différentes suivant et cellulaire (SC1-4). Millar et Pawson, 2004
114 et al., 2009b et Taratula et al., 2011). Cependant, les rôles spécifiques de l'hormone et de son récepteur dans ces différents tissus ou lignées cellulaires restent à être élucidés. Les récepteurs hypophysaires des cellules gonadotropes et des tissus périphériques diffèrent pour les voiormone (Neill, 2002 ; Millar et al., 2008). Les R-GnRH hypophysaires sont associés à la sous unités G q/11 et G des protéines G (Cheng et Leung, 2005 ; Naor, 2009 ; Liu et al., 2002). En revanche, R-GnRHs périphériques dans les tissus cancéreux ou les lignées dérivées de cancers sont couplés aux voies de signalisation activées par la sous-unité G i. Cet évènement induites par les MAPKs (MAPK p38, ERK1/2, et JNK), la phosphatidylinositol-3-kinase (PI3K) et la phosphotyrosine phosphatase (pour revue : Limonta et al., 2012). Les réponses des récepteurs périphériques aux molécules pharmacologiques apparaissent différentes de celles retrouvées au niveau des récepteurs hypophysaires. Les analogues de la GnRH, qui ont un effet antagoniste au niveau hypophysaire (comme Cetrorelix) possèdent un effet agoniste dans les tissus périphériques surtout dans le cas de cancers (Kleinman et al., 1994 ; Yano et al., 1994). étaient dues à des différences moléculaires entre les récepteurs hypophysaires et périphériques. En effet, même si les séquences des transcrits du récepteur sont identiques les protéines correspondantes peuvent connaître des modifications post-traductionnelles (glycosylation ou phosphorylation) générant des protéines différentes (Rama et Rao, 2001). L du messager et de la protéine du R-GnRH a été vérifiée dans des lignées cellulaires placentaires (Lin et al., 1995) puis dans les tissus reproducteurs, mâles ou femelles, hypophysaire (Chi et al., 1993 ; Kakar et al., 1994 ; Limonta et al., 1999, 2012). Millar et son équipe, en 2008, ont proposé la théorie de la «signalisation sélective induite par le ligand» pouvant expliquer les différences observées. Un type de ligand stabilise une certaine conformation du R-GnRH et active une voie intracellulaire. Chaque analogue (agoniste ou antagoniste) peut se fixer au récepteur de manière sélective pour activer des voies intracellulaires différentes et spécifiques, déclenchant des effets biologiques différents (Millar et al., 2008) (). La connaissance de ces mécanismes est importante afin de bien comprendre les actions à la fois physiologiques et thérapeutiques de la GnRH et de son récepteur et ainsi pouvoir généraliser l'utilisation des analogues de la GnRH en endocrinologie et en thérapie anticancéreuse (Hapgood et al., 2005). 82
115 Le R-GnRH est très fortement exprimé dans de nombreux tissus extra-hypophysaires. La détection du récepteur dans ces tissus suggère que la GnRH synthétique et les analogues de la GnRH (agonistes ou antagonistes) pourraient avoir une action directe sur des cibles thérapeutiques périphériques. En thérapeutique, la GnRH synthétique est administrée défauts hormonaux (Pitteloud et al., 2002). Elle peut aussi être prescrite en cas de pathologies liées à la fertilité chez les p mentale. Elle est aussi administrée aux patients avec un cryptorchidie ou un retard pubertaire (Millar et Newton, 2013). La GnRH synthétique à un temps de demi-vie très court (entre 2 et 4 minutes) principalement dû à la dégradation de la liaison entre les acides-aminés de nombreux analogues de la GnRH ont été développés et peuvent être divisés en deux classes : les agonistes et les antagonistes. Contrairement à la stimulation par la GnRH synthétique, l (FSH et LH) et des stéroïdes sexuels (Chillik et Acosta, 2001). Les analogues de la GnRH sont très souvent prescrits dans les traitements de maladies hormono-dépendantes (endométriose, syndrome des ovaires polykistiques, fibrose utérine ou ation ainsi que pour des cancers du système reproducteur masculin (cancer de la prostate) ou féminin (cancer du sein Chillik et Acosta, 2001 ; Millar et Newton, 2013). Les agonistes naturel. Ils ont été circulation sanguine. Le domaine C-terminal du décapeptide est crucial pour sa fixation au récepteur et la queue N-terminale ont donc été développés en remplaçant la Gly 6 par un acide aminé avec une chaîne latérale apolaire et volumineuse (Leu, Trp, Ser(tBu), His(Blz)), augmentant la demi-vie dans le sang 83
116 tbu: tert-butyl; Et: ethyl; AzaGly: aza-glycine; Ac: acetyl; Cpa: chlorophenylalanine; Pal: 3-pyridylalanine; Cit: citrulline; Aph: 4- aminophenylalanine; Hor: L-hydroorotyl; Cba: carbamoyl; ipr: isopropyl; Nal: 2-naphtylamine; harg: homoarginine
117 et certains des analogues ont en plus un résidu éthylamide sur la proline en position 9 (Pro 9 ), augmentant leur affinité pour le récepteur (Engel et Schally, 2007) (). : la triptoréline, le leuprolide, la goséréline et la buséréline. Une forte dose et une administration prolongée des agonistes de la GnRH -gonadique, par la régulation négative des R-GnRHs. Ils sont indiqués dans le traitement de la puberté précoce, traitement des cancers hormono-dépendants et sont surtout administrés par injection souscutanée (Limonta et al., 2012). Pour notre étude nous avons recherché des traitements déjà commercialisés à base a GnRH existant en spray nasal. Deux composés existent : nafaréline acétate (Synarel ) ). Notre choix la buséréline. Ce médicament est prescrit pour le traitement du cancer de la in vitro. Les antagonistes ont capables de bloquer - composés entrent en compétition avec la GnRH pour la fixation sur le récepteur et bloquent le R-GnRH des cellules gonadotropes entraîoutenue de la sécrétion des gonadotropines (Padula, 2005 ; Montagnani Marelli et al., 2006). Les premiers antagonistes produits présentaient une structure complexe avec entraî et des réactions anaphylactiques (Padula, 2005). Des antagonistes dépourvus ont été développés et contiennent en N-terminale la séquence suivante : Ac-D-Nal-D-Cpa-D- Pal (Ac=Acetyl, Nal=2-naphtylamine, Cpa=chlorophenylalanine, Pal=3-pyridalanine) et une D- Alanine en position 10 de la chaîne en acides aminés. Ils présentent aussi des substitutions à différentes positions 5, 6 et 8 (Herbst, 2003) ( ). Certains antagonistes, ayant un fort pouvoir inhibiteur et une forte affinité pour le récepteur, sont commercialisés: la cetrorelix, ganirelix, abarelix et degarelix (Limonta et al., 2012). Les antagonistes de la GnRH sont utilisés par exemple pour assistance à la procréation. 84

118 (A) Domaine formant le canal Domaine kinase (B) Domaine mélastatine (C) (A) le gene humain TRPM7 est localisé sur le chromosome 15 et comprend 39 exons. Les régions correpondantes au canal et au domaine kinase sont surlignés en gris sur la representation du gène. (B) Le gène code pour une protéine de 1863 acides aminés comprenant 6 hélices transmembranaires. La queue N-terminale cytoplasmique contient un domaine très conservé chez les canaux TRP de la sous-famille mélastatine (domaine mélastatine). TRPM7 présente un domaine kinase en C-terminale qui possède de nombreux sites. (C) TRPM7 est une protéine multimérique dont le pore est formé par les boucles entre les helices transmembranaires 5 et 6 de chaque monomère. Paravicini et al., 2012
119 La protéine CFTR est régulée par le Mg 2+ intracellulaire (Bargon et al., 1992 ; Li et al., 1996 ; Ikuma et Welsh, 2000). La régulation du CFTR par le Mg 2+ sera détaillée dans la partie La concentration intracellulaire de Mg 2+ ([Mg 2+ ] i) est principalement régulée par un canal ubiquitaire appelé TRPM7 (Transient Receptor Potential Melastatin 7) (Ryazanova et al., 2010). Ce canal aussi appelé TRP-PLIK (Transient receptor potentialphospholipase c-interacting kinase, Runnels et al., 2001), ChaK1 (channel-kinase 1, Ryazanov, 2002) ou LTRPC7 (long transient receptor potential channel 7, Nadler et al., 2001) appartient à la famille des TRP (Transient receptor potential). C, cette famille de protéine comprend 27 isoformes réparties, suivant leurs homologies de séquences, en 6 sousgroupes nommés : TRPC (Canonical, 7 isoformes), TRPV (Vanilloid, 6 isoformes), TRPM (Melastatin, 8 isoformes), TRPA (Ankyrin, 1 isoforme), TRPML (Mucolipin, 3 isoformes) et TRPP (Polycistic, 3 isoformes) (Nilius et Owsianik, 2011). Ils permettent la transduction de messages sensoriels (Clapham, 2003 ; Daman et al., 2008, Nilius et Owsianik 2010 ; Venkatachalam et Montell, 2007). Le gène TRPM7 se trouve sur le bras long du chromosome 15 en position 21 et comprend 39 exons (). Il a été identifié et cloné pour la première fois en 2001, dans le cerveau murin (Runnels et al., 2001), comme étant un acteur clé dans la régulation de Mg 2+ intracellulaire. Il code une protéine appelée TRPM7 de 1863 acides aminés pour un poids moléculaire évalué à environ 212 kda (Asrar et Aarts, 2013). TRPM7 est un canal perméable aux cations divalents dont la perméabilité aux différents ions est la suivante : Zn 2+ Ni 2+ >>Ba 2+ >Co 2+ >Mg 2+ Mn 2+ Sr 2+ Cd 2+ Ca 2+ (Monteilh- Zoller et al., 2003). Tout comme son homologue TRPM6, TRPM7 possède un domaine de type ser/thr-kinase qui leur est caractéristique (Nilius et Owsianik, 2011) ( Le canal TRPM7 est composé de 4 sous-unités comprenant 6 hélices transmembranaires reliées par des boucles intra- et extra-cellulaires. Les boucles entre les hélices 5 et 6 forment le pore ( )., ce canal est ubiquitaire mais il possède une différence les tissus adipeux (Fonfria et al., 2006 ; Nadler et al., 2001). Etant un canal ionique avec une activité enzymatique, TRPM7 est appelé chanzyme (Montell, 2003 ; Asrar et Aarts, 2013). 85
120 TM1: TM2: TM3: TM4: TM5: TM6: Milieu extracellulaire Domaines transmembranaires Boucles formant le pore Milieu intracellulaire Domaine TRPM conservé Domaine TRP Sites possibles de fixation de PIP 2 K1112, R1115,K1125 N-term Régions possibles de fixation de PIP 2 : et Intéragit avec les isoformes de la PLC C-term Région superenroulées: Cibles (in vitro): Annexine-A1 Myosine isoforme II Tropomoduline PLC1 Domaine -Kinase: Résidus clés pour la catalyse: K1648, D1775 TRPM7 a besoin de PiP 2 pour fonctionner mais les mécanismes moléculaires impliqués dans de TRPM7 par le PiP 2 ne sont pas définis. TRPM7 est directement associé à actomyosine du cytosquelette de manière kinase dépendante suggérant que TRPM7 régule la dynamique de cette protéine par phosphorylation. -A1 a aussi été identifié in vitro comme substrat de TRPM7. Visser et al., 2014
121 Dans les cellules de mammifères, le domaine kinase de TRPM7 phosphoryle des protéines du cytosquelette (Clark et al., 2006, 2008a). essentielle pour son association avec le cytosquelette. En effet, l région riche en sérine/thréonine localisée en N-terminal du domaine catalytique de TRPM7 semble nécessaire au recrutement des substrats du domaine kinase du canal (Clark et al., 2008b). migration (Clark et al., 2008b). La phosphorylation de la myosine IIA par TRPM7 stabilise les filaments et leur localisation cellulaire (Clark et al., 2008b) (). Dorovkov et Ryazanov ont identifié, in vitro, annexine A1 (AnxA1) comme substrat de TRPM7 (Dorovkov et Ryazanov, 2004 ; Dorovkov et al., 2011). Les annexines sont des protéines liant le Ca 2+ qui interagissent avec les phospholipides de la bicouche membranaire. ion et : Gerke et Moss, 2002 ; Hanano et al., 2004 ; Aarts et al., 2003). TRPM7 serait impliquée dans la prolifération et la mort cellulaire (Dorovkov et Ryazanov, 2004). L AnxA1 par TRPM7 déstabilise son interaction avec la membrane (Dorovkov et al., 2011) (). Le canal TRPM7 est impliqué dans de nombreux processus physiologiques tels que la mais aussi pathologiques comme par exemple dans la mort neuronale et les cancers (Aarts et al., 2003 ; Monteilh-Zoller et al., 2003, Chubanov et al., 2012, Mandavilli et al., 2012, Tashiro et al., 2013, Zhang et al., 2014). cationique de TRPM7, dans les différents processus cellulaires, des protéines TRPM7 mutantes ou des inhibiteurs du canal TRPM7 tels que la waixenicine A sont utilisés (pour revue : Visser et al., 2014). De nombreux composés inhibant les courants dépendants de TRPM7 ont été décrits comme par exemple le carvacrol ou les cations trivalents (La 3+, Gd 3+ ) ( ) canaux (Chubanov et al., 2014). Zierler et collaborateurs, en 2011, utilisent la waixenicine A et la prolifération cellulaire. 86


122 * IC50 values were shown for recombinant TRPM7 currents measured in the absence of internal Mg2+. The dose-dependent effect of spermine was evaluated on endogenous TRPM7 currents in divalent-free external solution. n.d. - not determined. Chubanov et al., 2014 Chubanov et al., 2014
123 al., 2011). 20 molécules organiques activatrices de TRPM7 ont aussi été identifiées comme par exemple le naltriben (Hofmann et al., 2014, Chubanov et al., 2014) (). Les premières études du canal TRPM7 en 2001 suggéraient que le domaine kinase al., 2001 ; Runnels et al., 2001). Cependant, les gating» de TRPM7 en régulant sa sensibilité au Mg 2+ et au Mg-ATP (Demeuse et al., 2006; Matsushita et al., 2005; Schmitz et al., 2003 peut réguler de manière indirecte la fonction de TRPM7 al., TRPM7 pourrait aussi influencer le «gating» du canal par un mécanisme similaire à celui décrit pour son homologue TRPM6 (van der Wijst et al., 2014). La dimérisation des séquences - par ce canal (van der Wijst et al., 2014) (Crawley et Cote, 2009). Les mécanismes moléculaires impliqués encore totalement élucidés (Chubanov et al., 2017 ; pour revues : Visser et al., 2014 ; Paravicini et al., 2012). Cependant, deux modèles de régulation prédominent. Le premier concerne la régulation de TRPM7 par le Mg-ATP et le Mg 2+ qui se fixent au niveau de la queue C-terminale sur deux sites : un au niveau de site catalytique et un autre en dehors (Demeuse et al., 2006 ; Schmitz et al., 2003). Cette fixation du Mg 2+ inhibe les courants médiés par le canal TRPM7. Le deuxième modèle prédit une régulation de TRPM7 par le PiP 2 intracellulaire (Runnels et al., 2002). diminution de PiP 2 du PiP 2 en DAG et IP 3. Cette déplétion PiP 2 inhibe le canal TRPM7 même en absence de Mg 2+ (Runnels et al., 2002). Il a été suggéré que le Mg 2+ et PiP 2 interagissaient par un mécanisme commun pour supprimer les courants dépendant de TRPM7 (Kozak et al., 2005). Une autre hypothèse concernant la régulation de TRPM7 existe proposant la formation TRPM7 et TRPM6. La phosphorylation de TRPM7 par le domaine kinase de TRPM6 affecte la sensibilité de TRPM7 pour le Mg-ATP (Brandao et al., 2014 ; Zhang et al., 2014). 87
124 sont associées à de nombreuses pathologies dont quelques exemples vous seront cités dans la suite du chapitre. TRPM7 est impliqué dans la prolifération, la migration cellulaire ou la transition épithélio-mésenchymateuse qui sont dérégulés en cas de cancer. Son rôle dans le développement tumoral a été mis en évidence dans de nombreuses lignées cellulaires dérivant de cancers mais aussi chez des patients présentant par exemple un adénocarcinome du sein (Dhennin-Duthille et al., 2011), un adénocarcinome pancréatique (Rybarczyk et al., 2012) ou un cancer de la prostate (Sun et al., 2013). Une surexpression du canal TRPM7 est observée dans les cancers cités ci-dessus. Un rôle de TRPM7 dans les accidents ischémiques a été mis en évidence in vitro dans des cultures de cellules neuronales (Aarts et al., 2003) et in vivo chez la souris (Sun et al., 2009). ROS et accumulent du Ca 2+ sion de TRPM7 est supprimée, les réserves de Ca 2+, la production de ROS et la mort cellulaire sont diminuées (Aarts et al., 2003). Chez la souris, in vivo neurones de la mort cellulaire après une ischémie cérébrale (Sun et al., 2009). Une implication de TRPM7 dans les formes familiales de proposée préséniline 1 (associée aux formes familiales de la mal PiP 2 et al., 2006 ; Oh et al., 2012). TRPM7 joue un rôle dans la fonction cardiovasculaire. Les courants médiés par TRPM7 dans les fibroblastes cardiaques sont plus importants chez les patients présentant une fibrillation atriale par rapport aux patients avec un rythme sinusoïdal normal (Du et al., 2010). De plus TRPM7 libère un facteur de croissance (TGF-, Transforming growth factor beta 1) impliqué dans la fibrose cardiaque (Du et al., 2010). Des études sur de lignées cellulaires hépatiques de rat ont montré un rôle clé de et al., 2013). Les cellules stellaires sont importantes pour la sécrétion, le dépôt protéines matricielles et jouent un rôle prépondérant dans la fibrose du foie (Hernandez-Gea et Friedman, 2011 ; Bian et al., 2012). La diminution de la prolifération des cellules stellaires hépatiques de rat (HSC-T6) suite tion de TRPM7 (Fang et al., 2013) suggère un traitement potentiel pour la fibrose hépatique. 88
125 89
126 : Stimulation de cellules épithéliales bronchiques humaines par la GnRH : CFTR. La mucoviscidose est due à une mutation du gène CFTR (Cystic Fibrosis Transmembrane Conductance Regulator) codant la protéine CFTR. Cette protéine est principalement un canal chlorure (Cl - ) AMPc- dépendant. Elle est localisée dans la membrane apicale de cellules épithéliales. La mutation F508del est la plus fréquente et Cette délétion entraîne un défaut de maturation de la protéine qui est retenue dans le réticulum endoplasmique par le système de contrôle qualité du RE (ERQC) Cependant, une faible quantité de protéines malformées échappe à ce système de contrôle et parvient à la membrane plasmique où elle semble avoir une activité canal Cl - malgré une stabilité réduite. de molécules pouvant améliorer la conformation la stabilité de la protéine CFTR-F508del à la membrane plasmique est un axe majeur dans le développement de thérapies pour la mucoviscidose et la recherche de nouveaux partenaires de la protéine CFTR. Une interaction directe entre le canal CFTR et a été mise en évidence au laboratoire. 2+ et ATP-et al., 2007 ; Le Drévo et al., 2008). La suite de ces travaux a consisté à -wt et CFTR-F508del (Le Drévo et al., a été corrélée à une augmentation des efflux I - et Cl - ), AMPc-dépendant, qui serait due à une expression membranaire du CFTR plus importante (Le Drévo et al., 2008). Il a donc été émis comme -AnxA5 stabiliserait le CFTR à la surface cellulaire et influerait les sécrétions ioniques médiées par le canal. Au vu de ces résultats, semble être une cible potentielle pour corriger causées par la mutation F508del. e capable 90
127 littérature la GnRH se présente comme un candidat potentiel. En effet, plusieurs études décrivent un lien entre les voies intracellulaires activées par la liaison de la GnRH sur son (Shibata et al., 1997 ; Yamamoto et al., 2001 ; Kawaminami et al., 1998, 2002a, 2002b, 2003 ; Yao et Kawaminami, 2008 ; Kawaminami et al, 2008). importante est observée après traitement par un analogue de la GnRH dans la lignée cellulaire utérins humains (Shibata et al., 1997 ; Yamamoto et al., 2001). La modulation de la synthèse cellules du tractus génital (Kawaminami et al., 1998, 2002a, 2002b, 2003 ; Yao et participe à la transduction du signal déclenchée par la fixation de la GnRH ou de ces analogues sur le R-GnRH, modulant la synthèse et la sécrétion des gonadotropines ou la stéroïdogénèse. Le déclenchement de la voie des MAPKs de type ERK (notamment ERK 1/2) associée au R-GnRH active la régulation et al, 2008). Cette hormone semble donc être une piste Le R-GnRH est exprimé dans de nombreux tissus extra-hypophysaires et dans différentes lignées cellulaires des voies aériennes dérivées de tumeurs (Calu-3, A549 et H1299). Dans un premier temps il a fallu vérifier lde -GnRH dans les lignées cellulaires dérivées humain exprimant le CFTR-wt (16HBE et CFBE41o-/wt) ou le CFTR-F508del (CFBE41o- et CFBE41o-/F508del), puis tester les effets de la GnRH et la fonction du canal CFTR. basale des messagers -GnRH a été vérifiée par PCR. La technique de western blot a permis de mettre en évidence la présence des protéines, AnxA5 et R-GnRH, dans les quatre lignées cellulaires (16HBE14o-, CFBE41o-/wt, CFBE41oet CFBE41o-/F508del). La GnRH se fixe sur son récepteur pour activer les voies de signalisations intracellulaires, nos modèles cellulaires exprimant le R- peut être étudiée. Plusieurs concentrations de GnRH ont été testées. Des effets sur ont été observés entre 10-9 et 10-7 M. Pour travailler à la concentration la plus faible possible, la concentration de GnRH à 10-9 M a été sélectionnée. Ces résultats ne sont pas présentés dans le papier. 91
128 -9 M) sur la quantité 5, une cinétique 180 et 310 minutes) a été effectuée sur les modèles cellulaires. Une augmentation significative protéique dans les quatre modèles cellulaires, après pendant 60 minutes. été testé par les significative des transports ioniques dépendant du CFTR dans les lignées exprimant le CFTR sauvage ou muté par rapport aux cellules non traitées. Pour apprécier membranaire du canal CFTR des ie ont été effectuées et ont permis de corréler cette augmentation des efflux ioniques à une augmentation du nombre de canaux CFTR insérés dans la membrane plasmique. R-GnRH a pour la première fois été mise en évidence dans des lignées cellulaires épithéliales bronchiques humaines non dérivées de tumeurs. Un traitement à la GnRH (1h à 10-9 M) déclenche une augmentation d dans les cellules épithéliales bronchiques exprimant le CFTR sauvage ou muté lignées cellulaires épithéliales bronchiques humaines exprimant le CFTR sauvage ou muté F508del avec la GnRH conduit à une augmentation des efflux ioniques médiés par le CFTR, due à une quantité plus importante de canaux CFTR exprimés à la membrane plasmique. Cette observation utilisation de la GnRH ou ces analogues pour restaurer les courants Cl - dépendant du CFTR dans les cellules CF. 92
129 Improvement of Chloride Transport Defect by Gonadotropin-Releasing Hormone (GnRH) in Cystic Fibrosis Epithelial Cells Nathalie Benz 1,2, Sophie Le Hir 1,3, Caroline Norez 4, Mathieu Kerbiriou 1,5, Marie-Laure Calvez 1,5, Frédéric Becq 4, Pascal Trouvé 1 *, Claude Férec 1,3,5,6 * 1 Institut National de la Santé et de la Recherche Médicale, UMR1078, Brest, France, 2 Association Gaetan Saleun, Brest, France, 3 C.H.U. Brest, Hôpital Morvan, Laboratoire de Génétique Moléculaire, Brest, France, 4 Institut de Physiologie et Biologie Cellulaires, Centre national de la recherche scientifique FRE 3511, Université de Poitiers, Poitiers, France, 5 Université de Bretagne Occidentale, Faculté de Médecine et des sciences de la santé, Brest, France, 6 Etablissement Français du Sang - Bretagne, Brest, France Abstract Cystic fibrosis (CF), the most common autosomal recessive disease in Caucasians, is due to mutations in the CFTR gene. F508del, the most frequent mutation in patients, impairs CFTR protein folding and biosynthesis. The F508del-CFTR protein is retained in the endoplasmic reticulum (ER) and its traffic to the plasma membrane is altered. Nevertheless, if it reaches the cell surface, it exhibits a Cl 2 channel function despite a short half-life. Pharmacological treatments may target the F508del- CFTR defect directly by binding to the mutant protein or indirectly by altering cellular proteostasis, and promote its plasma membrane targeting and stability. We previously showed that annexine A5 (AnxA5) directly binds to F508del-CFTR and, when overexpressed, promotes its membrane stability, leading to the restoration of some Cl 2 channel function in cells. Because Gonadotropin-Releasing Hormone (GnRH) increases AnxA5 expression in some cells, we tested it in CF cells. We showed that human epithelial cells express GnRH-receptors (GnRH-R) and that GnRH induces an AnxA5 overexpression and an increased Cl 2 channel function in F508del-CFTR cells, due to an increased stability of the protein in the membranes. Beside the numerous physiological implications of the GnRH-R expression in epithelial cells, we propose that a topical use of GnRH is a potential treatment in CF. Citation: Benz N, Le Hir S, Norez C, Kerbiriou M, Calvez M-L, et al. (2014) Improvement of Chloride Transport Defect by Gonadotropin-Releasing Hormone (GnRH) in Cystic Fibrosis Epithelial Cells. PLoS ONE 9(2): e doi: /journal.pone Editor: Dominik Hartl, University of Tübingen, Germany Received September 2, 2013; Accepted January 16, 2014; Published February 19, 2014 Copyright: ß 2014 Benz et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited. Funding: The authors thank the association Gaëtan Salaün, the SFR Scinbios (Brest) and the French foundation Vaincre la Mucoviscidose for their financial support. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript. Competing Interests: The authors have declared that no competing interests exist. * pascal.trouve@univ-brest.fr (PT); claude.ferec@univ-brest.fr (CF) Introduction Cystic fibrosis (CF) is the most common autosomal recessive disease in Caucasians. It is caused by mutations in the gene encoding the cystic fibrosis transmembrane conductance regulator (CFTR) protein [1]. CFTR which is a member of the ATPbinding cassette (ABC) transporter superfamily, functions as an ion channel [1,2]. It is mostly expressed in the apical membrane of epithelial cells and helps to maintain the fluid and electrolyte balance across the cell membrane. CFTR comprises two membrane-spanning domains (MSDs), two nucleotide-binding domains (NBDs) and a regulatory domain (RD). The CFTR protein undergoes a complex biosynthetic pathway in the endoplasmic reticulum (ER) in which molecular chaperones and co-chaperones are involved [3]. Wild-type CFTR (Wt-CFTR) biogenesis initiates in the ER where the protein is coreglycosylated, leading to an immature precursor form known as band B (,145 KDa). It further undergoes maturation and glycosylation through the Golgi, originating a complex mature form (band C,,170-kDa) [4]. Only 25% to 70% of the precursor Wt-CFTR matures, depending on the cell type [5]. The remaining part undergoes ubiquitination and degradation by the proteasomal pathway [6,7]. In membranes, once phosphorylated by protein kinase A (PKA) in the R domain, CFTR functions as an ATPgated chloride (Cl 2 ) channel [8]. Among the 1930 described mutations in the CFTR gene ( the deletion of phenylalanine at position 508 (F508del) is the most common one, associated with,70% of CF alleles [9]. The F508del mutation causes a protein folding defect, the nascent protein beeing retained in the ER. As a result, most of the F508del-CFTR channels are degraded intracellularly and very little is targetted to the plasma membrane [10 13]. Nevertheless, in the plasma membrane it exhibits a Cl 2 channel activity despite an open probability 15 times lower than that of Wt-CFTR [14 16]. Furthermore, the F508del-CFTR protein has a faster turnover at the cell surface than Wt-CFTR [17]. In order to overcome the F508del-CFTR defects and provide a potential treatment for CF, potentiators aimed to correct the channel s activity and correctors of the F508del-CFTR trafficking defect have been developed. Potentiators that increase the mutated CFTR s channel activity when it is phosphorylated function via direct binding to modulate the NBDs dimerization or to increase ATP binding and hydrolysis [18 20]. Correctors may act PLOS ONE 1 February 2014 Volume 9 Issue 2 e
130 GnRH Improves Chloride Transport in CF Cells indirectly or by a direct binding to F508del-CFTR. Indirect correctors such as 4-phenylbutyrate [21], glycerol [22], low temperature [23] and miglustat [24] act upon folding, ER retention, degradation and F508del-CFTR trafficking. To date, only few correctors that act specifically on F508del-CFTR by a direct binding and called pharmacological chaperones, are reported. These are VRT325, Corr4a, VRT532 and benzo[c]quinolizinium compounds [25 27]. Among potential proteins that bind to CFTR, we demonstrated that annexin A5 (AnxA5) binds directly to Wt- and F508del-CFTR when the channel is in the plasma membrane of cells. Indeed, we showed that AnxA5 is associated with the NBD1 of CFTR and using a sirna and overexpression approach, we showed that CFTR s channel function and membrane localization were dependent on AnxA5 expression [28]. Because our previous results showed that raised AnxA5 expression induced an augmented function of F508del-CFTR due to increased membrane localization [29], the aim of the present study was to highlight a mean to increase AnxA5 expression in F508del-CFTR expressing cells, using a drug with no side effect upon cell survival. Because it was previously showed that AnxA5 is synthesized in gonadotropes and in various cell types under the effect of gonadotropin-releasing hormone (GnRH) or some of its analogs, through mitogen activated protein kinase (MAPK), we tested the hypothesis that GnRH could increase AnxA5 expression in a human epithelial cell line and subsequently induce an increased F508del-CFTR function [30 32]. Using PCR, western blottings, iodide (I 2 ) efflux experiments and patchclamp experiments we show here that human epithelial cells express GnRH- receptors (GnRH-R) and that GnRH induces an AnxA5 overexpression and subsequently an increased Cl 2 channel function of F508del-CFTR, likely due to an increased localization of the protein in membranes. In conclusion, we show here that human epithelial cells do express GnRH-R, which has numerous physiological implications. We also propose that a topical use of GnRH is a potential treatment for CF. Methods Cell Culture The 16HBE14o 2 and the CFBE41o 2 airway epithelial cell lines were obtained from Professor D. Gruenert [33,34]. The 16HBE14o2 cell line was originally developed from human bronchial epithelium to study CFTR and retains many features of differentiated bronchial epithelial cells [33]. The CFBE41orespiratory epithelial cell line was derived from the bronchus of a patient with cystic fibrosis and immortalized with SV40. Cell culture media and supplements were purchased from Lonza (Basel, Switzerland) and PAA (Pasching, Austria). The transduced CFBE41o 2 cell lines (CFBE41o 2/ corrected called CFBE41o2/ corr and CFBE41o 2/ F508del) used in experiments were cultured as described previously [35]. CFBE41o 2/ corr and CFBE41o 2/ F508del cells were obtained by wild-type and F508del-CFTR cdna stably introduction into CFBE41o 2 cells, harbouring therefore the same genetic background [34,36]. Cells were treated with M synthetic GnRH ([Gly-OH10]-LH-RH) from Sigma-Aldrich (Saint-Louis, MO, USA) for 30, 60, 120, 180 and 310 min. Transfection For GnRH-R overexpression and inhibition, cells were transfected using Lipofectamine 2000 (Life Technologies Corporation, Carlsbad, CA, USA), according to the manufacturer s instructions, with either the human cdna clone pcmv6-xl5/gnrh-r (OriGene Technologies Inc., Rockville, MD, USA) or the human GnRH-R sirna (59-GGAAUUUGGUAUUGGUUUG-39, si- GENOME individual duplex (Thermo Fisher Scientific Inc., Waltham, MA, USA). The scrambled control sirna was sigenome Non-Targeting (Thermo Fisher Scientific Inc.). RNA Extraction and Quantitative Real-time PCR Quantitative Real-Time PCR was performed to assess the basal GnRH-R and AnxA5 mrna expression in cells and to study their modulation after GnRH treatment. Cultured medium was removed and cells were washed twice with phosphate-buffered saline (PBS). RNA was extracted using RNeasy Plus mini kit (QIAGEN, Mississauga, ON, Canada) according to the manufacturer s instructions. Extracted RNA was eluted in RNase-free water and the concentration was determinated using a nanophotometer (Implen GmbH, München, Germany). Relative quantification of the transcripts was assessed in a two steps format (RT and qpcr). Real-Time PCR was performed using a QuantiTectH SYBRH Green PCR kit (QIAGEN, Mississauga, ON, Canada), according to the manufacturer s instructions. A Chromo 4 TM System (Bio-Rad Laboratories Inc., Hercules, CA, USA) was used to amplify cdnas and detect emitted fluorescence. The following primers were used: GnRH-R (319 pb), forward 59- GACCTTGTCTGGAAAGATCC-39 and reverse 59-CAGGCT- GATCACCACCATCA-39; AnxA5 (149 pb), forward 59- TTCTCAGAGGCACTGTGACTGACT-39 and reverse 59- GATTTCCTGGCGCTGAGCATTACT-39; G3PDH (121 pb), forward 59-CCCATGTTCGTCATGGGTGTGAAC-39 and reverse 59-CAAAGTTGTCATGGATGACCTTGGC-39. Reactions were carried out with the following parameters: enzyme activation at 95uC for 15 min, denaturation at 95uC for 30 sec, annealing at 57uC for 30 sec, extension at 72uC for 30 sec and a final extension at 72uC for 10 min. 30 and 35 cycles were used for AnxA5 and GnRH-R, respectively. For negative controls (NTC), cdna was replaced by sterile RNase free water. Poly-A + mrna from human breast adenocarcinoma (MCF 7 cells) and Poly-A + mrna from human pituitary gland (both from Clontech, Member of Takara Bio Inc., Shigan, Japan) were used as positive controls. Protein Extraction and Western Blot Analysis Untreated cells and GnRH treated cells were washed twice with cold PBS and were lyzed in RIPA buffer (25 mm Tris, 150 mm NaCl, 1% Triton X-100, 1% Na-Deoxycholate, 0.1% SDS, 10 mm iodoacetamide, 100 mm PMSF; ph = 7.5) in the presence of Complete Protease Inhibitor Cocktail (Roche, Basel, Switzerland). Total protein concentrations were determined using Lowry s methodology using bovine serum albumin as a standard [37]. Samples were resolved by SDS-PAGE (7.5 to 10%) and transferred onto a PVDF membrane (GE Healthcare, Chalfont St Giles, UK). After blotting, membranes were blocked with 5% (w/v) non-fat dried skimmed milk in PBS-0.1%Tween 20 or TBST (Tris-buffered saline plus 0.1% Tween 20). Blots were probed overnight at 4uC with mouse monoclonal antibody anti- GnRH-R (1:150, LH-RH Receptor Ab-3, clone GNRH03, MM France, Francheville, France) or goat polyclonal antibody anti- AnxA5 (1:400, clone R-20, Santa Cruz Biotechnology, CA, USA). Blots were further incubated with HRP-conjugated secondary antibodies [1:20000, anti-mouse and anti-goat secondary antibody were from Santa Cruz Biotechnology and Abcam (Cambridge, UK), respectively] and visualized by enhanced chemiluminescence. ECL Plus and Forte chemiluminescence detection kits were from GE Healthcare and Merk Millipore (Billerica, MA, USA), PLOS ONE 2 February 2014 Volume 9 Issue 2 e
 after separation (2% agarose gel).")
131 GnRH Improves Chloride Transport in CF Cells Figure 1. Basal mrna and protein expression of GnRH-R. A. The upper left panel shows representative PCR bands for GnRH-R (319 bp) after separation (2% agarose gel). A single band is observed in 16HBE14o 2 (lane 1), CFBE41o 2 (lane 2), CFBE41o 2/ corr (lane 3) and CFBE41o 2/ F508del (lane 4) cells. No signal is observed in the negative control (lane 5). The upper right panel shows positive controls (human pituitary gland and human MCF7cell line, lanes 1 and 2, respectively) for the detection of GnRH-R. cdnas were from the human pituitary gland and the human MCF7cell line (breast adenocarcinoma), respectively. The mrna expression of G3PDH (121 bp) is shown lower panel for the same samples. MW is the molecular weight given in base pairs (bp). B. The quantitative analysis (n = 5) of the mrna of GnRH-R. Ct is the Cycle Threshold. A hight Ct correspond to low mrna abundance because more PCR cycles are needed to detect it. Endogenous control has a lower Ct than the target mrna. Therefore, a low DCt ( = Ct gene Ct control) correspond to high mrna abundance C. Upper panel shows representative immunoblots of GnRH-R (64 kda) and lower panel shows representative immunoblots of G3PDH (37 kda). A protein expression is observed in 16HBE14o 2 (lane 1), CFBE41o 2 (lane 2), CFBE41o 2/ corr (lane 3) and CFBE41o 2/ F508del (lane 4) cells. A positive control (lane 5) was used for the detection of GnRH-R (human breast duct carcinoma (T-47D) whole cell lysate). G3PDH was used for further normalization. D. The densitometric analysis of GnRH-R expression (n = 6) did not show any difference between cell lines. doi: /journal.pone g001 respectively. The relative intensity of each band was estimated by densitometry using BIO-1D software (BioRad). Each value was normalized to the amount of G3PDH in the same lane which was assessed on the same immunoblots probed with mouse monoclonal antibody anti-g3pdh (1:30000, clone 6C5, Interchim SA, Montluçon, France). Positive controls for GnRH-R and AnxA5 were a whole cell lysates of human breast duct carcinoma (T-47D, Tebu-Bio, Le-Perray-en-Yvelines, France) and pure AnxA5 from human placenta (33 kd, Sigma-Aldrich), respectively. PLOS ONE 3 February 2014 Volume 9 Issue 2 e
132 GnRH Improves Chloride Transport in CF Cells Figure 2. Validation of the detection of GnRH-R in immunoblots. A. Representative immunoblots of GnRH-R detection (upper panel) after 48 h transfection with the Human cdna clone pcmv6-xl5/gnrh-r in 16HBE14o 2 (1) and CFBE41o 2 (2) cells. pcmv6-xl5 empty plasmid was used as a control. B. The densitometric analysis after normalization by G3PDH expression and comparison with the controls, indicate that the GnRH-R expression in significantly increased, (n = 5). C. Representative immunoblots of GnRH-R detection after 72 h transfection with a sigenome individual duplex targeting GnRH-R in 16HBE14o 2 (1) and CFBE41o 2 (2) cells. sigenome Non-Targeting was used as control. A decreased expression of GnRH-R is observed in both cell types. D. The densitometric analysis after normalization by G3PDH expression and comparison with the controls, indicate that the GnRH-R expression is significantly decreased in 16HBE14o 2 (1) and CFBE41o 2 (2) cells (n = 7) in the presence of sirna. doi: /journal.pone g002 PLOS ONE 4 February 2014 Volume 9 Issue 2 e

133 GnRH Improves Chloride Transport in CF Cells to establish the baseline (k peak-k basal, min 1 ). Activators were included in the efflux buffer from time 3 min, and collection continued at 1-min intervals for an additional 7 min in the continued presence of tested compounds (for other details, [38])). To test the effect of GnRH exposure on halide permeability, CF and non-cf cells monolayers were incubated with 0.1 to 100 nm GnRH for 1 h before the efflux experiment. Figure 3. Basal mrna and protein expression of AnxA5. A. The left image shows representative PCR bands for AnxA5 (149 bp) and the right image shows representative PCR bands for G3PDH (121 bp, 2% agarose gel). A single band is observed in 16HBE14o 2 (lane 1), CFBE41o 2 (lane 2), CFBE41o 2/ corr (lane 3) and CFBE41o 2/ F508del (lane 4) cells. No signal is observed in the negative control (lane 5). MW is the molecular weight given in base pairs (bp). B. The gene expression of AnxA5 was determined by quantitative Real-Time PCR in each cell line and data were analyzed by using DCt (target reference). Comparison between CFBE41o 2/ corr (3) and CFBE41o 2/ F508del (4) cells show a higher expression in CF cells. C. The upper image shows a representative immunodetection of AnxA5 protein (35 kda) and the lower image shows the immunodetection of G3PDH (37 kda) protein expression in 16HBE14o 2 (lane 1), CFBE41o 2 (lane 2), CFBE41o 2/ corr (lane 3) and CFBE41o 2/ F508del (lane 4) cells. Pure AnxA5 was used as a positive control (lane 5). G3PDH was detected to show that the loading was identical in each lane and for further normalization. D. Statistical analysis of AnxA5 expression (n = 6) indicate that the expression of AnxA5 does not vary among cell lines although it tends to be higher in CFBE41o 2/ corr and CFBE41o 2/ F508del cells than in 16HBE14o 2 or CFBE41o 2 cells. doi: /journal.pone g003 I 2 Effluxes Experiments Cells were cultured in 24-well microplates for 3 to 7days. Confluent monolayers were used, and iodide effluxes were assayed as previously described [38]. Briefly, the F508del-CFTR Cl channel activity was assayed by measuring the rate of iodide ( 125 I) efflux from cells and time-dependent rates of 125 I efflux were calculated from the following: ln ( 125 I t1/ 125 It2)/(t1 t2), where 125 It is the intracellular 125 I at time t, and t1 and t2 successive time points. Curves were constructed by plotting rates of 125 I versus time. All comparisons were based on maximal values for the timedependent rates (k = peak rates, min 1 ), excluding the points used Patch-clamp Experiments Patch-clamp experiments were performed with an automatic Nanion s electrophysiology workstations Port-a-Patch (Nanion Technologies GmbH, Germany) coupled to an external amplifier unit HEKA EPC-10 [39]. Whole-cell patch-clamp recordings were performed at room temperature (20 25uC) on isolated CFBE41o2/corr and CFBE41o2/F508del cells, with and without GnRH treatment. Before recording, the culture medium was replaced by the external solution contained the following (in mm): 140 NaCl, 2 CaCl2, 1 MgCl2, 10 Hepes, 5 D-glucose monohydrate. Its final ph value was adjusted to 7.4 with NaOH and its osmolarity was 298 mosmol. The internal solution contained the following (in mm): 50 CsCl, 10 NaCl, 60 Cs- Fluoride, 20 EGTA, 10 Hepes/CsOH, and 5 Mg-ATP (Mg salt); ph 7.2; 285 mosmol. CFTR-inhibitor solution (10 mm CFTRinh172, Sigma) and CFTR-activator solution (10 mm forskolin and 30 mm genistein both from Sigma) were added to the external solution to inhibit or activate CFTR, respectively. Voltage-clamp pulses were generated and data were captured using the Patchmaster program (Nanion Technologies GmbH, Germany). Voltage-dependent properties were assessed by applying voltage steps of 100 ms duration from a holding potential of - 80 mv to test potentials ranging from 280 to +80 mv with 10 mv increments. Series resistance was compensated. For graphic representations of I/V relationship currents were normalized by membrane capacitances to remove variability due to differences in cell sizes. Cell Surface Biotinylation Cells were grown in serum-free medium overnight before GnRH incubation. Culture medium was then removed and cells were washed three times with PBS plus 0.1 mm CaCl2 and 1 mm MgCl2 (ph 7.4) and once with PBS (ph 8). Cells were exposed to sulfo-nhs-ss-biotin for 30 min on ice, rinsed twice with BSA quenching buffer (1% BSA in PBS, ph 7.4) and incubated 10 min in the same buffer. Cells were scraped in PBS, ph 7.4 and centrifuged at 2400 rpm (6 min, 4uC). Cell pellets were suspended in RIPA buffer plus anti-proteases and incubated for 30 min on ice. The resulting lysates were centrifuged at g for 15 min at 4uC and total cellular protein content was determined using Lowry s method. The supernatants were incubated with streptavidin beads (Novagen/EMD Chemicals, Madison, WI, USA) overnight at 4uC. After a brief centrifugation, supernatants were removed and the beads were washed four times in RIPA. Biotinylated proteins were eluted from streptavidin beads using 5X sample buffer containing 2-mercaptoethanol to cleave the NHS-SS-biotin and western blots were probed overnight at 4uC with a mouse monoclonal anti-cftr antibody (1:500, clone M3A7, Merk Millipore). For normalization, Na + /K + -ATPase expression was assessed on the same blots probed with a mouse monoclonal antibody (1:200, clone 9-A5, Santa Cruz Biotechnology). A positive control was used to assess the mature band of CFTR (T84, human colorectal carcinoma whole cell lysate). PLOS ONE 5 February 2014 Volume 9 Issue 2 e

134 GnRH Improves Chloride Transport in CF Cells Figure 4. AnxA5 expression when cells are treated for 60 min with GnRH. A. Representative immunoblots showing AnxA5 and G3PDH expression in 16HBE14o 2 (1), CFBE41o 2 (2), CFBE41o 2/ corr (3) and CFBE41o 2/ F508del (4) cells after 60 min of treatment by GnRH (10 29 M). An increased expression is observed in all cell types. B. Densitometric analysis of AnxA5 expression. Data are normalized by G3PDH. AnxA5 protein expression is presented as mean 6 S.E.M (7 independent experiments with n $2 for each experiment). For each cell line, the statistical analysis was performed by comparing the normalized amount of AnxA5 between cells incubated with buffer alone (white bars adjusted to 1) and cells incubated with GnRH (black bars). doi: /journal.pone g004 Confocal Microscopy CFBE41o 2/ corr and CFBE41o 2/ F508del cells were plated on 18-mm diameter glass coverslips at low density and maintained in culture at 37uC for 24 to 48 h before exposure to GnRH. Cells were rinsed in TBS, fixed (3% paraformaldehyde in TBS, 20 min, room temperature), rinsed in TBS, permeabilized (0.10% Triton X-100 in TBS) and rinsed again. The samples were then stained by indirect immunofluorescence. After fixation, non-specific sites were blocked with 0.5% bovine serum albumin (BSA) in TBS (60 min at room temperature). The cells were then stained with primary antibody (anti-cftr antibody 24-1, 1:50, RD Systems, Minneapolis, MN, USA) for 90 min at room temperature and rinsed four times in TBS containing 0.5% BSA. Next, samples were incubated with the goat anti-mouse secondary antibody conjugated to Cy3 (1:400, Jackson Immunoresearch Laboratories, West Grove, PA, USA) for 60 min at room temperature. After final washes, samples were mounted in VectaShield plus DAPI (VECTOR Laboratories Inc., Burlingame, CA, USA), dried, and viewed with a confocal laser-scanning microscope (LSM 780, Axio Observer, Plan-Apochromat 63X/1.40 oil; Carl Zeiss GmbH, Jena, Germany). Negative controls in which primary antibody was omitted was performed. Statistics Results are expressed as the means 6 S.E.M of n observations. Data were compared using the Student s t test analysis with STATGRAPHICS version 4.1 (SIGMA PLUS, Levallois-Perret, France) and differences were considered statistically significant when p,0.001 (***); p,0.01 (**) and p,0.05 (*). Results Basal Expression of GnRH-R Basal mrna and protein expression of GnRH-R was assessed in 16HBE14o 2, CFBE41o 2, CFBE41o 2/ corr and CFBE41o 2/ F508del cells. As shown in Fig. 1A, a single PCR band for GnRH-R (319 bp) was observed in the four cell lines whereas no signal was observed in the negative control. cdnas from human pituitary gland and human MCF7 cell line (breast adenocarcinoma), were used as positive controls. The mrna expression of G3PDH (121 bp) in the same samples showed no variation of the signal intensity. The mrna expression of GnRH- R was quantified (Fig. 1B). Because the genetic background is only comparable between CFBE41o2/corr and CFBE41o2/F508del cells, statistical analysis was only performed for these cells and a higher mrna expression was observed in CFBE41o2/F508del cells. Western blots were performed to detect GnRH-R (64 kda) and G3PDH (37 kda) in the cell lines (Fig. 1C), using a whole cell lysate of human breast duct carcinoma (T -47D) as a control. The results indicate that the GnRH-R is present in all the samples. The densitometric analysis of the signals (Fig. 1D) did not show any difference between CFBE41o2/corr and CFBE41o2/F508del cells. To assess the GnRH-R antibody s specificity, the GnRH-R s expression was modulated before immunodetection. First, PLOS ONE 6 February 2014 Volume 9 Issue 2 e
135 GnRH Improves Chloride Transport in CF Cells expression in the cells expressing the mutated CFTR. AnxA5 expression was further assessed by western blottings (Fig. 3C). The statistical analysis (Fig. 3D) indicated that AnxA5 expression does not vary within cell lines. Nevertheless, the amount of AnxA5 protein tended to be higher in CFBE41o2/corr and CFBE41o 2/ F508del cells than in 16HBE14o 2 or CFBE41o 2 cells. AnxA5 Expression in Cells Submitted to GnRH Treatment for 60 min In order to determine whether GnRH modulates AnxA5 expression and to determine the shortest time to be used for GnRH treatment, 16HBE14o 2, CFBE41o 2, CFBE41o 2/ corr and CFBE41o 2/ F508del cells were treated with M GnRH for 30, 60, 120, 180 and 310 min (not shown). At each time point, the protein expression of AnxA5 was evaluated and normalized with that of G3PDH. The AnxA5 expression was compared to that of cells incubated with buffer alone used as a control. Fig. 4A shows an example of the obtained western blots at the 60 min time point. An increased intensity of the AnxA5 band was observed in all cell types. Densitometry of the bands, normalization using G3PDH and quantification were performed (Fig. 4B). It was concluded that a 60 min treatment led to a significant increase of AnxA5 expression in the four cell types. Figure 5. Activation of wild-type and F508del-CFTR chloride channel activity in cells treated by GnRH. A. Example of mean traces showing the restoration of an iodide efflux in 16HBE14o- cells untreated or treated 1 h with 1 nm GnRH and stimulated by 10 mm forskolin and 30 mm genistein. B. Bar graph showing the enhancement of the chloride channel function of CFTR in 16HBE14o 2, CFBE41o 2, CFBE41o 2/ corr and CFBE41o 2/ F508del cells (1, 2, 3 and 4, respectively) using iodide efflux experiments. Histograms show the mean relative rate of iodide efflux in each cell type, with and without GnRH treatement. A significant increase is observed in the presence of GnRH (n = 4). doi: /journal.pone g005 16HBE14o 2, CFBE41o 2 cells were transfected with a GnRH-R expression vector in which the cdna encoding GnRH-R was ligated. As shown in Fig. 2A, the immunoblots indicated that the transfection induced an increased expression of the receptor. The control cells were transfected with the empty vector and G3PDH was detected for further normalization. The quantitative analysis of the blots (Fig. 2B) showed that the transfection of the expression vector induced an increased expression of GnRH-R of 70% and 44% in 16HBE14o 2, CFBE41o 2 cells, respectively, when compared to the control cells. Second, a sirna directed against GnRH-R was used and a decreased expression of the protein was observed in both cell types (Fig. 2C). The quantitative analysis of the blots (Fig. 2D) showed that the sirna induced an decrease expression of GnRH-R of 64% and 65% in 16HBE14o 2, CFBE41o 2 cells, respectively, when compared to the control cells which were transfected with a scrambled sirna. Basal Expression of AnxA5 Basal mrna and protein expression of AnxA5 was assessed in 16HBE14o 2, CFBE41o 2, CFBE41o 2/ corr and CFBE41o2/ F508del cells. As shown in Fig. 3A, AnxA5 mrna (149 bp) was detected in all cell lines whereas, no signal was observed in the negative control. The gene expression of AnxA5 was quantified in cells using Real-Time PCR. Analysis of the data (Fig. 3B) between CFBE41o2/corr and CFBE41o2/F508del cells showed a higher Increased Endogenous F508del-CFTR s Function by GnRH Treatment To test the activity of CFTR channels under GnRH treatment in 16HBE14o 2, CFBE41o 2, CFBE41o 2/ corr and CFBE41o _ / F508del cells, functional analysis was performed by measuring the camp regulated and CFTR-dependent iodide efflux after 1 h incubation at 37uC with GnRH (10 29 M). The activity of CFTR was stimulated by a mixture of adenylate cyclase activator forskolin (Fsk, 10 mm) plus isoflavone genistein (Gst, 30 mm) or inhibited by the thiazolidinone specific CFTR blocker CFTRinh- 172 (10 mm). Example of mean traces showing an increased iodide efflux in 16HBE14o 2 cells, untreated or treated with GnRH is shown in Fig. 5A. This example shows that after adding Fsk/Gst, the rate of iodide efflux increased at a maximum rate (peak value) after 4 min and that the peak was higher in the presence of GnRH. Experiments were conducted for all cell types and the results are represented as bar graphs in Fig. 5B. Statistical analysis indicated that in the presence of GnRH, the rates of iodide efflux were significantly increased in 16HBE14o 2, CFBE41o 2, CFBE41o 2/ corr and CFBE41o 2/ F508del cells when compared to untreated cells, indicating that CFTR and F508del-CFTR s function are increased by GnRH. The concentration-response effect of GnRH was further determined in the presence or absence of Fsk and Gst (Fig. 6). Calculated an EC50 in 16HBE14o 2, CFBE41o 2, CFBE41o 2/ corr and CFBE41o 2/ F508del cells were nm, nm, nm and nm, respectively. CFTR s function in CFBE41o2/corr and CFBE41o_/F508del cells was also assessed by patch-clamp experiments in the whole cell configuration ((n = 5 to 9 cells from different cell cultures, Fig. 7). Mean cell capacitances were 21.4 and pf for CFBE41o2/corr and CFBE41o_/F508del cells, respectively. Fig. 7A (upper panel) show the normalized I/V curve obtained when CFBE41o2/corr cells were treated or not with GnRH for 1 hour, with activators and with the CFTR inhibitor. It can be observed that activation and inhibition were efficient in both cases showing the specificity of the recordings. It was also observed that GnRH treatment of the cells induced an increased current density. PLOS ONE 7 February 2014 Volume 9 Issue 2 e
136 GnRH Improves Chloride Transport in CF Cells Figure 6. Concentration-dependent effect of GnRH. Concentration-dependent enhancement of CFTR s function by GnRH in 16HBE14o 2 (A), CFBE41o 2 (B), CFBE41o 2/ corr (C) and CFBE41o 2/ F508del (D): CFTR activity was assessed with the iodide effluxes technique in the presence of forskoline (10 mm) plus genistein (30 mm) after 1h of incubation. Results are expressed as normalized means (6 SEM, n = 4). doi: /journal.pone g006 Statistical analysis was further performed and presented as a bar graph (Fig. 7A, lower panel). In untreated cells inhibition and activation of CFTR were significantly efficient whereas in treated cells only inhibition led to a significant decreased current density. When untreated cells were compared to treated cells, solely the basal current densities were significantly increased by GnRH. The absence of activation by Fsk/Gst in GnRH treated cells could be explained by the fact that the concentration of the activators was the same in both treated and untreated cells and because the basal current density was higher in the presence of GnRH. The drawn conclusion is that GnRH induces an increased current density in CFBE41o2/corr cells. I/V curves obtained with CFBE41o_/ F508del cells in the same conditions as for CFBE41o2/corr cells are shown in Fig. 7B (upper panel). As in Fig. 7A, basal normalized currents, as well as modulated currents by activation and inhibition were higher in the presence of GnRH. Statistical analysis and bar graphs (Fig. 7B, lower panel) showed a basal current in untreated CFBE41o_/F508del cells, likely due to the presence of Mg-ATP in the internal solution, activating the weak quantity of CFTR within membranes. The use of Fsk/Gst and CFTRinh-172 was inefficient upon the mutated CFTR s function. In treated cells, the basal current density was close to 5 times higher than in untreated cells. This difference was highly significant despite being lower than in CFBE41o2/corr cells. In these conditions, Fsk/Gst and CFTRinh-172 were efficient as in corrected cells. Therefore, F508del-CFTR s function under GnRH treatment exhibited the same pattern as Wt-CFTR, with a much lower intensity (in the range of 100 pa). Increased Membrane Expression of CFTR after GnRH Treatment The presence of CFTR within membranes after 1 hour treatment by GnRH was assessed by biotinylation experiments. Cells were biotinylated with Sulfo-NHS-SS-biotin, pulled-down with streptavidin-agarose and subjected to 7.5% SDS-PAGE. Total cell lysates were used as controls. Representative immunoblots showing the detection of CFTR in biotinylated surface proteins from 16HBE14o 2 and CFBE41o 2/ corr cells are shown in Fig. 8A and 8B, respectively. CFTR was present in total lysates and in pulled down samples. The densitometric analysis of CFTR s cell surface expression was performed and normalized with the biotinylated Na + /K + -ATPase level. The statistical analysis clearly indicates that the amount of biotinylated CFTR is higher after GnRH treated cells (Fig. 8C). An increased amount of CFTR in membranes of 33 and 36% was observed in 16HBE14o 2 and CFBE41o 2/ corr cells, respectively. Whereas CFTR was detected in controls, we failed to detect it in biotinylated surface proteins from CFBE41o 2 and CFBE41o 2/ F508del cells (not shown). PLOS ONE 8 February 2014 Volume 9 Issue 2 e
137 GnRH Improves Chloride Transport in CF Cells Increased Membrane Localization of CFTR after GnRH Treatment Because we failed to detect CFTR in CFBE41o 2 and CFBE41o 2/ F508del cells due to the poor expression of the protein, membrane localization of CFTR in CFBE41o 2/ F508del cells was assessed by confocal microscopy, in the absence (Fig. 9A) and in the presence (Fig. 9B) of a GnRH treatment (120 min). Without any treatment, CFTR was mainly observed around the nuclei of the cells. We therefore speculated that it was present in the ER. When cell were treated with GnRH, CFTR had a more spread localization and it was observed at the periphery of the cells, looking like the plasma membrane, as shown in Fig. 9B by white arrows. Controls performed without CFTR antibody showed no labelling (not shown). Comparison of the Effect of Miglustat and GnRH The time-dependent effects of miglustat and GnRH were assessed and compared in CFBE41o 2 and CFBE41o 2/ F508del cells (Fig. 10). CFTR s function was assessed by the iodide effluxes technique in the presence of 1 nm GnRH or 100 mm miglustat. As shown in Fig. 10A, miglustat induced an increased iodide efflux in CFBE41o 2 cells after 1 hour treatment. The increase was higher at the 2 hours time point. GnRH also induced an increased iodide efflux but the effect was lower, but significant, and did not increase after 2 hours of treatment. In CFBE41o 2/ F508del cells (Fig. 10B), the same time-dependent effect as in CFBE41o 2 cells was observed. Figure 7. Activation of CFTR current in CFBE41o2/corr and CFBE41o2/F508del cells by patch clamp experiments. A. Upper panel shows I/V curves (normalized by cell capacitance pf, mean6sem) for CFTR current in CFBE41o2/corr cells in the whole cell configuration. Basal condition is with MgATP in the intracellular buffer. CFTR was activated by forskolin plus genistein and inhibited by CFTR inhibitor 172. Curves obtained with and without GnRH treatment (1 hour) were plotted. The lower panel shows the statistical analysis of the currents noted at +80 mv. B. Same as in A for CFBE41o2/F508del cells. doi: /journal.pone g007 Discussion The most common mutation causing CF, F508del-CFTR, is known for more than 20 years [1]. The F508del-CFTR nascent protein is recognized by the cellular quality control machinery and is degraded. A very small amount succeeds in trafficking to the cell membrane where it functions as a Cl 2 channel. Many efforts were performed in order to identify molecules that could correct the trafficking defect. Indeed, it is estimated that the required correction to ameliorate the symptoms in CF patients is between 10% and 30% of wild-type CFTR function [40,41]. Among proteins that directly bind to CFTR, we previously showed that AnxA5 binds to NBD1 and, when over expressed, may restore some Cl 2 channel function due to an increased stabilization of CFTR in membranes [28,29]. AnxA5 belongs to the annexin family which is characterized by calcium-dependent phospholipid binding. Annexins are found in a wide variety of species, from plants to human [42 44]. They are known to inhibit PLA2, PKC, and blood coagulation [42]. Under specific conditions, AnxA5 can also form a calcium channel [45]. Despite these molecular characteristics, their physiological role within cells remains unclear. Whereas the CFTR/AnxA5 interaction as well as the augmented Wt-CFTR whole-cell currents, due to over-expression of AnxA5, was further confirmed, it was not seen in F508del- CFTR expressing cells [46]. The discrepancy with our previous results and within the referred study is likely due to the used cell system. These authors suspect that AnxA5 does not interact specifically with CFTR. This discrepancy is likely due to the use of a non specific AnxA5 antibody [46]. Because it is suggested that some CFTR is expressed at the apical membrane in epithelial cells from CF patients [47], despite a reduced level when compared to non-cf cells, and because we found a positive effect of AnxA5, our aim was to highlight a non toxic mean to increase AnxA5 expression in F508del-CFTR expressing cells. Some previous results clearly demonstrate that GnRH stimulation induces the PLOS ONE 9 February 2014 Volume 9 Issue 2 e
138 GnRH Improves Chloride Transport in CF Cells Figure 8. Membrane expression of CFTR after GnRH treatment. The presence of CFTR within membranes was detected by biotinylation experiments. A. Representative immunoblots showing the detection of CFTR in biotinylated surface proteins from 16HBE14o 2 cells. CFTR is present in total proteins (left) and in biotinylated surface proteins (right) after GnRH treatment (10 29 M) for 60 min. The arrowheads indicate the fully glycosylated (180 kda) CFTR. B. Same image as in A. with CFBE41o 2/ corr cells. C. Histograms of the densitometric analysis of CFTR s cell surface expression (n = 4) in 16HBE14o 2 (1) and CFBE41o 2/ corr (2) cells The biotinylated CFTR level is normalized to the biotinylated Na + /K + -ATPase level. Cells incubated with buffer alone are referred as 1 arbitrary unit. doi: /journal.pone g008 expression of AnxA5 in the gonadotropes, luteal cells and other cell types [30 32]. Therefore, we tested the hypothesis of an AnxA5 increased synthesis in response to GnRH stimulation in human epithelial cells of the airways. GnRH is secreted by GnRH neurons in the hypothalamus and is transported to the gonadotropes of the anterior pituitary gland. It is a decapeptide involved in the control of reproduction and binds to a specific G protein-coupled receptor to allow gonadotropin secretion. GnRH and its receptor are reported to be expressed in human tissues outside of the hypothalamus, such as liver, heart, skeletal muscle and kidney where its functions are unclear [48]. The first step of the present work was to assess the expression of GnRH-R and AnxA5 in human epithelial cells (Fig. 1 to 3). PLOS ONE 10 February 2014 Volume 9 Issue 2 e
 and in CFBE41o 2/ F508del cells (B).")
139 GnRH Improves Chloride Transport in CF Cells Figure 10. Comparison of the effect of miglustat and GnRH. Comparison of the time-dependent effect of GnRH or miglustat upon F508del-CFTR function in CFBE41o- cells (A) and in CFBE41o 2/ F508del cells (B). CFTR activity was assessed with the iodide effluxes technique in the presence of forskolin (10 mm) plus genistein (30 mm) after 1 nm GnRH or 100 mm miglustat incubation (n = 4). doi: /journal.pone g010 Figure 9. Immunolocalization of CFTR in CFBE41o2/F508del cells after GnRH treatment. A. Representative confocal photomicrographs of the localization of F508del-CFTR in cells without any treatment. CFTR is likely observed around the nuclei, in the endpoplasmic reticulum. B. Representative confocal photomicrographs of the localization of F508del-CFTR in cells after GnRH treatment. CFTR is likely observed in the endpoplasmic reticulum but also in membranes. doi: /journal.pone g009 16HBE14 o 2, CFBE41o 2 and the transduced CFBE41o 2 airway epithelial cell lines CFBE41o 2/ corr and CFBE41o 2/ F508del were used [35]. Using PCR and western blottings, we observed an AnxA5 and GnRH-R expression in these cells. Because the genetic background of 16HBE14o 2, CFBE41o 2 cells are different, their GnRH-R expression was not compared. The GnRH-R expression (protein) did not vary between CFBE41o 2/ corr and CFBE41o 2/ F508del cells. Because anti-gnrh-r antibodies are debated, we decided to assure us of the results. For this, GnRH-R expression was modulated (increased and decreased) before immunodetection and we showed that the observed band was indeed GnRH-R. The GnRH-R expression in airway epithelial cells is in accordance with previous results showing its expression in extrapituitary tissues. A high expression level of the mrna was found in prostate, thymus, and kidney. In heart, brain, placenta, lung, liver, skeletal muscle, pancreas, colon, ovary, small intestine, spleen, and testis the mrna was also detected [49]. The presence and localization of GnRH-R in airway epithelial cells (Calu-3) was also shown using immunochemical methods [49,50]. Here, we present for the first time the GnRH-R expression in 16HBE14o 2, CFBE41o 2 and the transduced CFBE41o 2 airway epithelial cell lines, using immunoblots. AnxA5 expression was studied in our cell models at the mrna and protein level and we found that it is also expressed. This result is in accordance with previous results [51]. Therefore, GnRH-R and AnxA5 expression being shown, further experiments were conducted. The second step of the work was to study AnxA5 expression under GnRH treatment and the effect of the hormone on the function of CFTR (Fig. 4 to 6). The four cell types were treated with GnRH (10 29 M) for 30, 60, 120, 180 and 310 min (not shown) and western-blottings were performed. A significant increased AnxA5 expression was observed at the 60 min time point and remained stable until 310 min. This increased expression of AnxA5 under GnRH treatment was previously PLOS ONE 11 February 2014 Volume 9 Issue 2 e
.")
140 GnRH Improves Chloride Transport in CF Cells Figure 11. Schematic representation of the GnRH s effect on F508del-CFTR function. We found that F508del-CFTR s membrane localization and function are increased under GnRH treatment, associated with an increased expression of AnxA5 (red lines). Nevertheless, according to previous results, AnxA5 over expression is likely not the solely event triggered in cells by GnRH. Described events are summarized in black lines and their possible involvement upon F508del-CFTR activity are shown in dotted black lines. doi: /journal.pone g011 described. Indeed, a continuous exposure to GnRH driving a specific gene expression of AnxA5 without desensitization was reported [32]. Nevertheless, our further experiments were conducted at 60 min of GnRH treatment, corresponding to a significant increased AnxA5 expression in 16HBE14o 2, CFBE41o 2, CFBE41o 2/ corr and CFBE41o 2/ F508del cells. Functional analysis was performed by measuring the CFTRdependent iodide efflux in the absence and in the presence of GnRH. Statistical analysis indicated that with GnRH, the rates of iodide efflux were significantly increased in 16HBE14o 2, CFBE41o 2, CFBE41o 2/ corr and CFBE41o 2/ F508del cells when compared to untreated cells. EC 50 were relatively low (2.2 nm in CFBE41o 2/ F508del cells). To provide further evidence of an increased channel function of CFTR in CFBE41o2/F508del cells, whole-cell patch-clamp experiments were performed with and without GnRH treatment of the cells. The main conclusion drawn from these experiments is that in CFBE41o2/corr as well as in CFBE41o2/F508del cells, current densities via CFTR are increased in the presence of GnRH, corroborating the results obtained in iodide efflux experiments. In CFBE41o2/F508del cells, GnRH also lead to a regulation of CFTR by Fsk/Gst and CFTRinh-172, what was not observed in untreated cells. Therefore, GnRH increases sufficiently the chloride channel function of the mutated CFTR to lead to the observation of its modulation. Our third step was to give some potential explanations to the observed increased Cl 2 channel function of CFTR in the presence of GnRH. We assessed a possible increased anchoring of CFTR within membranes by biotinylation and confocal microscopy (Fig. 8, 9). According to our previous work [28,29], we found that the presence of F508del-CFTR was increased in membranes. Our results showing in the CFBE41o2/F508del cells an increased CFTR membrane localization after GnRH treatment could not be related to the additional plasmid inserted in the CFBE41o- cells because in biotinylation experiments without GnRH, the amount of CFTR in 16HBE14o- cells is not different than in CFBE41o2/ corr cells. Nevertheless, the observed increased Cl 2 channel function of CFTR in the presence of GnRH is likely not only due to the weak AnxA5 overexpression. The presence of CFTR within membranes after treatment was difficult to detect and due to the pleiotropic effects of GnRH, we hypothesize that the increased AnxA5 expression in likely not the only reason for this effect. Indeed, the expression of AnxA5 is directly stimulated by GnRH, probably through protein kinase C, MAKP and calcium pathways which are also involved in the regulation and in the targeting of CFTR in membranes [30 32,38,52 54]. A schematic representation of our findings together with known effects of GnRH is presented in Fig. 11. The involved pathways explaining the increased membrane localization of F508del-CFTR when cells are stimulated by GnRH, as well as the involvement of calcium are PLOS ONE 12 February 2014 Volume 9 Issue 2 e
141 GnRH Improves Chloride Transport in CF Cells under investigation in our laboratory. Interestingly, the rapid and sustained action of GnRH seems different of the action of correctors or potentiators. We found of interest to compare the action of the well known corrector miglustat [24] with that of GnRH (Fig. 10). Time dependant effects of miglustat and GnRH were different. Whereas, miglustat induced a continuous increased iodide efflux until 2 hours, the GnRH effect observed at the 1 hour time point remained stable until the 2 hours. This observation permit to say that actions are likely different and reinforce the hypothesis that GnRH does not act as a corrector. Miglustat permits F508del-CFTR to escape the ER whereas GnRH maintains it in membranes. The association of both miglustat and GnRH will be further explored in primary cultures. Nevertheless, beside miglustat, several classes of F508del-CFTR correctors have been identified and could be tested. Indeed, despite many efforts to identify potentiators (such as VX-770) and correctors (such as VX-809), and, recently, a class of compounds with dual potentiator and corrector activities, much work remain to find out new molecules with beneficial effects on F508del- CFTR defects (for review [55]). The study of a combined effect of GnRH together with recently described compounds would be of interest to assess the benefit of the association of such molecules. References 1. Riordan JR, Rommens JM, Kerem B, Alon N, Rozmahel R et al. (1989) Identification of the cystic fibrosis gene: cloning and characterization of complementary DNA. Science 245: Bear CE, Li CH, Kartner N, Bridges RJ, Jensen TJ et al. (1992) Purification and functional reconstitution of the cystic fibrosis transmembrane conductance regulator (CFTR). Cell: 68: Amaral MD (2004) Cftr and chaperones: Processing and degradation. J MolNeurosci 23: Cheng SH, Gregory RJ, Marshall J, Paul S, Souza DW et al. (1990) Defective intracellular transport and processing of cftr is the molecular basis of most cystic fibrosis. Cell 63: Lukacs GL, Mohamed A, Kartner N, Chang XB, Riordan JR et al. (1994) Conformational maturation of cftr but not its mutant counterpart (delta f508) occurs in the endoplasmic reticulum and requires atp. EMBO J 13: Jensen TJ, Loo MA, Pind S, Williams DB, Goldberg AL et al. (1995) Multiple proteolytic systems, including the proteasome, contribute to cftr processing. Cell 83: Ward CL, Omura S, Kopito RR (1995) Degradation of cftr by the ubiquitinproteasome pathway. Cell 83: Hwang TC, Sheppard DN (2009) Gating of the CFTR Cl 2 channel by ATPdriven nucleotide-binding domain dimerisation. J Physiol 587: Zielenski J, Tsui LC (1995) Cystic fibrosis: genotypic and phenotypic variations. Annu Rev Genet 29: Cheng SH, Gregory RJ, Marshall J, Paul S, Souza DW et al. (1990) Defective intracellular transport and processing of CFTR is the molecular basis of most cystic fibrosis. Cell 63: Denning GM, Anderson MP, Amara JF, Marshall J, Smith AE et al. (1992) Processing of mutant cystic fibrosis transmembrane conductance regulator is temperature-sensitive. Nature 358: Sato S, Ward CL, Krouse ME, Wine JJ, Kopito RR (1996) Glycerol reverses the misfolding phenotype of the most common cystic fibrosis mutation. J Biol Chem 271: Welsh MJ, Denning GM, Ostedgaard LS, Anderson MP (1993) Dysfunction of CFTR bearing the delta F508 mutation. J Cell Sci. Suppl. 17: Dalemans W, Barbry P, Champigny G, Jallat S, Dott K et al. (1991) Altered chloride ion channel kinetics associated with the DF508 cystic fibrosis mutation. Nature 354: Ostedgaard LS, Rogers CS, Dong Q, Randak CO, Vermeer DW et al. (2007) Processing and function of CFTR-DF508 are species-dependent. Proc Natl Acad Sci USA 104: Miki H, Zhou Z, Li M, Hwang TC, Bompadre SG (2010) Potentiation of disease-associated cystic fibrosis transmembrane conductance regulator mutants by hydrolyzable ATP analogs. J Biol Chem 285: Lukacs GL, Chang XB, Bear C, Kartner N, Mohamed A et al. (1993) The delta F508 mutation decreases the stability of cystic fibrosis transmembrane conductance regulator in the plasma membrane. Determination of functional half-lives on transfected cells. J. Biol. Chem. 268: Hwang TC, Wang F, Yang IC, Reenstra WW (1997) Genistein potentiates wildtype and delta F508-CFTR channel activity. Am. J. Physiol. 273: C988 C Cai Z, Sheppard DN (2002) Phloxine B interacts with the cystic fibrosis transmembrane conductance regulator at multiple sites to modulate channel activity. J. Biol. Chem. 277: In conclusion, we show here for the first time that GnRH-R are expressed in the used airway epithelial cells at the protein level and we propose that GnRH restores some Cl 2 channel function in CF cells. Because some GnRH analogs such as leuprolide, buserelin and goserelin are already used in medical practice as nasal sprays, we intend to test these medications in human primary cells. Acknowledgments We thank the association Gaëtan Salaün, the SFR Scinbios (Brest) and the French foundation Vaincre la Mucoviscidose. We also thank Dr Mohamed Keir (Nanion Technology, Germany) as well as Dr Gérard Sinquin and Dr Philippe Elies of the PIMM facility (Université de Brest, France). Author Contributions Conceived and designed the experiments: PT CF. Performed the experiments: NB SLH CN MLC PT. Analyzed the data: PT NB MK CN FB. Contributed reagents/materials/analysis tools: NB SLH CN MLC. Wrote the paper: PT. 20. Vergani P, Lockless SW, Nairn AC, Gadsby DC (2005) CFTR channel opening by ATP-driven tight dimerization of its nucleotide-binding domains. Nature 433: Rubenstein RC, Egan ME, Zeitlin PL (1997) In vitro pharmacologic restoration of CFTR-mediated chloride transport with sodium 4-phenylbutyrate in cystic fibrosis epithelial cells containing delta F508-CFTR. J. Clin. Invest. 100: Brown CR, Hong-Brown LQ, Biwersi J, Verkman AS, Welch WJ (1996) Chemical chaperones correct the mutant phenotype of the delta F508 cystic fibrosis transmembrane conductance regulator protein. Cell Stress Chaperones 1: Denning GM, Anderson MP, Amara JF, Marshall J, Smith AE et al. (1992) Processing of mutant cystic fibrosis transmembrane conductance regulator is temperature-sensitive. Nature 358: Norez C, Noel S, Wilke M, Bijvelds M, Jorna H et al. (2006) Rescue of functional delf508-cftr channels in cystic fibrosis epithelial cells by the alphaglucosidase inhibitor miglustat. FEBS Lett. 580: Wang Y, Bartlett MC, Loo TW, Clarke DM (2006) Specific rescue of cystic fibrosis transmembrane conductance regulator processing mutants using pharmacological chaperones. Mol. Pharmacol. 70(1): Wang Y, Loo TW, Bartlett MC, Clarke DM (2007) Correctors promote maturation of cystic fibrosis transmembrane conductance regulator (CFTR)- processing mutants by binding to the protein. J. Biol. Chem. 282: Becq F, Mettey Y, Gray MA, Galietta LJ, Dormer RL et al. (1999) Development of substituted Benzo[c]quinolizinium compounds as novel activators of the cystic fibrosis chloride channel. J. Biol. Chem. 274: Trouvé P, Le Drévo MA, Kerbiriou M, Fichou Y, Gillet D et al. (2007) Annexin V is directly involved in cystic fibrosis transmembrane conductance regulator s chloride channel function. Biochim Biophys Acta. 1772: Le Drévo MA, Benz N, Kerbiriou M, Gioux M, Pennec JP et al. (2008) Annexin A5 increases the cell surface expression and the chloride channel function of the DF508-Cystic Fibrosis Transmembrane Regulator. Biochim Biophys Acta 1782: Kawaminami M, Etoh S, Miyaoka H, Sakai M, Nishida M et al. (2002) Annexin 5 messenger ribonucleic acid expression in pituitary gonadotropes is induced by gonadotropin-releasing hormone (GnRH) and modulates GnRH stimulation of gonadotropin release. Neuroendocrinology 75(1): Yao B, Kawaminami M (2008) Stimulation of annexin A5 expression by gonadotropin releasing hormone (GnRH) in the Leydig cells of rats. J Reprod Dev 54(4): Kawaminami M, Uematsu N, Funahashi K, Kokubun R, Kurusu S (2008) Gonadotropin releasing hormone (GnRH) enhances annexin A5 mrna expression through mitogen activated protein kinase (MAPK) in LbetaT2 pituitary gonadotrope cells. Endocr J. 55(6): Cozens AL, Yessi MJ, Kunzelmann K, Ohrui T, Chin L et al. (1994) CFTR expression and Chloride secretion in polarized immortal human bronchial epithelial cells. Am J Respir Cell Mol Biol 10: Bruscia E, Sangiuolo F, Sinibaldi P, Goncz KK, Novelli G et al. (2002) Isolation of CF cell lines corrected at DeltaF508-CFTR locus by SFHR-mediated targeting. Gene Ther. 9: PLOS ONE 13 February 2014 Volume 9 Issue 2 e
142 GnRH Improves Chloride Transport in CF Cells 35. Illek B, Maurisse R, Wahler L, Kunzelmann K, Fischer H et al. (2008) Cl transport in complemented CF bronchial epithelial cells correlates with CFTR mrna expression levels. Cell. Physiol. Biochem. 22: Bebok Z, Collawn JF, Wakefield J, Parker W, Li Y et al. (2005) Failure of camp agonists to activate rescued deltaf508 CFTR in CFBE41o- airway epithelial monolayers. J. Physiol. 569: Lowry OH, Rosebrough NJ, Farr AL, Randall RJ (1951) Protein measurement with the Folin phenol reagent. J. Biol. Chem. 193: Norez C, Antigny F, Becq F, Vandebrouck C (2006) Maintaining Low Ca2+ Level in the Endoplasmic Reticulum Restores Abnormal Endogenous F508del- CFTR Trafficking in Airway Epithelial Cells. Traffic 7: Farre C, Stoelzle S, Haarmann C, George M, Brüggemann A et al. (2007) Automated ion channel screening: patch clamping made easy. Expert Opin Ther Targets 11(4): McKone EF, Emerson SS, Edwards KL, Aitken ML (2003) Effect of genotype on phenotype and mortality in cystic fibrosis: a retrospective cohort study. Lancet 361: Zhang L, Button B, Gabriel SE, Burkett S, Yan Y et al. (2009) CFTR delivery to 25% of surface epithelial cells restores normal rates of mucus transport to human cystic fibrosis airway epithelium. PLoS Biol. 7 e Gerke V, Moss SE (2002) Annexins: from structure to function. Physiol Rev 82: Crompton MR, Moss SE, Crumpton MJ (1988) Diversity in the lipocortin/ calpactin family. Cell 55: Moss SE, Morgan RO (2004) The annexins. Genome Biol 5: Rojas E, Pollard HB, Haigler HT, Parra C, Burns AL (1990) Calcium-activated endonexin II forms calcium channels across acidic phospholipid bilayer membranes. J Biol Chem 265: Faria D, Dahimène S, Alessio L, Scott-Ward T, Schreiber R et al. (2011) Effect of Annexin A5 on CFTR: regulated traffic or scaffolding? Mol Membr Biol. 28(1): Borthwick LA, Botha P, Verdon B, Brodlie MJ, Gardner A et al. (2011) Is CFTR-delF508 really absent from the apical membrane of the airway epithelium? PLoS One 6(8): e Kakar SS, Jennes L (1995) Expression of gonadotropin-releasing hormone and gonadotropin-releasing hormone receptor mrnas in various non-reproductive human tissues. Cancer Lett. 98(1): Tieva A, Stattin P, Wikström P, Bergh A, Damber JE (2001) Gonadotropinreleasing hormone receptor expression in the human prostate. Prostate 47(4): Koushik K, Bandi N, Sundaram S, Kompella UB (2004) Evidence for LHRH- Receptor Expression in Human Airway Epithelial (Calu-3) Cells and Its Role in the Transport of a LHRH Agonist. Pharmaceutical Research 21(6): Reutelingsperger CP, van Heerde W, Hauptmann R, Maassen C, van Gool RG et al. (1994) Differential tissue expression of Annexin VIII in human. FEBS Lett. 349(1): Kawaminami M, Tsuchiyama Y, Saito S, Katayama M, Kurusu S et al. (2002) Gonadotropin-releasing hormone stimulates annexin 5 messenger ribonucleic acid expression in the anterior pituitary cells. Biochem Biophys Res Commun. 291(4): Alzamora R, King JD Jr, Hallows KR (2011) CFTR regulation by phosphorylation. Methods Mol Biol. 741: Baudouin-Legros M, Hinzpeter A, Jaulmes A, Brouillard F, Costes B et al. (2005) Cell-specific posttranscriptional regulation of CFTR gene expression via influence of MAPK cascades on 39UTR part of transcripts. Am J Physiol Cell Physiol 289(5): C1240 C Rowe SM. Verkman AS (2013) Cystic fibrosis transmembrane regulator correctors and potentiators. Cold Spring Harb Perspect Med. 3(7): a PLOS ONE 14 February 2014 Volume 9 Issue 2 e
143 La détection du R-GnRH dans les lignées cellulaires bronchique humain exprimant un CFTR muté F508del suggère que la GnRH synthétique ou ses analogues (agonistes ou antagonistes) puissent avoir une action directe sur cette cible thérapeutique. Une augmentation - après un traitement par la GnRH (10-9 M) des lignées cellulaires (16HBE14o-, CFBE41o-/wt, CFBE41o- et CFBE41o-/F508del) a été observée. Les lignées cellulaires dérivent de cellules bronchiques immortalisées et sont incapables de se différencier en cellules ciliées. Les cultures primaires de cellules épithéliales physiologiques plus proches des tissus in vivo. de la GnRH chez les patients CF, il sera nécessaire de répondre à de nombreuses questions telles que : le R-GnRH est-il retrouvé dans les cultures primaires de cellules épithéliales respiratoires humaines? Un traitement par un analogue de la GnRH augmente-t-il les efflux ioniques dépendants du CFTR? Si oui, quelle(s) voie(s) de signalisation(s) active-t-il et impacte-t- a été de tester un analogue de la GnRH, la buséréline, comme primaires de cellules épithéliales nasales humaines homozygotes pour la mutation F508del. Cet analogue, déjà prescrit en thérapeutique humaine, peut être administré par voie nasale sous forme de spray. Dans un premier temps nous avons vérifié la présence du R-GnRH dans notre modèle cellulairela buséréline sur les efflux ioniques dépendants du CFTR dans les cellules épithéliales nasales humaines exprimant le CFTR muté. la ou les voie(s) de signalisation(s) déclenchée(s) par la fixation dans notre modèle cellulaire et une étude des protéines différentiellement exprimées après traitement a été entreprise. le journal Plos one (PONE-D R1). Ils sont présentés et discutés dans le prochain chapitre de ce manuscrit. 107
144 Partie III. Résultats Article 2 : Etude de la régulation du canal CFTR, impliqué dans la mucoviscidose, par un analogue de la GnRH Avant-propos Les analogues de la GnRH sont utilisés en thérapeutique humaine depuis plus de 30 - dépendant du CFTR suite à une stimulation par la GnRH de lignées cellulaires dérivant exprimant le CFTR-F508del. Dan la GnRH ou de ces analogues dans le propriétés physiologiques plus proche de celles présentées par les tissus analogue de la GnRH, la buséréline, sur des cultures primaires de cellules épithéliales nasales humaines cultivées à partir de prélèvements effectués chez des patients atteints de mucoviscidose (F508del/F508del). Résumé des résultats endogène du R-GnRH dans les cellules épithéliales nasales humaines exprimant le CFTR sauvage ou muté (F508del) par RT-PCR et western blot. La présence du R-GnRH dans nos modèles permet de poursuivre les investigations et - dépendants du CFTR dans des cellules épithéliales nasales humaines exprimant le CFTR-F508del. Les techniques de patch- Ussing une - médiées par le CFTR après une incubation des cellules épithéliales CF avec la buséréline à M pendant 2h. Pour expliquer les effets observés sur la fonction du canal CFTR, la concentration intracellulaire en AMPc a été évaluée et une électrophorèse bidimensionnelle entreprise. cellules CF peut expliquer, en partie, - plus résultats met en évidence 6 protéines différentiellement exprimées (ATPA, NDUV1, TCPZ, GRP94, la serpine B3 et VDAC2) suite à la stimulation des cellules épithéliales nasales humaines de deux patients CF. 108
145 Conclusion Le R-GnRH est exprimé dans les cellules épithéliales bronchiques (saines) et nasales (saines et CF) humaines. Une amélioration de la sortie des ions Cl - dépendantes du canal CFTR est observée lorsque les cellules épithéliales nasales sont traitées avec la buséréline (2h, M) certaines protéines comme par exemple les protéines chaperons TCPZ et GRP94 et les protéines de la membrane mitochondriale ATPA et NDUV1. 109
146 110
147 111
148 112
149 Cell cultures 113
150 Histology Immunocytochemistry RNA extraction and RT-PCR 114
151 Protein extraction and Western Blot analysis Patch-clamp analysis Ussing Chamber camp measurements Two-dimensional Gel Electrophoresis (2-DE)and Mass Spectrometry (MS) 115
152 Statistics 116
153 in vivo 117
154 118
155 119
156 120
157 Proc Natl 121
158 .. Proc Natl Acad Sci USA 122
159 123
160 124
161 125
162 126
163 127

164 * 128





165
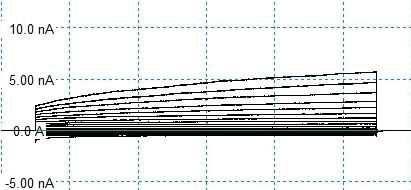



166 130
167 ) 131
168 132


169 133



170 134
171 Partie IV. Discussion enjeu majeur dans la recherche sur la mucoviscidose. Parmi ces modulateurs nous retrouvons les potentiateurs, les correcteurs et les stabilisateurs. menée dépendants du bronchique humain exprimant le CFTR-wt (16HBE et CFBE41o-/wt) ou le CFTR-F508del (CFBE41o- et CFBE41o-/F508del), après traitement à la GnRH pendant 1h à 10-9 M (Benz et 2014). Les cultures primaires des cellules humaines possèdent des propriétés physiologiques plus proches des tissus Cpourquoi les études conduites dans des cellules non immortalisées et non transfectées sont considérées comme le «gold standard» pour la recherche sur la mucoviscidose (Harris et 2004). Pour mieux comprendre les mécanismes physiopathologiques impliqués lors du développement des atteintes pulmonaires dans la mucoviscidose (lésions, fibrose), des études à partir de cellules provenant de prélèvements de l nécessaires (Mosler et 2008). Les tissus utilisés pour établir des cultures primaires de cellules épithéliales des voies aériennes sont généralement issus de trachées, de bronches ou de bronchioles obtenues lors de chirurgie, e récupérer une quantité importante de cellules pour la mise en culture mais cela nécessite un acte chirurgical sous anesthésie générale complications. Des biopsies des cornets ou des polypes nasaux sont également utilisés facilement, des méthodes atraumatiques et moins invasives telles que les brossages nasaux ont été mises en place (Bridges et 1991a). Cette technique, moins chère et plus facile à pratiquer chez les patients, a été intr de la mucoviscidose. Elle est bien tolérée et possible même chez les enfants (Mosler et 2008). Elle permet des prélèvements de cellules nasales répétés chez un même donneur (Dimova et 2005). considéré comme représentatif de 2008). de CFTR dans les cellules épithéliales respiratoires humaines a considérablement avancé lorsque leur obtention par brossage nasal a été développée. En 2004, Harris et collaborateurs ont décrit une technique fraîchement isolées à partir de brossages nasaux effectués chez des patients atteints de mucoviscidose et porteurs homozygotes de la mutation F508del (Harris et 2004). 135
172 une étude de de Courcey et collaborateurs, les cultures primaires de cellules épithéliales respiratoires obtenues après des brossages nasaux chez des patients F508del possèdent des telles que le patch-clamp automatique 2012). Ces cultures sont donc des systèmes intéressants pour tester de nouvelles drogues. épithéliales nasales humaines a dirigé notre choix vers analogue de la GnRH sur le canal CFTR impliqué dans la mucoviscidose. La substitution des acides aminés en position 6 et 10 de la chaîne en acides aminés de la GnRH synthétique permet aux analogues de de présenter une plus forte affinité pour le R-GnRH dans les cellules gonadotropes, une meilleure résistance à la dégradation enzymatique et un temps de demi-vie prolongé (pour revue : Chillik et Acosta 2001). de la GnRH présentant de nombreux avantages, nous avons décidé de choisir la buséréline. Dans un premier temps nous avons validé son utilisation sur nos CFTR-F508del (CFBE41o-/F508del). Les manipulations de patch-clamp nous ont permis ut comme la GnRH synthétique, une incubation des CFBE-F508del pendant 1h avec la buséréline (10-9 M) entraîne une augmentation des efflux de Cl - par le canal CFTR. Par la suite, nous avons dû nous assurer de la présence du R-GnRH dans les cultures primaire, nous rapportons pour la première fois la présence du messager (RT-PCR) et de la protéine (western blot) R- GnRH dans des cultures primaires de cellules épithéliales respiratoires saines ou mucoviscidosiques. L engendrés par la buséréline sur les sécrétions ioniques dépendantes du canal CFTR. Comme dit précédemment, d Courcey et collaborateurs en 2012, il est possible les cultures primaires de cellules épithéliales nasales collectées par brossage pour les manipulations de patch-clamp automatique (de Courcey et 2012). Lors de cette technique nous nous sommes assurés que les courants mesurés correspondent à des courants Cl - dépendants du CFTR en utilisant des activateurs (forskoline et génistéine) et un inhibiteur (CFTR inh172) spécifique du canal. Les résultats de patch clamp révèlent une augmentation significative de - dans les cellules épithéliales nasales humaines CF après deux ou quatre heures de traitement, par rapport aux cellules nasales CF non traitées. - médié 136
173 par le CFTR bservé. Une étude menée par Koushik et collaborateurs, en 2003, a line) pendant 1h avec des fractions membranaires de cellules de carcinome pulmonaire, Calu-1, entraîne la dégradation 2003)., le temps de demi-vie de 300 µg de buséréline administrée en spray nasal est de 66 minutes (Saito et 1985). Dllik et nasale présente des différences inter-individuelles très importantes et une réadministration 2 à 4 fois par jour peut-être nécessaire pour que le traitement soit efficace (Chillik et Acosta, 2001). Lors de notre étude, il - après 8h de traitement. Un des avantages de la culture primaire est la capacité des cellules épithéliales nasales obtenue par brossage à se différencier en un épithélium pseudostratifié présentant les caractéristiques morphologiques et bioélectriques transépithéliale est mesurée avec la présence de jonctions serrées et de cellules ciliées. s obtenus par brossage chez des patients (N=4), la culture ALI a été mise en place au laboratoire. Pour, la résistance transépithéliale (TEER) a été mesurée dans les cultures primaires de cellules épithéliales nasales humaines CF après 4 immunohistochimie sur membrane entière a permis de vérifier la présence de jonctions serrées (ZO-1), de cellules à mucus (Muc-5AC) et de cellules ciliées (-tubuline), caractéristiques des épithéliums différenciés. courants traversant les épithéliums. Pour nous assurer que nous mesurons des courants dépendants du CFTR nous avons utilisé les mêmes activateurs et inhibiteur patchclamp. épithéliums reconstitués de trois patients CF. Une augmentation faible mais significative des courants Cl - dépendants du CFTR, après e, a été mesurée après deux ou quatre heures de traitement à la buséréline (10-12 M). Par ailleurs, certains épithéliums différenciés après 2h traitement. drogues. Les épithéliums différenciés,, sont utilisés comme modèle pour la mesure de cette résistance mais il est difficile de déterminer une valeur de référence pour les épithéliums reconstitués. En effet, il existe de grandes variabilités inter- et intra-individuelles. Les résistances transépithéliales mesurées pour un même patient peuvent être différentes suivant les inserts. Pour contourner cet inconvénient, Tosoni et collaborateurs conseillent de mesurer les TEEms différenciés par donneur et par condition pour 137
174 pouvoir interpréter les résultats (Tosoni et 2016). Il est à noter que la culture primaire après une grande quantité de cellules lors du prélèvement. De plus, après la remise en culture, des sphéroïdes (amas de cellules en suspension) apparaissent au sein de la flasque, ce qui fait que les cultures primaires ne peuvent supporter que deux ou trois passages au maximum (Hull et Harris, 1994). par Tosoni et collaborateurs, par condition et par prélèvement. courants Cl - dans les cultures cellulaires de patients CF, nous avons voulu savoir quel(s) mécanisme(s) cellulaire(s) entrai(en)t en jeu. Dans les cellules gonadotropes, la fixation de la GnRH sur son récepteur active majoritairement les voies intracellulaires dépendantes de la sous-unité G q11, activant la PLC, et dans une moindre mesure celles déclenchées par la sous-unité G Cheng et Leung, 2005 ; Naor, 2009 ; Liu et 2002). Cependant, dans les tissus extra-hypophysaires la part des voies de signalisation mise en place La voie de signalisation couplée à la sous-unité G est intéressante à explorer, AMPc mesuré la concentration intracellulaire en AMPc ([AMPc] i) après traitement à la buséréline (2h, M). Nous avons noté une augmentation significative de la [AMPc] i pour les cultures de cellules issues de deux patients CF après traitement à la buséréline. D bidimensionnelle (2-D) à partir de lysats protéiques totaux issus des cellules nasales de deux patients CF non traitées et traitées à la buséréline. Cette analyse pourrait mettre en évidence quelques pistes quant aux voies activées suite à la sur son récepteur ainsi que de nouvelles cibles thérapeutiques, serait -D a conduit à sélectionner les protéines candidates suivant leur sur- ou sous-expression après traitement. 11 protéines ont été identifiées comme différentiellement exprimées chez un premier patient et 14 chez un deuxième patient. L (SM), protéomique de Paris Sud-Ouest, a permis des prot -. Cette analyse a mis en évidence la sous-expression de la protéine ENPL (ou GRP94). GRP94 est une protéine chaperon du système de dégradation du RE (ERAD) (Singh et 2008). Elle ee (Gomes-Alves et 2010 ; Ma et 138
175 Hendershot, 2004). Elle est associée à la forme B de la protéine CFTR (Singh et 2008). Une étude de Pankow et collaborateurs a montré, en 2015, que le CFTR-F508del interagit avec GRP94 (Pankow et 2015). Dans le cadre de notre étude, lorsque les cultures primaires de cellules nasales sont traitées à la buséréline, une sous-expression de Grp94 est observée ce qui pourrait indiquer que la protéine CFTR-F508del échappe au système de contrôle du RE. La protéine TCPZ est surexprimée suite au traitement. Cette protéine est une sous-unité du complexe TCP1 ( ), aussi connu sous le nom de CCT () (Yue et 2009). Wang et collaborateurs ont montré en 2006 que le complexe TCP1 interagit avec la protéine CFTR (Wang et 2006). Cuellar et collaborateurs, en 2008, ont déterminé une association entre les protéine chaperons Hsp70 et le complexe CCT (Cuellar et 2008). Le complexe CCT permet aux protéines malformées libérées par Hsp70 une conformation correcte (Cuellar et 2008). observations, aiderait les protéines CFTR-F508del malformées, ayant échappées au système de contrôle qualité du RE, à acquérir une conformation correcte ciliées. Les protéines ATPA () et NDUV1 ( []) sont sur-exprimées après le traitement à la buséréline (2h, M) chez les patients CF. Ces protéines sont présentes dans les complexes de la chaîne respiratoire retrouvée dans la membrane des mitochondries. La chaîne respiratoire permet la tion oxydative et comprend 5 complexes : les complexes synthase. La protéine NDUV1 appartient au complexe I qui fixe le NADH. complexe I est modulée par le CFTR. Sa fonction est diminuée dans les lignées cellulaires - 16HBE) (Valdivieso et 2012). NDUV1 est surexprimée, après traitement à la buséréline, nous pouvons corréler cette observation à de la fonction de la protéine CFTR-F508del par le traitement. ATPA appartient à la sous-unité catalytique F1 complexe permet la synthè de protons produit tout au long de la chaîne respiratoire parée dans les cellules exprimant le CFTR-F508del (Valdivieso et Santa-Coloma, 2013 ; Atlante et 2016). La surexpression des protéines NDUV1 et ATPA suggère une activation de la e. Le canal CFTR est un canal Cl - AMPc-dépendant présent dans les cellules épithéliales polarisées (Gadsby 2006). 139
176 La production plus élevée conduit activant le canal CFTR. du traitement sur la fonction du canal CFTR. Pour vérifier cette hypothèse, il serait intéressant de mesurer le de SM la protéolyse appelée serpine B3 (Jeanson et 2014). Les serpines sont des inhibiteurs des protéases à sérine et cystéine impliqués epliement des protéines (Gettins, 2002). Dans le poumon, la surexpression de la protéine serpine B3 induit une prolifération des cellules épithéliales conduisant à une fibrose pulmonaire (Lunardi et 2011). La sous-expression de la serpine B3 après traitement - mais pourrait être une cible pour diminuer le risque de fibrose pulmonaire. Enfin, lues de la GnRH se fait principalement par injection ou par spray nasal. Par voie orale, ils sont dégradés par les enzymes protéolytiques gastro-intestinales. La voie sous-cutanée est la plus commune et se caractérise par une forte biodisponibilité avec peu de variabilité inter- rapidement absorbé et la concentration plasmatique est maintenue pour quelques heures, donc une seule injection par jour est suffisante. Dans le cas de la voie intranasale, utilisée pour les agonistes moins hydrophobes tels que la nafaréline et la buséréline, les analogues peuvent être dégradés par des enzymes présentes dans le mucus et une importante va entre les patients est observée. Il faut donc administrer la drogue deux à quatre fois par jour (Chillik et Acosta 2001). recherche pour des pathologies telles que. de drogues. La muqueuse nasale possède une surface relativement importante due à un nombre important de microvillosités, une perméabilité relativement importante pour des composés hydrophiles ou lipophiles (inférieurs à 1000 Da). Elle est aussi caractérisée par une gas drogues au niveau nasal (Dimova et 2005). La buséréline existe sous la forme de spray nasal et pourrait cibler les voies respiratoires dans ln utilisation pour le traitement de la mucoviscidose. Le R- 140
177 -GnRH dans les cellules épithéliales bronchiques CF humaines et étudier - dépendant du CFTR. Cependant, inhibition de la sécrétion des gonadotropines (LH et FSH) et des hormones sexuelles par les analogues de la GnRH rapportés des symptômes principalement rencontrés lors de la ménopause et Si une utilisation de la buséréline est envisagée, de nombreux paramètres devront être pris en compte. La dose, dministrations ainsi que le moment et étudiés -bronchique est i pourraient fortement diminuer son effet. 141
178 142
179 Le Mg 2+ de M. Il est essentiel pour de nombreuses réactions biochimiques et fonctions - Le [Mg 2+ ] i est principalement régulé par un canal ubiquitaire appelé TRPM7 (Transient Receptor Potential Melastatin 7) (Ryazanova et al., 2010). TRPM7 est impliqué dans de développement embryonnaire. Ce canal est lui-même régulé par le Mg 2+ intracellulaire et le Mg-ATP. concentration intracellulaire des ions divalents, Ca 2+ et Mg 2+ CFTR (Bargon et al., 1992). Pour cela, ils ont utilisé les ionophores (A23187 et ionomycine) et la CFTR dépend de la concentration intracellulaire en Mg 2+. De plus, la protéine CFTR est aussi régulée par le Mg 2+ intracellulaire (Bargon et al., 1992; Li et al., 1996 ; Ikuma et Welsh, 2000). Le canal CFTR appartient à la famille des transporteurs ABC et présente des NBDs qui contiennent des motifs très conservés, dont les séquences Walker A et Walker B. Ces séquences constituent le noyau catalytique et -facteur de la catalyse, le Mg 2+ (ter Beek et al., 2014). Le Mg 2+ les NBDs. Une diminution de la concentration en Mg 2+ fermeture du canal (Bargon et al., 1992). Le Mg 2+ du canal CFTR (Dousmanis et al., -ATP sur la fonction du canal CFTR e par le Mg 2+ (pour revue Hwang et Kirk, 2013). Mg 2+ est aussi diminuée (Li et al., 1996). Enfin, le CFTR est un canal AMPc- In vivonylate cyclase qui catalyse (Choi et al., 2007 ; Tarran et al., 2006). Cette réaction nécessite la présence de Mg
180 es par le Mg 2+ intracellulaire, l TRPM7, principal régulateur de la concentration intracellulaire de Mg 2+ ([Mg 2+ ] i). Dans un premier temps, nous avons regardé la [Mg 2+ ] i présente dans des lignées cellulaires transfectées stablement avec le CFTR sauvage (wt) ou muté (G551D ou F508del) puis nous avons dans nos modèles cellulaires. Ensuite, nous avons étudié la fonction et la régulation de TRPM7 par le Mg 2+ dans nos modèles exprimant le CFTR wt ou muté. Afin de déterminer tre les canaux TRPM7 et CFTR nous avons étudié la fonction du canal CFTR après traitement des cellules par un activateur ou un inhibiteur de TRPM7. le publié dans le journal Cellular and Molecular Life Science, en
181 : La fonction et la régulation de TRPM7 ainsi que la [Mg 2+ ] i sont altérées dans les cellules exprimant les mutations CFTR- F508del ou CFTR-G551D La mucoviscidose est due à des mutations du gène CFTR codant la protéine CFTR. La protéine CFTR est régulée par des interactions protéines/protéines ainsi que par certains ions tels que le Ca 2+ et le Mg 2+. Le principal régulateur de la concentration intracellulaire en Mg 2+ est le canal TRPM7. 2+, Mg 2+ et de métaux divalents. Il est appelé chanzyme car il possède un domaine sérine- -kinase et une activité canal. Il est lui-même régulé par le Mg 2+ et le Mg-ATP. 2+ dans des cellules exprimant le CFTR wt ou muté (F508del et G551D). La fonction du canal CFTR est étudiée alors que la fonction de TRPM7 est modulée par des activateurs ou inhibiteurs spécifique de TRPM7. La [Mg 2+ ] i est significativement plus faible dans les cellules exprimant le CFTR muté par rapport aux cellules exprimant le CFTR-wt. La protéine TRPM7 est beaucoup plus exprimée dans les cellules exprimant le CFTR muté (G551D ou F508del) par rapport aux cellules CFTR-wt et sa localisation est différente suivant nos modèles cellulaires. Un liserai de protéine TRPM7 à la membrane plasmique apparait pour les cellules exprimant le CFTR-wt et le CFTR-F508del mais il est absent pour les cellules CFTR-G551D. Cependant, nous observons une expression vésiculaire de TRPM7 Ensuite, la fonction du canal TRPM7 à la membrane plasmique a été analysée par la technique de patch clamp. Elle est fortement réduite dans les cellules présentant le CFTR muté. augmentée dans les cellules exprimant le CFTR-F508del ou CFTR-, un activateur de TRPM7. Mais il y a une 145
182 diminution de la fonction du canal CFTR dans les cellules CFTR-F508del et une augmentation dans les cellules G551D. Les [Mg 2+ ] i et [Ca 2+ ] i sont modifiées dans les cellules exprimant le CFTR muté. de Ca 2+ pour celles exprimant le CFTR-F508del. TRPM7 sont altérées dans les cellules exprimant le CFTR-F508del ou le CFTR-G551D. Il existerait un lien fonctionnel entre [Mg 2+ ] i dans les cellules CF. TRPM7 serait directement impliqué dans la physiopathologie de la mucoviscidose et le Naltriben serait un nouveau potentiateur du CFTR-G551D. 146

183 Cell. Mol. Life Sci. DOI /s Cellular and Molecular Life Sciences ORIGINAL ARTICLE Function and regulation of TRPM7, as well as intracellular magnesium content, are altered in cells expressing DF508-CFTR and G551D-CFTR F. Huguet 1,2 M. L. Calvez 1,2,3 N. Benz 1,3 S. Le Hir 1,4 O. Mignen 1,2 P. Buscaglia 1,2 F. D. Horgen 5 C. Férec 1,2,4,6 M. Kerbiriou 1,2 P. Trouvé 1 Received: 8 June 2015 / Revised: 14 January 2016 / Accepted: 25 January 2016 Springer International Publishing 2016 Abstract Cystic fibrosis (CF), one of the most common fatal hereditary disorders, is caused by mutations in the cystic fibrosis transmembrane conductance regulator (CFTR) gene. The CFTR gene product is a multidomain adenosine triphosphate-binding cassette (ABC) protein that functions as a chloride (Cl - ) channel that is regulated by intracellular magnesium [Mg 2? ] i. The most common mutations in CFTR are a deletion of a phenylalanine residue at position 508 (DF508-CFTR, % of CF phenotypes) and a Gly551Asp substitution (G551D-CFTR, 4 5 % of alleles), which lead to decreased or almost abolished Cl - channel function, respectively. Magnesium ions have to be finely regulated within cells for optimal expression and function of CFTR. Therefore, the melastatin-like transient receptor potential cation channel, F. Huguet and M. L. Calvez participated to the same extend to the present work. & C. Férec claude.ferec@univ-brest.fr & P. Trouvé pascal.trouve@univ-brest.fr Inserm, UMR1078, 46, rue Félix le Dantec, CS 51819, Brest Cedex 2, France Faculté de Médecine et des sciences de la santé, Université de Bretagne Occidentale, Brest 29200, France Association G. Saleun, Brest 29218, France Laboratoire de Génétique Moléculaire, Hôpital Morvan, C.H.U. Brest, Brest 29200, France Laboratory of Marine Biological Chemistry, Department of Natural Sciences, Hawaii Pacific University, Kaneohe, HI 96744, USA Etablissement Français du Sang - Bretagne, Brest 29200, France subfamily M, member 7 (TRPM7), which is responsible for Mg 2? entry, was studies and [Mg 2? ] i measured in cells stably expressing wildtype CFTR, and two mutant proteins (DF508-CFTR and G551D-CFTR). This study shows for the first time that [Mg 2? ] i is decreased in cells expressing DF508-CFTR and G551D-CFTR mutated proteins. It was also observed that the expression of the TRPM7 protein is increased; however, membrane localization was altered for both DF508del-CFTR and G551D-CFTR. Furthermore, both the function and regulation of the TRPM7 channel regarding Mg 2? is decreased in the cells expressing the mutated CFTR. Ca 2? influx via TRPM7 were also modified in cells expressing a mutated CFTR. Therefore, there appears to be a direct involvement of TRPM7 in CF physiopathology. Finally, we propose that the TRPM7 activator Naltriben is a new potentiator for G551D-CFTR as the function of this mutant increases upon activation of TRPM7 by Naltriben. Keywords Cystic fibrosis TRPM7 CFTR G551D-CFTR DF508-CFTR Naltriben Introduction Cystic fibrosis (CF), one of the most common fatal hereditary disorders in Caucasians, is caused by mutations in the cystic fibrosis transmembrane conductance regulator (CFTR) gene on chromosome 7 [1 3]. Translation of CFTR mrna results is a 1480 residue with a predicted molecular mass of 168 kilodaltons (kda) [2]. CFTR is a multidomain adenosine triphosphate-binding cassette (ABC) protein responsible for the regulation of chloride (Cl - ) transmembrane transport, as well as many other ions such as halides (permeation sequence P I [ P Br [ P Cl [ P F ; [4]) and polyatomic anions
184 F. Huguet et al. [5], and also to some small uncharged molecules [6]. Also, CFTR function is critical in maintaining fluid homeostasis, airway fluid clearance and airway submucosal glands secretion [7, 8]. The most common mutation causing CF is a deletion of a phenylalanine residue at position 508 (DF508-CFTR), which is responsible for % of CF phenotypes worldwide [8 10]. The DF508-CFTR protein is misfolded, retained in the endoplasmic reticulum (ER) and prematurely degraded. The limited amount of DF508-CFTR protein that reaches the plasma membrane is unstable, resulting in a rapid redirection from endosomal recycling towards lysosomal delivery and degradation [11 13]. Another mutation, Gly551Asp (G551D) also called the Celtic mutation, accounts for 4 5 % of alleles, and exhibits normal protein expression and cell surface localization. Nevertheless, it is associated with a severe form of CF pathology as Cl - channel function is lacking [14 17]. Under normal conditions, wildtype CFTR (WT-CFTR) is a Cl - channel activated by protein kinases A and C (PKA and PKC) and deactivated by dephosphorylation by a high number of protein phosphatases [15, 16]. WT-CFTR is also regulated by protein protein interactions involving synaptosomal-associated protein 23 (SNAP23) and syntaxin 1A, Na? /H? exchanger regulatory factor (NHERF), CFTR-associated protein of 70 kda (CAP70), CFTR associated ligand (CAL) and annexin A5 [20 28]. CFTR is also regulated by divalent cations, the most studied being calcium (Ca 2? ) ions. Indeed, it was clearly reported that Ca 2? homeostasis is altered in CF [29]. An abnormal spatial diffusion of Ca 2? in airway epithelial cells derived from CF patients with the DF508-CFTR mutation was observed, due to an accumulation of inositol 1,4,5- trisphosphate (IP3) in the ER [30] and to dysfunction Ca 2? uptake in mitochondria [31]. An increased Ca 2? uptake is a hallmark of CF [32], but when the function of CFTR is restored, for example using Miglustat, Ca 2? homeostasis is normalized [33]. Intracellular magnesium ([Mg 2? ] i ) is also important for the regulation of CFTR. The use of ionophore A23187 or ionomycine in human bronchial cells increases [Mg 2? ] i and leads to a time- and dose-dependent decrease in the expression of CFTR mrna [34]. By contrast, decreased [Mg 2? ] i leads to reduced opening and closing rates of the CFTR channel [34]. In fact, [Mg 2? ] i controls the closing rate of CFTR, likely in conjuction with ATP. It is well established that ATP hydrolysis by nucleotide binding domain 2 (NBD2) of CFTR is decreased when [Mg 2? ] i is also decreased [35], the role of Mg-ATP upon CFTR s function being more efficient than that of ATP alone [36]. Due to the fact that Mg 2? has to be finely regulated within cells for optimal CFTR expression and function, our aim was to study the melastatin-like transient receptor 123 potential cation channel, subfamily M, member 7 (TRPM7) in non-transfected Hela cells (NT) and in cells expressing WT-CFTR, DF508-CFTR and G551D-CFTR. TRPM7, which is a member of the melastatin-like transient receptor potential cation channel, subfamily M (TRPM) family, is ubiquitously expressed and possesses both an ion pore channel and a functional serine/threonine a-kinase domain to provide cellular influx for Ca 2?, Mg 2? and divalent metals [37 40]. Being a channel and having an enzymatic activity, TRPM7 is often called chanzyme. The TRPM7 protein, which is the main cellular regulator of [Mg 2? ] i [41], is slightly more permeable to Mg 2? than to Ca 2? and is regulated by Mg 2? or Mg-ATP [38, 42 44]. Indeed, TRPM7 has the following selectivity profile for divalent cations: Zn 2? & Ni 2? [[ Ba 2? [ Co 2? [ Mg 2? C Mn 2? C Sr 2? C Cd 2? C Ca 2? [43]. A remarkable finding being that trace divalent metal ions are significantly more permeable through TRPM7 than the dominant physiological divalent metal ions Ca 2? and Mg 2? [43]. Therefore, we studied the expression, the function and the channel regulation of TRPM7 by [Mg 2? ] i in NT cells and in cells expressing WT-CFTR or mutated CFTR (DF508-CFTR and G551D-CFTR). The function of CFTR was further studied when TRPM7 function was modulated by specific drugs. We show for the first time that [Mg 2? ] i is decreased in cells expressing DF508-CFTR and G551D-CFTR. The protein expression of TRPM7 was increased in these cells whereas its function and Mg 2? regulation were decreased. In order to highlight a possible functional link between TRPM7 and CFTR, TRPM7 activity was increased in cells expressing DF508-CFTR and G551D-CFTR by the use of Naltriben and the function of CFTR was studied. We found that the activation of TRPM7 resulted in a decreased CFTR function in DF508-CFTR cells and in a highly increased function in cells expressing G551D-CFTR. Mg 2? and Ca 2? flux through TRPM7 were found to be modulated in cells expressing mutated CFTR. We conclude that the link between TRPM7 and CFTR varies with the type of CFTR mutation. Finally, we propose that Naltriben is a new potentiator of G551D-CFTR with potential implications for CF therapy. Indeed, the TRPM7 activator Naltriben rescues the altered function associated with the G551D-CFTR mutation. Materials and methods Cell culture, quantitative PCR and western blotting HeLa cells stably expressing WT-CFTR, DF508-CFTR and G551D-CFTR were obtained by lentiviral infection in which the cdna encoding the human wildtype CFTR (WT-CFTR, Transgene SA, ptg5985, France, access 148
185 Function and regulation of TRPM7, as well as intracellular magnesium content, are altered in Genbank No. M28668) was used, before and after site directed mutagenesis. Hela cells stably expressing WT- CFTR, DF508-CFTR or G551D-CFTR were characterized previously [45]. Cells were cultured in Dulbecco s Modified Eagle Medium (DMEM; E Healthcare, USA) plus 10 % calf serum in the presence of blasticidine (2 lg/ml). Total RNA was extracted from cells using the RNeasy Plus Mini kit (Qiagen, France), according to the manufacturer s instructions, and the quality was assessed using an Agilent 2100 Bioanalyzer (Agilent, France). RNA was further quantified using a nanophotometer for further use in Reverse Transcription Polymerase Chain Reaction (RT- PCR). RT-PCR was performed using Superscript II (InVitrogen, France). Quantitative PCR was performed using a Quantitect SYBR Green kit (Qiagen, France) and 3 lg of cdna as tempate. TRPM7 primers were used as following: forward CTTATGAAGAGGCAGGTCATGG and reverse CATCTTGTCTGAAGGACTG. Actin primers were forward GCATGGAGTCCTGTGGCATCC and reverse GAGGCCAGGATGGAGCCG. Primers were tested for efficiency (E) using a standard curve analysis. E values were and % (R 2 [ 0.98) for TRPM7 and actin, respectively. Optimal annealing temperature for the qpcr assay was determined by testing a range of temperatures above and below the calculated Tm (54 C) of the primers. PCRs yielded 213 and 238 bp amplicons for TRPM7 and actin, respectively [46]. Reactions for TRPM7 were carried out with the following parameters: initial denaturation at 95 C for 15 min followed by 40 cycles of denaturation at 95 C for 30 s, annealing at 54 C for 30 s and extension at 72 C for 30 s. Relative quantification of the transcripts were assessed in a one-step format (300 ng total RNA) using a QuantiTect SYBR Green RT-PCR Kit (Qiagen) with an ABI PRISM 7700 sequence detection system. The cells were lyzed in 50 mm Tris HCl, 100 mm NaCl, 0.1 % SDS, in the presence of 1.1 lm leupeptin, 0.7 lm aprotinin, 120 lm phenylmethanesulfonylfluoride (PMSF), 1 lm iodoacetamide, 0.7 lm pepstatin and 1 mm diisopropyl phosphorofluoridate (DIFP). Protein concentrations were determined using Lowry s methodology with bovine serum albumin as a standard [47]. Equal amounts of total protein for each sample (50 lg) were subjected to 4 12 % gradient SDS/PAGE (InVitrogen, France). After blotting, the polyvinylidene difluoride (PVDF) membranes were incubated with anti-trpm7 (1/1000; Abcam, France, [48]) or anti-glycerol-3-phosphate Dehydrogenase (G3PDH, 1/7000) antibodies and with secondary antibody (anti-mouse, abcam ab6820, 1/20,000). Visualization was performed using an ECL? detection kit (Amersham, France). Densitometric analysis of the signals was performed using a GelDoc 2000 apparatus (Bio-Rad, France) and each value was normalized to the total amount of loaded proteins per lane determined by Coomassie blue staining of the membrane or to the G3PDH, which was estimated by densitometric analysis. Measurements of [Mg 21 ] i with Mag-Fura-2-AM Cells were grown in glass bottomed, black wall 96-well plates (BD Falcon) for 1 day before experiments. They were washed three times with normal buffered salt solution (NBSS) containing: 135 mm NaCl, 5.4 mm KCl, 1.2 mm CaCl 2, 1 mm MgCl 2, 0.33 mm NaH 2 PO 4, 5 mm Glucose and 10 mm HEPES (ph 7.4). Measurement of the background fluorescence was performed after the last wash. Cells were further incubated for 30 min at 37 C in NBSS containing 5 lm Mag-Fura-2-AM (Molecular Probes, France) and 0.1 % Pluronic F-127. Cells were washed twice with NBSS containing 0.1 mm K 2 EGTA in place of 1.2 mm CaCl 2 for 20 min at 37 C and measurement of fluorescence was performed. All fluorescence measurements were performed on Varioskan Flash (Thermo Scientific) at 37 C. Excitation wavelengths were 335 nm and 375 nm and the emission wavelength was 505 nm (12 nm bandwidth). All measurements of background fluorescence were subtracted from fluorescence measurements for each wavelength. [Mg 2? ] i was calculated from the ratio of fluorescence at the two excitation wavelengths described above, using a dissociation constant (K d ) of 1.9 mm, for the Mag-Fura-2-AM/Mg 2? complex. Fluorescence at 335 (F(335)) and 375 (F(335)) nm were used to determine the R value (R = F(335)/F(375)). All values of the measured R were normalized to the standard R value with identical optics and converted to [Mg 2? ] i using the standard equation [Mg 2? ] i = K d 9 (R - R min / R max - R). Cells were permeabilized with 20 lm digitonin and the Rmax value was obtained by the addition of 50 mm MgCl 2 in absence of Ca 2. Rmin was determined by chelating Mg 2? with addition of 100 mm EDTA (ph 7.2). Measurements of Ca 21 influx through TRPM7 Cells were plated (30,000 cells/well) in glass bottomed, black wall 96-well plates (BD Falcon) in DMEM containing 10 % calf serum without blasticidine during 1 day before experiment. The culture medium was removed and replaced with loading buffer containing: 137 mm NaCl, 4.7 mm KCl, 1.8 mm CaCl 2, 560 lm MgCl 2, 1 mm Na2HPO4, 5.5 mm Glucose, 10 mm HEPES (ph 7.4) and the Fura-2 QBT Calcium dye (Molecular Devices) during 60 min at 37 C in 5 % CO 2 atmosphere. The Fura-2 QBT TM Calcium Kits contain Fura-2, AM, a cell permeable calcium indicator that is ratiometric and UV-light excitable. Loading buffer was removed and replaced by the same buffer without Fura-2 and Ca 2? (1.8 mm CaCl 2 was
186 F. Huguet et al. replaced by 100 lm EDTA). Fluorescence quenching measurements (excitation wavelength: 360 nm; emission wavelength: 510 nm) were realized using Flextation3 plate reader (Molecular Devices; SoftMax Pro v5.4) at 37 C [49]. Basal fluorescence at 360 nm excitation was read during 2 min before the addition of 2 mm MnCl 2 to follow constitutive calcium entry for 13 min. In order to assess the implication of TRPM7 in Ca 2? entries, activator Naltriben and inhibitor Waixenicin A were added 15 min before starting the reading (50 and 15 lm, respectively). Slope values were first normalized to average initial fluorescence (F 0 ) measured between 0 and 2 min (F t = (DF/F 0 ; DF = F t(360) F 0 ) and then normalized to control values (WT-CFTR = 100 %). The slope of the quenching which was estimating Ca 2? entry was calculated with GraphPad Prism 5 and presented as average ± SD. Different conditions were compared using t test. Patch clamp analysis Patch-clamp experiments were performed with an automatic electrophysiology workstation, Port-a-Patch (Nanion Technologies GmbH, Germany), coupled to an external amplifier unit HEKA EPC-10 [28, 50]. Whole-cell patchclamp recordings were performed at room temperature on isolated cells. Before recording, the culture medium was replaced by the external solution contained the following: 140 mm NaCl, 2 mm CaCl 2, 1 mm MgCl 2, 10 mm HEPES and 5 mm D-glucose monohydrate [41, 51]. The final ph value was adjusted to 7.4 with NaOH and the osmolarity was 298 mosmol. The internal solution contained the following: 50 mm CsCl, 10 mm NaCl, 60 mm Cs-Fluoride, 20 mm EGTA and 10 mm HEPES/CsOH (ph 7.2; 285 mosmol). Inhibition of TRPM7 was performed by increasing the free [Mg 2? ] i from 0 to 200 lm, using MaxChelator software available at stanford.edu/*cpatton/webmaxcs.htm, for Mg 2? calculation [41, 52]. In some experiments, seals were enhanced using the seal enhancer solution containing: 80 mm NaCl, 3 mm KCl, 10 mm MgCl 2, 35 mm CaCl 2, 10 mm HEPES (298 mosmol). All voltages were corrected for a liquid junction potential of 10 mv between external and internal solutions. For TRPM7 measurements, currents were elicited by a ramp protocol from -100 to?100 mv over 50 ms acquired at 0.5 Hz and a holding potential of 0 mv, as previously described [41, 52]. Voltage-clamp pulses were generated and data were captured using the Patchmaster program (Nanion Technologies GmbH, Germany). Outward currents at?80 mv, normalized to cell size ranging from 20 to 35 pf, and plotted against time, were recorded. For CFTR measurements, voltage-dependent properties were assessed by applying voltage steps of 100 ms duration from a holding potential of -80 mv to 123 test potentials ranging from -80 to?80 mv, with 10 mv increments [28]. Activation and inhibition of TRPM7 were obtained by Naltriben methanesulfonate (50 lm) and Carvacrol (500 lm), respectively, as previously described [53 56]. CFTR-inhibitor solution (10 lm CFTRinh172, Sigma) and CFTR-activator solution [10 lm forskolin (FSK) and 30 lm genistein (GST)], both from Sigma were added to the external solution to inhibit or activate CFTR, respectively. For all graphic representations of I V relationships, currents were normalized by membrane capacitances (20 35 pf) to remove variability due to differences in cell sizes. For each patch-clamp analysis, measurements were made on three different cells from four different cultures. Immunofluorescence Cells were seeded on cover slips and incubated at 37 C until % confluence was obtained. After two washes with phosphate-buffered saline (PBS; 1X), cells were fixed in 4 % Paraformaldehyde (PFA) for 10 min. After fixation and three 5 min washes with PBS, cells were permeabilized with 0.2 % Triton X-100 in PBS (PBS-T) for 10 min. After 5 min washes with PBS, non-specific binding was prevented by blocking with 3 % Bovine Serum Albumin (BSA) in 1X PBS (2 h at room temperature). Primary antibody raised against TRPM7 (ab85016, Abcam) was applied overnight at room temperature with 3 % BSA in 1X PBS. After three 5 min washes with PBS-T, cells were incubated with Alexafluor 488 donkey anti-mouse IgG (Santa Cruz) secondary antibody (1:800 in 3 % BSA) for 1 h. Nuclei were labelled with 4 0,6-diamidino-2-phenylindole (DAPI, Sigma; 1:1000 in PBS 1X). Slides were mounted with Fluorsave (Calbiochem) and examined under a Zeiss AxioStar-Plus microscope. Images were collected with a Zeiss 1009 objective. Cell surface biotinylation Cell surface biotinylation was performed as previously described [28]. Cells were grown in serum-free medium overnight. Culture medium was removed and cells were washed three times with PBS (1X plus 0.1 mm CaCl 2 and 1 mm MgCl 2 ) and once with PBS (1X). Cells were then exposed to sulfo-nhs-ss-biotin for 30 min on ice, rinsed twice with BSA quenching buffer (1 % BSA in PBS 1X, ph 7.4) and incubated 10 min in the same buffer. Cells were scraped in PBS (ph 7.4) and centrifuged at 2400 rpm (6 min, 4 C). Cell pellets were suspended in RadioImmunoPrecipitation Assay buffer (RIPA) plus anti-proteases and incubated for 30 min on ice. The resulting lysates were centrifuged at 16,000g for 15 min at 4 C and total cellular protein content was determined using Lowry s method 150
187 Function and regulation of TRPM7, as well as intracellular magnesium content, are altered in [47]. The supernatants were incubated with streptavidin beads overnight at 4 C. After a brief centrifugation, supernatants were removed and the beads were washed four times in RIPA. Biotinylated proteins were eluted from streptavidin beads using 5X sample buffer containing 2-mercaptoethanol to cleave the NHS-SS-biotin. Biotinylated proteins (membrane proteins) and eluated proteins (non biotinylated proteins) were probed to assess TRPM7 expression using western blot experiments (mouse monoclonal anti-trpm7, 1:200, N-17, sc-19562; Santa Cruz Biotechnology, USA). For normalization and to assess the membrane protein enrichment, Na? /K? -ATPase (a1 isoform) expression was assessed on the same blots, probed with a mouse monoclonal antibody (1:200, clone 9-A5, Santa Cruz Biotechnology, USA). For the detection of TRPM7 and Na? /K? -ATPase, the secondary antibody was anti-mouse (1:20,000; abcam ab6820). Experiments were performed twice with two different cultures. STRING query of TRPM7 s interaction network The potential interactions of TRPM7 were evaluated using the online Search Tool for the Retrieval of Interacting Genes/Protein (STRING 9.0, to predict both direct and indirect interactions [57]. The confident scoring of the interactions were based on known and predicted protein interactions collected from direct (physical) and indirect (functional) associations. The score was set at 0.7 to represent a high confidence level. The database quantitatively integrates interaction data from four sources genomic context, high-throughput experiments, coexpression and previous knowledge from research publications. The results are presented with the input protein (in this case TRPM7) in the context of a graphical network of interaction partners. Associations in STRING are provided with a probabilistic confidence score, which represents an estimate of how likely a given association describes a functional linkage between two proteins. The computed score indicates a high confidence when more than one type of information supports the association (direct physical interactions, text mining or experimental). Statistics Results were expressed as mean ± standard error of the mean (SEM). Differences between experimental groups were evaluated by a two-tailed unpaired Student s t test. p \ 0.05 was considered statistically significant (*), p \ was considered very statistically significant (**) and p \ was considered highly statistically significant (***). Results Measurements of [Mg 21 ] i in cells expressing WT-CFTR, DF508-CFTR or G551D-CFTR HeLa cells stably expressing WT-CFTR, DF508-CFTR and G551D-CFTR were previously described regarding CFTR expression and function [45, 57]. [Mg 2? ] i was measured using Mag-fura-2-AM, an intracellular Mg 2? indicator that is ratiometric and UV light-excitable [59 61]. Measurements were performed at 37 C because at room temperature cells did not stick in the wells and because this temperature was physiologically relevant. As shown in Fig. 1, the statistical analysis indicates that [Mg 2? ] i is significantly lower in cells expressing either DF508-CFTR or G551D-CFTR compared to those expressing WT-CFTR. Furthermore, the levels of [Mg 2? ] i in the cells expressing the mutated CFTR were close to that observed in NT Hela cells. [Mg 2? ] i was also significantly lower in cells expressing G551D-CFTR compared to those expressing DF508-CFTR. Detection and quantitation of TRPM7 s mrna and protein Basal TRPM7 mrna expression was assessed in NT, WT- CFTR, DF508-CFTR and G551D-CFTR cells. As shown in Fig. 2a (left panel), a single PCR amplicon (200 bp) was observed indicating that endogenous TRPM7 was relatively abundant in all four cell-lines. No signal was observed in the negative control. A control, G3PDH, was also amplified by PCR (with an amplicon of 121 bp) in the same samples and showed no variation in signal intensity Arbitrary Units 2, ,5 0 *** ** Non transfected Hela cells Wt-CFTR expressing Hela cells F508del-CFTR expressing Hela cells G551D-CFTR expressing Hela cells Fig. 1 Ratiometric measurement of [Mg 2? ] i. [Mg 2? ] i was estimated using Mag-Fura-2-AM in NT cells and in WT-CFTR, DF508-CFTR or G551D-CFTR cells. The bar chart represents the result of the statistical analysis (n = 6) and shows that [Mg 2? ] i is significantly lower in F508del-CFTR and G551D-CFTR experiments compared to comparable levels in cells expressing WT-CFTR or NT cells. Furthermore, [Mg 2? ] i is significantly lower in cells expressing G551D-CFTR compared to those expressing DF508-CFTR. Statistical analysis was performed with results obtained from three different cell cultures
 1,5 1 0,5 Non transfected Hela cells Wt-CFTR expressing Hela cells F508del-CFTR expressing Hela cells G551D-CFTR expressing Hela cells 0 B 1,2 ** * * TRPM7")

188 F. Huguet et al. A bp Ct(target-reference) 1,5 1 0,5 Non transfected Hela cells Wt-CFTR expressing Hela cells F508del-CFTR expressing Hela cells G551D-CFTR expressing Hela cells 0 B 1,2 ** * * TRPM7 (212 kda) Arbitrary Units 1 0,8 0,6 0,4 0,2 Non transfected Hela cells Wt-CFTR expressing Hela cells F508del-CFTR expressing Hela cells G551D-CFTR expressing Hela cells 0 Fig. 2 Expression of TRPM7. a The left image shows (1) the molecular weight marker and an example of representative PCR amplicons for TRPM7 (200 bp) in Hela cells (2) not transfected, or transfected with (3) WT-CFTR, (4) DF508-CFTR or (5) G551D- CFTR. (6) is the negative control. The right bar chart is the quantitative analysis of TRPM7 gene expression, determined by quantitative PCR in each cell line. Data were analyzed by using DCt (target reference) methodologies [28] with no differences observed between cells lines. b The left image shows representative immunodetection experiments of the TRPM7 protein (212 kda) in Hela cells (1) not transfected, or expressing (2) WT-CFTR, (3) DF508-CFTR or (4) G551D-CFTR. Statistical analysis (right) shows that the expression of TRPM7 is increased in both DF508-CFTR and G551D-CFTR experiments compared wildtype assays. Nevertheless, there is less TRPM7 protein in cells expressing G551D-CFTR compared to those expressing DF508-CFTR. Furthermore, TRPM7 expression is decreased in cells expressing WT-CFTR compared to NT cells (n = 6) (not shown). TRPM7 mrna expression was quantified by qpcr and no differences between the cells were observed (Fig. 2a, right panel). Western blots were performed using whole cell lysates to detect the basal expression of TRPM7 protein (212 kda, Fig. 2b, left panel) in the cell-lines. TRPM7 protein was observed in all four cell-lines. Statistical analysis showed some differences between TRPM7 protein levels between the cells, after normalization with an internal control, G3PDH (37 kda), or total protein (Fig. 2b, right panel). The levels of TRPM7 protein were significantly higher in cells expressing DF508-CFTR and G551D-CFTR compared to cells expressing WT-CFTR. No difference was seen between cells expressing mutated CFTR. Lower levels of TRPM7 protein were observed in cells expressing WT- CFTR or G551D-CFTR when compared to NT cells. This difference was not found in cells expressing DF508-CFTR. Cellular localization of TRPM7 Cellular localization of TRPM7 using a specific primary antibody was performed in NT cells and in cells expressing WT-CFTR, DF508-CFTR and G551D-CFTR (Fig. 3). First, negative controls in which the anti-trpm7 antibody 123 was omitted were performed in order to show the specificity of the labelling (Fig. 3a d). Whereas the membrane localization of TRPM7 was likely not observed in NT cells (Fig. 3e g), the protein was observed at the plasma membrane in the cells expressing WT-CFTR (Fig. 3h j) and DF508-CFTR (Fig. 3k m) despite a less strong labelling in these latest cells. As in all cell types, the protein was observed throughout the entire cell with a vesicular pattern in G551D-CFTR cells, with no labelling at the membrane (Fig. 3n p). The membrane localization of TRPM7 was thus increased in WT-CFTR, decreased in DF508-CFTR and absent in membranes of cells expressing G551D- CFTR. Nevertheless, the protein was observed throughout the entire cell with a vesicular pattern in all cell lines. To further confirm our findings regarding the localization of TRPM7, biotinylation experiments were performed (Fig. 4) [28]. Membrane proteins were biotinylated and TRPM7 expression was assessed by western blottings (Fig. 4a). In order to show the enrichment of the membrane proteins within the biotinylated fractions, Na? /K? -ATPase detection was performed. It was observed that the expression of TRPM7 is higher in cells expressing WT-CFTR and DF508-CFTR than in NT cells. The expression was found to be lower in DF508-CFTR than in WT-CFTR cells. The 152

189 Function and regulation of TRPM7, as well as intracellular magnesium content, are altered in A B C C D 10µm 10µm 10µm 10µm E F G 10µm 10µm 10µm H I J 10µm 10µm 10µm K L M 10µm 10µm 10µm N O P 10µm 10µm 10µm Fig. 3 Cellular localization of TRPM7. Immunofluorescence was performed to localize TRPM7 in NT cells (a, e, f, g) and in cells expressing WT-CFTR (b, h, i, j), DF508-CFTR (c, k, l, m) and G551D-CFTR (d, n, o, p). First, negative controls in which first antibody is omitted shows the specificity of the labelling (a, b, c, d). Vesicular labelling in the cytoplasm is observed in all four cell types; however, membrane localization is mostly seen in cells expressing WT-CFTR (i, j) and DF508-CFTR (l, m). This membrane localization is likely not observed in NT cells (f, g). The labelling is less visible in DF508-CFTR (l, m) and lost in G551D-CFTR cells (o, p). Nuclei were DAPI stained (blue, a, b, c, d, e, h, i, j, k, l, m, n). Images were collected with a 1009 objective. Bars represent 10 lm lowest expression of TRPM7 was observed in the membrane of cells expressing G551D-CFTR. Despite no statistical analysis was done because of the low number of experiments (n = 2), bar charts were drawn (Fig. 4b). After normalization by Na? /K? -ATPase expression, the densitometric analysis of the TRPM7 bands showed that the membrane localization of the protein was consistent with the results obtained by immunofluorescence
190 F. Huguet et al. AArbitrary Units B TRPM7 (212 kda) Na + /K + ATPase (100 kda) 1,6 1,4 1,2 1 0,8 0,6 0,4 0,2 0 Bionylated proteins Function and regulation of TRPM7 Eluates Non transfected Hela cells Wt-CFTR expressing Hela cells F508del-CFTR expressing Hela cells G551D-CFTR expressing Hela cells Fig. 4 Detection of TRPM7 within membranes by biotinylation experiments. a Representative immunoblots showing the detection of TRPM7 and Na? /K? -ATPase in NT (1), WT-CFTR (2), DF508- CFTR (3) and G551D-CFTR (4) cells, in biotinylated surface proteins and in non biotinylated proteins (Eluates). TRPM7 is present in the membrane fraction whereas it is likely not observed in non biotinylated proteins. The lower panel showing the presence of a specific membrane protein (Na? /K? -ATPase) in the biotinylated fractions indicates that membrane proteins were enriched in the biotinylated fractions. b Bar chart of the densitometric analysis of the cell surface expression of TRPM7 after normalization by Na? /K? - ATPase. The level of TRPM7 is higher in the membranes of WT- CFTR and DF508-CFTR cells than in NT and G551D-CFTR cells The native function of TRPM7 channels in the plasma membrane was assessed by recording of whole-cell currents in NT cells and in cells expressing WT-CFTR, DF508-CFTR and G551D-CFTR (Fig. 5). Activation of endogenous TRPM7-mediated currents was obtained by a voltage ramp (-100 to?100 mv in 50 ms intervals) and was used to determine current voltage relationships. I V relationships were determined at the time point when current magnitude reached maximum for each cell, with an internal Mg 2? free solution. I V curves were drawn after normalization of the currents by cell capacitances. From the I V curves (Fig. 5a, left panel) it was observed that TRPM7 channel function is drastically reduced in cells expressing mutant CFTR when compared to that of NT cells or to those expressing WT-CFTR. Currents in G551D-CFTR cells were weaker than those measured in cells expressing DF508-CFTR. Statistical analysis was performed and is presented as bar chart (Fig. 5a, right panel). Whereas currents are similar in NT and WT-CFTR cells, they were significantly decreased in cells expressing DF508-CFTR and G551D-CFTR. A difference was observed between cells expressing the mutated CFTR with lower currents in G551D-CFTR cells than in DF508-CFTR 123 cells. TRPM7 is inhibited by Mg 2? [48]. Therefore, inhibition was induced by adding 200 lm Mg 2? in the intracellular solution, and I V curves were recorded and compared to those obtained without divalent ions (Fig. 5b, left panel). It was observed that the presence of Mg 2? inhibited TRMP7 currents in NT cells as well as in cells expressing WT-CFTR. No effect was seen in DF508-CFTR or G551D-CFTR experiments. Statistical analysis (Fig. 5b, right panel) showed that 200 lm Mg 2? inhibited TRPM7 in NT and WT-CFTR cells and had no effect in the cells expressing either DF508-CFTR or G551D-CFTR. The latter results suggested that TRPM7 regulation by intracellular Mg 2? is altered in cells expressing mutant CFTR. Modulation of TRPM7 channel function by Mg-ATP, Naltriben and Carvacrol Outward currents at?80 mv were recorded, and curves were drawn after normalization to cell sizes and plotted against time. Inhibition and activation of TRPM7 were achieved by using 500 lm Carvacrol and 50 lm Naltriben in cells expressing WT-CFTR, respectively. The drugs were applied 100 s after the start of the experiments (Fig. 6a). Activation was performed and observed in cells expressing either DF508- CFTR or G551D-CFTR, as currents were already very low when compared to those observed in WT-CFTR experiments (Fig. 6b, c, respectively). These results indicated that the recorded currents were due to TRPM7, which is consistent with previous reports [53 56]. Nevertheless, due to lower basal currents in DF508-CFTR and G551D-CFTR experiments, the effect of Naltriben was lower when compared to cells expressing WT-CFTR. The measurements presented in Fig. 5 also confirm that the function of TRPM7 is altered when mutant CFTR is expressed in cells. Mg-ATP is requested to measure the Cl - channel activity of CFTR [19, 36] and also inhibits TRPM7 [38]; therefore, the effects of Carvacrol and Natrilben in cells expressing WT-CFTR were assessed in the presence of intracellular Mg-ATP (Fig. 7). The same conditions were used as the previous set of experiments (see Fig. 6), except that 5 mm Mg-ATP was present in the intracellular solution. Figure 7a shows the time course of the currents with or without Mg-ATP in the intracellular buffer. It is observed that Mg-ATP inhibited TRPM7 activity in WT-CFTR experiments as expected. Furthermore, inhibition and activation of TRPM7 by Carvacrol and Natrilben, respectively, were obtained in cells expressing WT-CFTR in the presence of Mg-ATP, (Fig. 7b, c). However, the action of the drugs was lowered by decreased TRPM7 function due to the presence of 5 mm Mg-ATP in the intracellular solution. Figure 8 shows the activation and the inactivation of TRPM7 in cells expressing either DF508-CFTR or G551D-CFTR in the presence of 5 mm Mg-ATP. Figure 8a (upper panel) 154
191 Function and regulation of TRPM7, as well as intracellular magnesium content, are altered in A pa/pf pa/pf *** x -400 NT Hela cells Wt-CFTR expressing Hela cells F508del-CFTR expressing Hela cells G551D-CFTR expressing Hela cells mv NT Hela cells Wt-CFTR expressing Hela cells F508del-CFTR expressing Hela cells G551D-CFTR expressing Hela cells * B pa/pf pa/pf x NT Hela cells; + 200µ M [Mg 2+ ] i Wt-CFTR expressing Hela cells; µ M [Mg2+ ] i F508del-CFTR expressing Hela cells; G551D-CFTR expressing Hela cells; + 200µ M [Mg 2+ ] i + 200µ M [Mg 2+ ] i mv Fig. 5 Function and regulation of TRPM7. Patch-clamp analysis was performed in NT Hela cells and in cells expressing, WT-CFTR, DF508-CFTR or and G551D-CFTR. a The left panel shows example current voltage (I-V) relationships for TRPM7 channels obtained in the four cell lines with Mg 2? -free intracellular solution. I V relationships were taken at the time point when current magnitude reached maximum for each cell. Statistical analysis was performed using the pa/pf values taken at 100 mv and are presented as bar chart (right panel). Whereas currents are similar in NT and WT- CFTR cells, they were significantly decreased in DF508-CFTR and *** *** + [Mg 2+ ] i + [Mg 2+ ] + [Mg 2+ i ] i NT Hela cells Wt-CFTR expressing Hela cells + [Mg 2+ ] i F508del-CFTR expressing Hela cells G551D-CFTR expressing Hela cells G551D-CFTR cells. A difference is also observed between cells expressing the mutated CFTR. The currents are significantly lower when in G551D-CFTR cells than in DF508-CFTR cells. b Comparison of the I V relationships for TRPM7 channels in the four cell lines, with 200 lm Mg 2? and without Mg 2? in the intracellular solution (left panel). Bar chart (right panel) shows that whereas the function of TRPM7 is inhibited by Mg 2? in NT and WT-CFTR cells, no inhibition is observed in the cells expressing either DF508-CFTR or G551D-CFTR shows the time course of DF508-CFTR-induced currents, with or without Mg-ATP in the intracellular buffer, and it was deduced that Mg-ATP inhibited TRPM7 activity in the presence of mutant CFTR. Figure 8a (lower panel) shows that TRPM7 was activated and inactivated in the presence of Naltriben and Carvacrol, respectively, despite the presence of Mg-ATP in the intracellular solution. In Fig. 8b (upper panel), it was observed that TRPM7 currents in cells expressing G551D-CFTR were poorly inhibited by Mg-ATP in the intracellular buffer. Once again, TRPM7 was activated in this mutant cell-line in the presence of Naltriben, despite the presence of Mg-ATP in the intracellular solution (Fig. 8b, lower panel). Inactivation by Carvacrol was lower than in DF508-CFTR cells
192 F. Huguet et al. Fig. 6 Modulation of TRPM7 function in the absence of intracellular Mg 2?. The temporal development of TRPM7 currents were extracted from individual ramp current recordings by measuring current amplitudes at a voltage of?80 mv. a (upper panel) Curve showing time-dependent currents measured in the absence of intracellular Mg 2? (black) and in the presence of Carvacrol (grey) in cells expressing WT-CFTR. The inhibitory effect of Carvacrol is observed. a (lower panel). Curve showing time-dependent currents measured in the absence of intracellular Mg 2? (black) and in the presence of Naltriben (grey) in cells expressing WT-CFTR. Activation of TRPM7 is observed. b Curve showing time-dependent currents measured in the absence of intracellular Mg 2? (black) and in the presence of Naltriben (grey) in cells expressing DF508-CFTR. Activation of TRPM7 is observed. c Curve showing time-dependent currents measured in the absence of intracellular Mg 2? (black) and in the presence of Naltriben (grey) in cells expressing G551D-CFTR. Activation of TRPM7 is observed. The curves presented in a, b and c are the mean curves of three cells from three different cell cultures A pa/pf 600 B C pa/pf 2000 Wt-CFTR expressing Hela cells Carvacrol (500µ M) Naltriben (50µ M) Time (s) Time (s) pa/pf pa/pf Wt-CFTR expressing Hela cells F508del-CFTR expressing Hela cells Naltriben (50µ M) 0 Time (s) G551D-CFTR expressing Hela cells Naltriben (50µ M) Time (s) Measurement of CFTR channel function in the presence of Naltriben CFTR function was measured by patch-clamping in cells expressing WT-CFTR, DF508-CFTR or G551D-CFTR (n = 3 cells from four different cell cultures for each cell 123 type). Activators (FSK and GST) and inhibitor (Inhibitor 172) of CFTR were used to ensure the specificity of the recordings (Fig. 9a). Figure 9b shows the normalized I V curve obtained in WT-CFTR experiments, in the presence of either Naltriben or Carvacrol. It was observed that TRPM7 inhibition had no effect on CFTR function, 156
193 Function and regulation of TRPM7, as well as intracellular magnesium content, are altered in Fig. 7 Modulation of TRPM7 function in WT-CFTR experiments, in the presence of 5 mm Mg-ATP. The same conditions as in Fig. 5 were used except that 5 mm Mg- ATP was present in the intracellular solution. a Time course of currents with (grey) or without (black) Mg-ATP in the intracellular buffer. Mg-ATP inhibited TRPM7 activity in cells expressing WT-CFTR. The inlet shows the initial decrease of currents due to Mg-ATP, whereas in the absence of Mg- ATP the initial currents increase. b Inhibition of TRPM7 by Carvacrol (grey). c Activation of TRPM7 by Natrilben A B pa/pf Wt-CFTR expressing Hela cells 600 0mM MgATP mM MgATP Time (s) pa/pf 50 Wt-CFTR expressing Hela cells, 5mM MgATP 40 Carvacrol (500µ M) Time (s) C Wt-CFTR expressing Hela cells, 5mM MgATP pa/pf Naltriben (50µ M) Time (s) whereas TRPM7 activation by Naltriben led to decreased WT-CFTR channel function (Fig. 9b). A significant decrease in WT-CFTR function (-26 %) was observed in the presence of Naltriben, but no significant effect was detected by the addition of Carvacrol (Fig. 9c). For cells expressing DF508-CFTR (Fig. 10), I V curves which were obtained in the presence of Naltriben and showed a significant decreased function of the mutated protein (Fig. 10a). This decreased function was more important than that of the specific inhibitor 172 (-39.8 %, Fig. 10b). I V curves were also obtained in the presence of Naltriben in cells expressing G551D-CFTR (Fig. 11). I V curves showed that Naltriben activated the function of G551D- CFTR and that this activation was higher than that of the addition of FSK plus GST. Indeed, a significantly increased chloride channel activity of G551D-CFTR (?824.2 %) was observed (Fig. 11b). Measurements of Ca 21 influx through TRPM7 TRPM7 channel conductance was monitored using a FlexStation 3 scanning fluorometer to monitor the Ca 2? - independent (360-nm excitation/510-nm emission) fura- 2 quenching by Mn 2? in cells, as previously described [62]. As shown in Fig. 12a, the curves representing Ca 2? influx which were measured in all the cell types (n = 6 for each cell type), presented some differences among the cell types. Bar charts (Fig. 12b) were drawn using the average slope values of the quenching. Quenching rates were not different in WT-CFTR and NT cells. Ca 2? influx through TRPM7 was lower in cells expressing DF508-CFTR than in WT-CFTR cells. At the opposite, it was increased in the cells expressing G551D-CFTR. Whereas, Waixenicin A had no significant effect upon the quenching rates, Naltriben
194 F. Huguet et al. Fig. 8 Activation of TRPM7 function in cells expressing either DF508-CFTR or G551D- CFTR, in the presence of 5 mm Mg-ATP. The same conditions as in Fig. 5 were used except that 5 mm Mg- ATP was present in the intracellular solution. a (upper panel) Time course of currents in cells expressing F508del- CFTR, with (grey) or without (black) Mg-ATP in the intracellular buffer. Mg-ATP inhibited TRPM7 activity in F508del-CFTR experiments. a (lower panel) TRPM7 is activated and inhibited in the presence of Naltriben or Carvacrol, respectively, despite the presence of Mg-ATP in the intracellular solution, b (upper panel). Time course of currents in cells expressing G551D- CFTR, with (grey) or without (black) Mg-ATP in the intracellular buffer. Mg-ATP failed to inhibit TRPM7 activity in these cells. b (lower panel) TRPM7 is activated and poorly inhibited by Naltriben or Carvacrol, respectively in cells expressing G551D-CFTR, despite the presence of Mg-ATP in the intracellular solution A pa/pf F508del-CFTR expressing Hela cells mM MgATP 80 5mM MgATP Time (s) pa/pf 5mM MgATP + Naltriben (50µ M) mM MgATP + Carvacrol (500µ M) Time (s) 300 B pa/pf G551Dl-CFTR expressing Hela cells 140 0mM MgATP 120 5mM MgATP Time (s) 300 pa/pf 250 5mM MgATP + Naltriben (50µ M) mM MgATP + Carvacrol (500µ M) Time (s) significantly increased the Ca 2? influx in cells expressing DF508-CFTR when compared to non-treated DF508- CFTR cells. The same recordings, using the same conditions as for Hela cells were performed in native HEK293 cells (n = 13). The quenching curves are presented in Fig. 12c (upper panel). The statistical analysis was performed and is presented as bar charts in Fig. 12c (lower panel). In these cells, a significant activation and inhibition of the Ca 2? influx through TRPM7 was obtained by the use of Naltriben (n = 10) and Waixenicin A (n = 6), respectively. This indicates that the absence of modulation observed in the Hela cells expressing WT-CFTR is likely due to the cell types and is not due to technical artifacts
195 Function and regulation of TRPM7, as well as intracellular magnesium content, are altered in Fig. 9 Inhibition of WT-CFTR currents by Naltriben. a I V curves (normalized by cell capacitance pf, mean ± SEM) for CFTR currents in WT-CFTR expressing cells, with 5 mm Mg-ATP in the intracellular buffer. CFTR was activated by FSK and GST, and inhibited by CFTR inhibitor 172, indicating that the recorded currents were due to CFTR. b Curves obtained in the presence of Naltriben and Carvacrol were plotted. Naltriben and Carvacrol activates and inhibites CFTR, respectively. c Bar chart showing the statistical analysis of the currents noted at?80 mv indicating that WT-CFTR function is decreased in the presence of Naltriben A B pa/pf pa/pf mv WT-CFTR plus (FSK + GST) WT-CFTR WT-CFTR plus Inhibitor CFTR CFTR plus Naltriben CFTR plus Carvacrol mv -400 C pa/pf *** -26% CFTR CFTR plus Naltriben CFTR plus Carvacrol Prediction of the protein protein interaction network of TRPM7 The interaction network of TRPM7 was evaluated with the online Search Tool for the Retrieval of Interacting Genes/ Protein to predict both direct and indirect interactions [54]. As shown in Fig. 13, based on bibliographic analysis, putative interacting partners of TRPM7 were found to be annexin A1 (ANXA1, score: 0.925); myosin heavy chain 9 (non-muscle, MYH9, score: 0.910); kininogen 1 (KNG1, score: 0.885); transient receptor potential cation channel 5, subfamily V, member 5 (TRPV5, score: 0.869); transient receptor potential cation channel 6, subfamily VI, member 6 (TRPV6, score: 0.859); myosin heavy chain 10 (non
196 F. Huguet et al. Fig. 10 Inhibition of DF508- CFTR currents by Naltriben. a I V normalized curves for CFTR currents in cells expressing DF508-CFTR (whole-cell configuration with 5 mm Mg-ATP in the intracellular buffer) in the presence or absence of Naltriben. CFTR was activated by FSK and GST and inhibited by CFTR inhibitor 172, indicating that the recorded currents were due to CFTR. The inhibition by Naltriben is more efficient than that of inhibitor 172. b Bar chart showing the statistical analysis of currents (?80 mv) indicating that DF508-CFTR function is decreased by Naltriben A pa/pf mv F508-CFTR + FSK + GST F508-CFTR F508-CFTR + Inhibitor172 F508-CFTR plus Naltriben B ** -39.8% F508-CFTR F508-CFTR plus Naltriben 20 0 muscle, MYH10, score: 0.853); transient receptor potential cation channel, subfamily V, member 1, (VR1, score: 0.834); transient receptor potential cation channel, subfamily V, member 1 (TRPV1, score: 0.832); transient receptor potential cation channel, subfamily V, member 4 (TRPV4, score: 0.832); and phospholipase C (beta 1, PLCB1, score: 0.836). Biochemical evidence was only depicted for annexina1 (ANXA1), myosin heavy chain 9 (MYH9), myosin heavy chain 10 (MYH10) and phospholipase C beta 1 (PLCB1). ANXA1 is a Calcium/ phospholipid-binding protein which promotes membrane fusion and is involved in exocytosis. It also regulates phospholipase A2 activity. MYH9 and MYH10 are nonmuscle cellular myosins that play a role in cytokinesis, cell shape and secretion. PLCB1 which is phosphoinositidespecific, catalyzes the formation of inositol 1,4,5-trisphosphate and diacylglycerol from phosphatidylinositol 4,5- bisphosphate. This reaction uses calcium as a cofactor and plays an important role in the intracellular transduction of many extracellular signals. Discussion Magnesium ions, being one of the most abundant intracellular cations present at a cellular molar concentration of [59], are essential for a wide variety of biochemical reactions and physiological functions. Mg 2? is
197 Function and regulation of TRPM7, as well as intracellular magnesium content, are altered in Fig. 11 Activation of G551D- CFTR currents by Naltriben. a I V normalized curves for CFTR currents in cells expressing G551D-CFTR (with 5 mm Mg-ATP in the intracellular buffer) plus Naltriben. CFTR was activated by FSK and GST, and inhibited by CFTR inhibitor 172, indicating that the recorded currents were due to CFTR. The activation by Naltriben is more efficient than that of FSK plus GST. b Bar chart showing the statistical analysis of currents (?80 mv) indicating that G551D-CFTR function is very significantly increased by Naltriben -80 A pa/pf G551D-CFTR G551D-CFTR + FSK +GST G551D-CFTR +Inhibitor172 G551D-CFTR + Naltriben mv -200 B pa/pf 300 *** % G551D-CFTR G551D-CFTR plus Naltriben especially important for enzymes that require nucleotides (e.g. kinases, G-proteins and ATPases), as the biologically active form of ATP is the Mg-ATP complex. [Mg 2? ] i is known to regulate the expression of CFTR. Knowing that a decrease in [Mg 2? ] i leads to reduced opening and closing rates of the CFTR channel [34], we first estimated the Mg 2? content of cells expressing WT-CFTR, and DF508- CFTR and G551D-CFTR mutants. The DF508 and G551D mutations were chosen because once mutant proteins reach plasma membranes, they exhibit decreased and altered Cl - channel function, independently of their membrane localisation [14 16]. We observed a decreased [Mg 2? ] i in cells expressing DF508-CFTR or G551D-CFTR compared to that of cells expressing WT-CFTR. The effects of changes in the concentration of free Mg 2? on the activity of CFTR Cl - channels were previously investigated and it was shown that Mg 2? facilitates ATP binding that opens the CFTR channel [36]. Indeed, in an excised inside-out membrane patch experiment, removing Mg 2? from the cytosolic solution reduced, but did not abolish, CFTR activity and addition of Mg 2? restored the Cl - currents. Therefore, altered levels of [Mg 2? ] i in cells expressing mutant CFTR could explain, at least in part, the observed altered Cl - channel functions related to some mutations of
198 F. Huguet et al. A Normalized fluorescence (360nm) 0,1 0-0,1-0,2-0,3-0,4-0,5-0,6-0,7-0,8 2mM MnCl Time (s) Wt-CFTR expressing Hela cells; F508-CFTR expressing Hela cells ; G551D-CFTR expressing Hela cells ; Non transfected Hela cells ; plus Naltriben plus Naltriben plus Naltriben plus Naltriben C Normalized fluorescence (360nm) 0,1 0-0,1-0,2-0,3-0,4-0,5-0,6-0,7-0,8 2mM MnCl Time (s) HEK293 plus Naltriben HEK293 plus waixenicin HEK293 0 B Normalized fluorescence (360nm) 0 0,1 0-0,1-0,2-0,3-0,4-0,5-0,6-0,7-0,8 2mM MnCl Time (s) Naltriben Wt-CFTR expressing Hela cells; F508-CFTR expressing Hela cells ; G551D-CFTR expressing Hela cells ; Non transfected Hela cells ; Waixenicin A plus waixenicin A plus waixenicin A plus waixenicin A plus waixenicin A Normalized quenching slope (%) *** ** HEK293 HEK293 plus naltriben HEK293 plus waixenicin A Normalized quenching slope (%) *** Non transfected Hela cells Wt-CFTR expressing Hela cells F508-CFTR expressing Hela cells G551D-CFTR expressing Hela cells ** *** ** * Fig. 12 Measurements of Ca 2? influx through TRPM7. a Example of quenching curves showing the Ca 2? influx which were measured in all the cell types. The upper panel shows a set of curves obtained in the absence and in the presence of TRPM7 activator. The lower panel shows the curves obtained without and in the presence of TRPM7 inhibitor. b Bar charts show the average slope values of the quenching, taken at the 300 s time point. A significant difference was observed among the different cell types. Without any modulator of TRPM7, the quenching rate was significantly higher in G551D-CFTR than in WT-CFTR or NT cells. At the contrary, the quenching rate CFTR. This hypothesis is reinforced by the fact that we saw less Mg 2? in cells expressing G551D-CFTR than in those expressing DF508-CFTR. This decreased [Mg 2? ] i in cells expressing DF508-CFTR and G551D-CFTR can be correlated to the described specific alterations in the channel function mediated by these two mutants [14 17, 36, 45, 58]. It was also observed that [Mg 2? ] i is higher in cells expressing WT-CFTR compared to NT cells. This latter point led us to postulate that CFTR itself is likely involved in the regulation of intracellular Mg 2?, as it does for Ca 2? regulation [32]. Nevertheless, this hypothesis requires further evidence for its validation. 123 was lower in cells expressing DF508-CFTR than in WT-CFTR, G551D-CFTR and NT cells. Whereas, the TRPM7 inhibitor Waixenicin A had no significant effect upon the quenching rates, Naltriben significantly increased the quenching rate in cells expressing DF508- CFTR. c Ca 2? influx through TRPM7 was measured in HEK293 cells. The upper panel shows an example of quenching curves obtained without and with modulation of TRPM7. The lower panel shows the corresponding bar charts indicating that Naltriben and Waixenicin A significantly activates and inhibits TRPM7, respectively [Mg 2? ] i is finely regulated by precise mechanisms that balance Mg 2? uptake, intracellular Mg 2? storage and Mg 2? efflux. Whereas extremely slow turn-over of Mg 2? across the cell plasma membrane under quiescent conditions is described, some efforts have been made to identify and characterize the complex network of Mg 2? -specific transporters (for review [61]). The main vertebrate Mg 2? cellular transporters identified are Ancient Conserved Domain Protein (Acdp2) [63]; Magnesium Transporter 1 (MagT1) [64]; Mitochondrial RNA splicing 2 protein (Mrs2) [65]; Claudin 16 [66]; SoLute Carrier family (SLC41A1 and A2) [67, 68]; TRPM6 [69]; and TRPM7 162
199 Function and regulation of TRPM7, as well as intracellular magnesium content, are altered in Fig. 13 Network of predicted interactions for TRPM7 obtained by STRING. Blue connecting lines indicate that the interaction/s are mentioned in PubMed abstracts. Interactions with Annexin A1 (ANXA1), myosin heavy chain 9 non-muscle (MYH9), myosin heavy chain 10 non-muscle (MYH10) and phospholipase C beta 1 (PLCB1) were given due to experimental data. Scores were set above 0.7 for high confidence [38, 42]. We focused our study on the ubiquitous TRPM7 chanzyme, which is presented as the main [Mg 2? ] i regulator of [41]. The quantitative mrna analysis of TRPM7 showed that there was no difference in the presence or absence of CFTR within cells or when mutant CFTR is expressed. Therefore, the gene expression of TRPM7 itself is unlikely involved in the [Mg 2? ] i differences that were observed. The quantitation of TRPM7 protein levels in the cells was surprising when it was correlated to the measured [Mg 2? ] i. Indeed, low [Mg 2? ] i in cells expressing either DF508-CFTR or G551D-CFTR corresponded to high levels of TRPM7 protein expression. By contrast, high [Mg 2? ] i, which was observed in WT-CFTR experiments, correlated well to a low amount of protein. Thus, we hypothesized that the cellular localization, function, and regulation of the TRPM7 channel could be involved in the decreased [Mg 2? ] i in the cells expressing mutant CFTR. Immunofluorescence and biotinylation experiments showed a vesicular labelling in the cytoplasm in all three cell types, with membrane expression being mostly seen in WT-CFTR experiments. Membrane localization was lower in cells expressing DF508-CFTR and likely absent when G551D-CFTR was expressed. These results are consistent with previously described studies [37, 41, 70, 71]. Furthermore, it was reported that TRPM7 can be translocated within cells in response to external stimulation. For example, TRPM7 accumulates within cells in response to an increased fluid flow and is localized at the plasma membrane in physiological condition, in \100 s [70]. In our experiments, the cellular modifications due to the CFTR mutations could explain the altered membrane localization of TRPM7. Furthermore, the altered membrane localization of TRPM7 in DF508-CFTR and mostly in G551D-CFTR cells could fully explain the decreased [Mg 2? ] i in these cells. Although the decreased membrane localization of TRPM7 in cells expressing either DF508-CFTR or G551D- CFTR is sufficient to explain decreased [Mg 2? ] i, we studied its channel function. We observed that TRPM7 channel activity was decreased and almost abolished in DF508-CFTR and G551D-CFTR experiments, respectively. TRPM7 regulation by increasing Mg 2? was also evaluated according to previous studies [41 44] and we found that it did not respond in cells containing mutant CFTR. As in our cell model, amyotrophic lateral sclerosis and Parkinson s disease are epidemiologically linked to a severe deficiency in Mg 2? [72]. In these disorders, a TRPM7 variant was found in some patients that produces a protein with a missense mutation (T1482I). Further
200 F. Huguet et al. analysis indicated that a recombinant T1482I-TRPM7 mutant exhibited the same kinase catalytic activity as wildtype TRPM7, with an increased sensitivity to inhibition by intracellular Mg 2? [69]. Therefore, in our study the kinase activity of the alpha-kinase domain fused to the ion channel moiety of TRPM7 (for review, [44]) is unlikely to be involved in altering channel function and regulation. The functional interaction between the TRPM7 channel and kinase activity is poorly understood, although it has been shown that TRPM7 kinase activity is not essential for channel activation [39]. Nevertheless, it is also reported that the kinase activity of TRPM7 plays a role in modulating channel activity and that it is itself affected by Mg 2? availability [41, 42, 44]. Thus, we propose that a vicious circle occurs in cells expressing mutant CFTR in which altered TRPM7 function leads to a decrease in [Mg 2? ] i, which in turn modifies the regulation of TRPM7 channel activity. Mg 2? coordinates the c phosphate of ATP and is required for hydrolysis by most enzymes [73]. Indeed, chelating Mg 2? abolishes ATP hydrolysis [32], which is necessary for CFTR channel function [19, 35]. Therefore, we hypothesized that the altered activity of TRPM7 presented here is linked to modifications in the function of DF508-CFTR and G551D-CFTR and we speculated that restoring TRPM7 function could play a positive role in the activity of CFTR. To answer this question, we performed experiments using Naltriben to activate TRPM7 [53, 54]. Whereas Mg-ATP inhibits TRPM7 [38], it has to be used in patch-clamp experiments to measure the function of CFTR [19, 36]. We performed these experiments in the presence of 5 mm Mg-ATP. This experimental condition permitting the electrophysiological recording of CFTR showed that TRPM7 was still active, despite a decreased function. CFTR Cl - channel function was thus recorded in the presence of a TRPM7 activator, according to previously described whole-cell methods [45, 58]. Naltriben led to a decrease in Cl - channel activity for WT-CFTR and DF508- CFTR. By contrast, we observed that TRPM7 activation leads to an increased G551D-CFTR channel function. CFTR is unique amongst ion channels as it is dependent not only on PKA-mediated phosphorylation, but also on ATP binding and hydrolysis [74, 75] by the nucleotidebinding domains (NBDs) which require Mg-ATP [35]. Mg- ATP was found to increase channel activity with a saturating effect above 1 mmol/l [76]; however, it was observed that whole-cell CFTR Cl - currents (activated by FSK) were diminished by increasing the cytosolic free Mg 2? level, even when maintaining Mg-ATP in the cytosol above millimolar concentrations [77]. This could explain why activation of TRPM7 leads to decreased CFTR function in cells expressing WT-CFTR and DF508-CFTR. In G551D-CFTR experiments, [Mg 2? ] i is much lower 123 compared to cells expressing either WT-CFTR or DF508- CFTR and TRPM7 is poorly present in membranes were it exhibits very weak channel activity. Therefore, it can be postulated that the activation of TRPM7 by Naltriben leads to increased [Mg 2? ] i that remains below the inhibitory level for CFTR. Therefore, we show here for the first time that Naltriben is a new potentiator of the G551D-CFTR mutant protein. The presented negative effect of Naltriben on the function of DF508-CFTR was also reported for other potentiators such as VX-770 (Ivacaftor, [78]). Indeed, whereas VX-770 treatments rescue G551D CFTR function, it inhibits functional rescue of DF508 CFTR and reduces the membrane stability of WT-CFTR. The role of TRPM7 upon CFTR function is unlikely due to [Mg 2? ] i modulation alone, but may also involve protein protein interactions and/or the serine/threonine protein kinase domain at the distal C-terminus. Indeed, the C-terminus of TRPM7 contains a TRP box (*25 highly conserved residues) that may interact with phosphatidylinositol 4,5-bisphosphate (PIP 2, for review [79]), which activates CFTR in the complete absence of Mg 2? [80]. Knowing that the kinase domain of TRPM7 also shows some similarity to camp-dependent PKA which also activates CFTR [18, 19], TRPM7 may act on CFTR in many ways involving a multitude of protein interactions. Amongst the proteins predicted to interact with TRPM7, biochemical evidence was only identified for ANXA1, MYH9, MYH10 and PLCB1. MYH and PLCB1 were shown to be involved in CFTR localization and/or function [81, 82]. Thus, it is clear that further experiments are required to ascertain whether altered TRPM7 function and regulation acts upon mutant CFTR or whether the converse interaction occurs. Beside protein protein interactions, Ca 2? is more likely involved in the altered function and regulation of TRPM7 in CF cells, for at least three reasons. First, it was previously described that TRPM7 channels are inhibited by intracellular Ca 2? [37 39, 44, 71, 83]. Second, a larger mobilization of Ca 2? ions in CF bronchial epithelial cells is well described [29 33, 84]. Third, the implication of other TRP channels (TRPV5 and TRPV6) [82] in the increase of Ca 2? influx in CF bronchial epithelial cells were studied and it was shown that this increase is mostly due to an upregulation of TRPV6 activity. Therefore, we propose that the altered function and regulation of TRPM7 presented here are due to an increase in Ca 2? influx in DF508-CFTR cells. This is reinforced by our findings where decreased [Mg 2? ] i in cells expressing mutant CFTR is not involved in the altered function of TRPM7. In order to assess our hypothesis, we measured Ca 2? influx via TRPM7 in cells expressing WT-CFTR, DF508-CFTR and G551D-CFTR. The Ca 2? conductance of TRPM7 has been previously measured using fluorescent cation binding dyes [43], 164
201 Function and regulation of TRPM7, as well as intracellular magnesium content, are altered in commonly using the ratiometric properties of the Ca 2? binding dye fura-2 [61]. Based on its TRPM7 permeability and high fura-2 binding affinity, we used Mn 2? as the quenching reagent in our assay. Measuring TRPM7-mediated Mn 2? entry, rather than Ca 2?, affords several advantages. First, some competing cations entry pathways [e.g., calcium-release activated calcium (CRAC) channels] are less permeable than TRPM7 to Mn 2?. Second, CRAC channel currents can be further disconnected by avoiding Ca 2? -deficient assay conditions, which are needed for optimal measurement of Ca 2? influx. Third, it allowed us to realize our experiment in the presence of physiological levels of Ca 2? and Mg 2? [43]. Because Waixenicin A (a xenicane diterpenoid from the Hawaiian soft coral S. edmondsoni) [85] inhibits TRPM7-mediated Mn 2? quench in a dose-dependent manner and because its inhibitory effect is strongly dependent on the intracellular Mg 2? concentration [86], it was used, due to the observed variations in [Mg 2? ] i among our cell types. We observed a decreased and increased Ca 2? influx through TRPM7 in DF508-CFTR and G551D-CFTR, respectively. Regarding the cells expressing DF508-CFTR, this is in accordance with our results showing a decreased function of TRPM7 in these cells. Regarding the cells expressing G551D-CFTR, we found an increased Ca 2? influx via TRPM7 which was not in accordance with a decreased function of the channel. Therefore, we hypothesize that in this cell type the behaviour of TRPM7 may vary among ions. This point has to be further studied. We also noticed that the drug modulation of TRPM7 by Naltriben and Waixenicin A was difficult to detect in Hela cells. In order to show that this was not due to our experimental conditions, we performed the same experiments in HEK293 cells which have a high level of TRPM7 expression [38]. In native HEK293 cells, a modulation of TRPM7 was observed. This led us to conclude that the TRPM7 modulation was altered in Hela cells expressing a mutated CFTR and that the involvement of TRPM7 in constitutive Ca 2? influx is different among cell types. This cell-dependent contribution in constitutive Ca 2? currents due to TRPM7 was previously suggested [87]. Indeed, it was reported that it can be difficult to detect Ca 2? influx mediated by TRPM7 in some cells because its contribution can be small, relative to the large variety of constitutive currents [43, 88]. Our conclusion is that TRPM7 is likely not involved in the large Ca 2? influx described in cells expressing DF508-CFTR and that the described [Ca 2? ] i overload in these cells is likely responsible for the altered function of TRPM7. Second, we show here for the first time that Ca 2? influx via TRPM7 in cells expressing G551D-CFTR are increased. We show here, for the first time, that the expression and function of TRPM7 are altered in cells expressing either DF508-CFTR or G551D-CFTR (Fig. 14). We also propose CFTR TRPM7 [Mg 2+ ] i Membrane localizaon Funcon Total protein expression Funcon Regulaon Membrane localizaon Acvaon by Naltriben Fig. 14 Summary of results. The table presents previously published information regarding CFTR (grey) and the [Mg 2? ] i and TRPM7 (black) results of this study in order to provide a synthetic view of the correlation between CFTR modulation and our findings that there is a functional link between TRPM7 and CFTR via decreased [Mg 2? ] i and increased Ca 2? influx in CF cells. Hypomagnesaemia has been reported as a common feature in CF patients [89]. Mean serum magnesium range in CF patients was shown to be mmol/l whereas the normal range is [89]. This could explain the low levels of [Mg 2? ] i that we observed in cells expressing the mutated CFTR. Patients who receive oral magnesium present better clinical symptoms [90]. If we hypothesize that Mg 2? supplementation induces a decreased function of TRPM7, this could be in opposition with our results showing that its activation by Naltriben has a positive effect on the function of G551D-CFTR. Nevertheless, the mechanism of action of Mg 2? in pulmonology is not fully understood due to a small correlation between serum and intracellular concentrations [90]. For example, [Mg 2? ] i in erythrocytes, mononuclear cells and granulocytes do not reflect the body s Mg 2? store because it is primarily concentrated in muscles and bones [91]. Therefore, we propose that the effect of supplementation may be different among the type of mutation of CFTR, which is linked to variations in the localization and function of TRPM7. Our results


202 F. Huguet et al. F508del-CFTR cells Wt-CFTR cells Cl Mg 2+ Mg Mg 2+ TRPM7 acvaon Cl - Cl - Cl - CFTR TRPM7 [Mg 2+ ] i - Cl - TRPM7 acvaon X X X - - Mg 2+ +/- +/- +/- Mg 2+ +/- +/- X G551D-CFTR cells Mg 2+ Mg 2+ - X Cl - X TRPM7 acvaon Fig. 15 Diagram presenting the results and the effect of Naltriben. The three cells represent WT-CFTR, DF508-CFTR and G551D- CFTR experiments with their status related to [Mg 2? ] i, CFTR and TRPM7. The effect of Naltriben is shown by arrows which indicate that the drug modifies TRPM7 activity and subsequently CFTR function together with the improvement of the symptoms in CF by Mg 2? supplementation also reinforce the poor correlation between serum and intracellular concentrations of Mg 2?. From a pharmacological point of view, we show here that Naltriben modifies the phenotype of CFTR-expressing cells (Fig. 15). Interestingly, the activation of TRPM7 leads to increased G551D-CFTR function; thus, we propose that Naltriben is a new potentiator for this particular mutation. Acknowledgments We thank the association Gaëtan Salaün and the French foundation Vaincre la Mucoviscidose for funding the present work. We also thank Dr Mohamed Keir (Nanion Technology, Germany) for his help. We acknowledge the grant that funds the ability to provide Waixenicin A: U.S. National Institute of Health Grant P20GM References 1. Kerem BS, Rommens JM, Buchanan JA, Markiewicz D, Cox T, Chakravarti A, Buchwald M, Tsui LC (1989) Identification of the cystic fibrosis gene: genetic analysis. Science 245: Riordan JR, Rommens JM, Kerem BS, Alon N, Rozmahel R, Grzelczak Z, Zielinski J, Lok S, Plavsik N, Chou JL, Drumm ML, Ianuzzi MC, Collins FS, Tsui LC (1989) Identification of the cystic fibrosis gene: cloning and characterization of complementary DNA. Science 245: Zielinski J, Rozmahel R, Bozon D, Kerem BS, Grzelczak Z, Riordan JR, Rommens J, Tsui LC (1991) Genomic DNA sequence of the cystic fibrosis transmembrane conductance regulator (CFTR) gene. Genomics 10: Tabcharani JA, Linsdell P, Hanrahan JW (1997) Halide permeation in wild-type and mutant CFTR chloride channels. J Gen Physiol 110: Linsdell P, Tabcharani JA, Rommens JM, Hou YX, Chang XB, Tsui LC, Riordan JR, Hanrahan JW (1997) Permeability of wildtype and mutant CFTR chloride channels to polyatomic anions. J Gen Physiol 110: Hasegawa H, Skach W, Baker O, Calayag MC, Lingappa V, Verkman AS (1992) A multifunctional aqueous channel formed by CFTR. Science 258: Ballard ST, Spadafora D (2007) Fluid secretion by submucosal glands of the tracheobronchial airways. Respir Physiol Neurobiol 159: Riordan JR (2008) CFTR function and prospects for therapy. Annu Rev Biochem 77: Rosenstein BJ, Zeitlin PL (1998) Cystic fibrosis. Lancet 351: Ratjen F, Döring G (2003) Cystic fibrosis. Lancet 361: Amaral MD, Farinha CM (2013) Rescuing mutant CFTR: a multi-task approach to a better outcome in treating cystic fibrosis. Curr Pharm Des 19:
Dr E. CHEVRET UE2.1 2013-2014. Aperçu général sur l architecture et les fonctions cellulaires
 Aperçu général sur l architecture et les fonctions cellulaires I. Introduction II. Les microscopes 1. Le microscope optique 2. Le microscope à fluorescence 3. Le microscope confocal 4. Le microscope électronique
Aperçu général sur l architecture et les fonctions cellulaires I. Introduction II. Les microscopes 1. Le microscope optique 2. Le microscope à fluorescence 3. Le microscope confocal 4. Le microscope électronique
Les cytokines et leurs récepteurs. Laurence Guglielmi laurence.guglielmi@univ-montp1.frli
 Les cytokines et leurs récepteurs Laurence Guglielmi laurence.guglielmi@univ-montp1.frli l i@ i 1 Les cytokines et leurs récepteurs 2 mécanismes principaux d interactions cellulaires : - contact membranaire
Les cytokines et leurs récepteurs Laurence Guglielmi laurence.guglielmi@univ-montp1.frli l i@ i 1 Les cytokines et leurs récepteurs 2 mécanismes principaux d interactions cellulaires : - contact membranaire
ULBI 101 Biologie Cellulaire L1. Le Système Membranaire Interne
 ULBI 101 Biologie Cellulaire L1 Le Système Membranaire Interne De la nécessité d un SMI Le volume augmente comme le cube de la dimension linéaire, alors que la surface n'est augmentée que du carré Une
ULBI 101 Biologie Cellulaire L1 Le Système Membranaire Interne De la nécessité d un SMI Le volume augmente comme le cube de la dimension linéaire, alors que la surface n'est augmentée que du carré Une
CHAPITRE 3 LA SYNTHESE DES PROTEINES
 CHAITRE 3 LA SYNTHESE DES ROTEINES On sait qu un gène détient dans sa séquence nucléotidique, l information permettant la synthèse d un polypeptide. Ce dernier caractérisé par sa séquence d acides aminés
CHAITRE 3 LA SYNTHESE DES ROTEINES On sait qu un gène détient dans sa séquence nucléotidique, l information permettant la synthèse d un polypeptide. Ce dernier caractérisé par sa séquence d acides aminés
TD de Biochimie 4 : Coloration.
 TD de Biochimie 4 : Coloration. Synthèse de l expérience 2 Les questions posées durant l expérience 2 Exposé sur les méthodes de coloration des molécules : Générique Spécifique Autres Questions Pourquoi
TD de Biochimie 4 : Coloration. Synthèse de l expérience 2 Les questions posées durant l expérience 2 Exposé sur les méthodes de coloration des molécules : Générique Spécifique Autres Questions Pourquoi
Le rôle de l endocytose dans les processus pathologiques
 UE7 Cours n 9 C. LAMAZE 24.11.11 Elise GODEAU (partie1) Guillaume MERGENTHALER (partie2) Le rôle de l endocytose dans les processus pathologiques SOMMAIRE : I. L endocytose à récepteurs : la voie des clathrines
UE7 Cours n 9 C. LAMAZE 24.11.11 Elise GODEAU (partie1) Guillaume MERGENTHALER (partie2) Le rôle de l endocytose dans les processus pathologiques SOMMAIRE : I. L endocytose à récepteurs : la voie des clathrines
Bases de données des mutations
 Bases de données des mutations CFMDB CFTR2 CFTR-France / Registre Corinne THEZE, Corinne BAREIL Laboratoire de génétique moléculaire Montpellier Atelier Muco, Lille, 25-27 septembre 2014 Accès libre http://www.genet.sickkids.on.ca/app
Bases de données des mutations CFMDB CFTR2 CFTR-France / Registre Corinne THEZE, Corinne BAREIL Laboratoire de génétique moléculaire Montpellier Atelier Muco, Lille, 25-27 septembre 2014 Accès libre http://www.genet.sickkids.on.ca/app
La voix en images : comment l évaluation objectivée par logiciel permet d optimiser la prise en charge vocale
 La voix en images : comment l évaluation objectivée par logiciel permet d optimiser la prise en charge vocale Stéphanie Perriere To cite this version: Stéphanie Perriere. La voix en images : comment l
La voix en images : comment l évaluation objectivée par logiciel permet d optimiser la prise en charge vocale Stéphanie Perriere To cite this version: Stéphanie Perriere. La voix en images : comment l
Fonctions non ventilatoires
 UE3-2 - Physiologie Physiologie Respiratoire Chapitre 11 : Fonctions non ventilatoires Docteur Sandrine LAUNOIS-ROLLINAT Année universitaire 2010/2011 Université Joseph Fourier de Grenoble - Tous droits
UE3-2 - Physiologie Physiologie Respiratoire Chapitre 11 : Fonctions non ventilatoires Docteur Sandrine LAUNOIS-ROLLINAT Année universitaire 2010/2011 Université Joseph Fourier de Grenoble - Tous droits
Système de diffusion d information pour encourager les PME-PMI à améliorer leurs performances environnementales
 Système de diffusion d information pour encourager les PME-PMI à améliorer leurs performances environnementales Natacha Gondran To cite this version: Natacha Gondran. Système de diffusion d information
Système de diffusion d information pour encourager les PME-PMI à améliorer leurs performances environnementales Natacha Gondran To cite this version: Natacha Gondran. Système de diffusion d information
Annales de Biologie Cellulaire QCM (niveau SVT 1 er année)
") Annales de Biologie Cellulaire QCM (niveau SVT 1 er année) Equipe pédagogique Université Bordeaux-1 Didier Morin, Michel Moenner, Sophie North, Gérard Tramu et IJsbrand Kramer Contact : i.kramer@iecb.u-bordeaux.fr
Annales de Biologie Cellulaire QCM (niveau SVT 1 er année) Equipe pédagogique Université Bordeaux-1 Didier Morin, Michel Moenner, Sophie North, Gérard Tramu et IJsbrand Kramer Contact : i.kramer@iecb.u-bordeaux.fr
Cellules procaryotes Service histologie Pr.k.mebarek
 Cellules procaryotes Service histologie Pr.k.mebarek I) Les cellules procaryotes II) Les cellules eucaryotes o 1) Caractéristiques générales des cellules eucaryotes o 2) Organisation des cellules eucaryotes
Cellules procaryotes Service histologie Pr.k.mebarek I) Les cellules procaryotes II) Les cellules eucaryotes o 1) Caractéristiques générales des cellules eucaryotes o 2) Organisation des cellules eucaryotes
PARTIE II : RISQUE INFECTIEUX ET PROTECTION DE L ORGANISME. Chapitre 1 : L Homme confronté aux microbes de son environnement
 PARTIE II : RISQUE INFECTIEUX ET PROTECTION DE L ORGANISME Chapitre 1 : L Homme confronté aux microbes de son environnement I- Les microbes dans notre environnement Qu est-ce qu un microbe? Où se trouvent-ils?
PARTIE II : RISQUE INFECTIEUX ET PROTECTION DE L ORGANISME Chapitre 1 : L Homme confronté aux microbes de son environnement I- Les microbes dans notre environnement Qu est-ce qu un microbe? Où se trouvent-ils?
Hépatite chronique B Moyens thérapeutiques
 Hépatite chronique B Moyens thérapeutiques Dr Olfa BAHRI Laboratoire de Virologie Clinique Institut Pasteur de Tunis INTRODUCTION Plus de 300. 10 6 porteurs chroniques de VHB dans le monde Hépatite chronique
Hépatite chronique B Moyens thérapeutiques Dr Olfa BAHRI Laboratoire de Virologie Clinique Institut Pasteur de Tunis INTRODUCTION Plus de 300. 10 6 porteurs chroniques de VHB dans le monde Hépatite chronique
IMMUNOLOGIE. La spécificité des immunoglobulines et des récepteurs T. Informations scientifiques
 IMMUNOLOGIE La spécificité des immunoglobulines et des récepteurs T Informations scientifiques L infection par le VIH entraîne des réactions immunitaires de l organisme qui se traduisent par la production
IMMUNOLOGIE La spécificité des immunoglobulines et des récepteurs T Informations scientifiques L infection par le VIH entraîne des réactions immunitaires de l organisme qui se traduisent par la production
PLAN. Intérêt des cellules souches exogènes (hématopoïétiques ou mésenchymateuses) dans la réparation/régénération
 dans la réparation/régénération") Cellules souches & appareil respiratoire : Perpectives thérapeutiques Pr Carole Planès Physiologie SMBH Bobigny EA 2363, UFR SMBH, Université Paris 13 carole.planes@avc.aphp.fr Master 2 B2PCR Respiration
Cellules souches & appareil respiratoire : Perpectives thérapeutiques Pr Carole Planès Physiologie SMBH Bobigny EA 2363, UFR SMBH, Université Paris 13 carole.planes@avc.aphp.fr Master 2 B2PCR Respiration
ÉCOLES NORMALES SUPÉRIEURES ÉCOLE NATIONALE DES PONTS ET CHAUSSÉES CONCOURS D ADMISSION SESSION 2013 FILIÈRE BCPST COMPOSITION DE BIOLOGIE
 ÉCOLES NORMALES SUPÉRIEURES ÉCOLE NATIONALE DES PONTS ET CHAUSSÉES CONCOURS D ADMISSION SESSION 2013 FILIÈRE BCPST COMPOSITION DE BIOLOGIE Épreuve commune aux ENS de Cachan, Lyon, Paris et de l ENPC Durée
ÉCOLES NORMALES SUPÉRIEURES ÉCOLE NATIONALE DES PONTS ET CHAUSSÉES CONCOURS D ADMISSION SESSION 2013 FILIÈRE BCPST COMPOSITION DE BIOLOGIE Épreuve commune aux ENS de Cachan, Lyon, Paris et de l ENPC Durée
Insulinothérapie et diabète de type 1
 Insulinothérapie et diabète de type 1 Introduction: la molécule d insuline L instauration de l insulinothérapie Dispositif d administration de l insuline Les propriétés de l insuline Insuline et schémas
Insulinothérapie et diabète de type 1 Introduction: la molécule d insuline L instauration de l insulinothérapie Dispositif d administration de l insuline Les propriétés de l insuline Insuline et schémas
Tableau récapitulatif : composition nutritionnelle de la spiruline
 Tableau récapitulatif : composition nutritionnelle de la spiruline (Valeur énergétique : 38 kcal/10 g) Composition nutritionnelle Composition pour 10 g Rôle Protéines (végétales) 55 à 70 % Construction
Tableau récapitulatif : composition nutritionnelle de la spiruline (Valeur énergétique : 38 kcal/10 g) Composition nutritionnelle Composition pour 10 g Rôle Protéines (végétales) 55 à 70 % Construction
Les différents types de cancers et leurs stades. Dr Richard V Oncologie MédicaleM RHMS Baudour et Erasme La Hulpe 1/12/07
 Les différents types de cancers et leurs stades Dr Richard V Oncologie MédicaleM RHMS Baudour et Erasme La Hulpe 1/12/07 CARCINOGENESE multiple steps, accumulation d altd altérations continuum lésionnel
Les différents types de cancers et leurs stades Dr Richard V Oncologie MédicaleM RHMS Baudour et Erasme La Hulpe 1/12/07 CARCINOGENESE multiple steps, accumulation d altd altérations continuum lésionnel
La filtration glomérulaire et sa régulation
 UE3-2 - Physiologie rénale Chapitre 4 : La filtration glomérulaire et sa régulation Professeur Diane GODIN-RIBUOT Année universitaire 2010/2011 Université Joseph Fourier de Grenoble - Tous droits réservés.
UE3-2 - Physiologie rénale Chapitre 4 : La filtration glomérulaire et sa régulation Professeur Diane GODIN-RIBUOT Année universitaire 2010/2011 Université Joseph Fourier de Grenoble - Tous droits réservés.
statique J. Bertrand To cite this version: HAL Id: jpa-00237017 https://hal.archives-ouvertes.fr/jpa-00237017
 Quelques théorèmes généraux relatifs à l électricité statique J. Bertrand To cite this version: J. Bertrand. Quelques théorèmes généraux relatifs à l électricité statique. J. Phys. Theor. Appl., 1874,
Quelques théorèmes généraux relatifs à l électricité statique J. Bertrand To cite this version: J. Bertrand. Quelques théorèmes généraux relatifs à l électricité statique. J. Phys. Theor. Appl., 1874,
Chapitre 2 : Respiration, santé et environnement.
 Chapitre 2 : Respiration, santé et environnement. Rappels : L air qui nous entoure contient 3 gaz principaux: 71% d azote, 21% d oxygène, 0,03 % de CO2 et quelques gaz rares. L eau de chaux se trouble
Chapitre 2 : Respiration, santé et environnement. Rappels : L air qui nous entoure contient 3 gaz principaux: 71% d azote, 21% d oxygène, 0,03 % de CO2 et quelques gaz rares. L eau de chaux se trouble
CONCOURS DE L INTERNAT EN PHARMACIE
 Ministère de l enseignement supérieur et de la recherche Ministère de la santé et des sports CONCOURS DE L INTERNAT EN PHARMACIE Valeurs biologiques usuelles Edition de Novembre 2009 (6 pages) Conseil
Ministère de l enseignement supérieur et de la recherche Ministère de la santé et des sports CONCOURS DE L INTERNAT EN PHARMACIE Valeurs biologiques usuelles Edition de Novembre 2009 (6 pages) Conseil
Les OGM. 5 décembre 2008. Nicole Mounier
 Les OGM 5 décembre 2008 Nicole Mounier Université Claude Bernard Lyon 1 CGMC, bâtiment Gregor Mendel 43, boulevard du 11 Novembre 1918 69622 Villeurbanne Cedex OGM Organismes Génétiquement Modifiés Transfert
Les OGM 5 décembre 2008 Nicole Mounier Université Claude Bernard Lyon 1 CGMC, bâtiment Gregor Mendel 43, boulevard du 11 Novembre 1918 69622 Villeurbanne Cedex OGM Organismes Génétiquement Modifiés Transfert
Transport des gaz dans le sang
 UE3-2 - Physiologie Physiologie Respiratoire Chapitre 9 : Transport des gaz dans le sang Docteur Sandrine LAUNOIS-ROLLINAT Année universitaire 2010/2011 Université Joseph Fourier de Grenoble - Tous droits
UE3-2 - Physiologie Physiologie Respiratoire Chapitre 9 : Transport des gaz dans le sang Docteur Sandrine LAUNOIS-ROLLINAT Année universitaire 2010/2011 Université Joseph Fourier de Grenoble - Tous droits
La fonction respiratoire
 La fonction respiratoire Introduction Fonction vitale : Au-delà de 3 min = atteintes graves Après 5 min = très altérés On parle de 3 phénomènes : Ventilation : respiration visible, échanges aériens intérieur
La fonction respiratoire Introduction Fonction vitale : Au-delà de 3 min = atteintes graves Après 5 min = très altérés On parle de 3 phénomènes : Ventilation : respiration visible, échanges aériens intérieur
Transport des gaz dans le sang
 UE3-2 - Physiologie Physiologie Respiratoire Chapitre 9 : Transport des gaz dans le sang Docteur Sandrine LAUNOIS-ROLLINAT Année universitaire 2011/2012 Université Joseph Fourier de Grenoble - Tous droits
UE3-2 - Physiologie Physiologie Respiratoire Chapitre 9 : Transport des gaz dans le sang Docteur Sandrine LAUNOIS-ROLLINAT Année universitaire 2011/2012 Université Joseph Fourier de Grenoble - Tous droits
Séquence 2. L expression du patrimoine génétique. Sommaire
 Séquence 2 L expression du patrimoine génétique Sommaire 1. La synthèse des protéines 2. Phénotypes, génotypes et environnement Synthèse de la séquence 2 Exercices de la séquence 2 Glossaire des séquences
Séquence 2 L expression du patrimoine génétique Sommaire 1. La synthèse des protéines 2. Phénotypes, génotypes et environnement Synthèse de la séquence 2 Exercices de la séquence 2 Glossaire des séquences
Chapitre 7 : Structure de la cellule Le noyau cellulaire
 UE2 : Structure générale de la cellule Chapitre 7 : Structure de la cellule Le noyau cellulaire Professeur Michel SEVE Année universitaire 2010/2011 Université Joseph Fourier de Grenoble - Tous droits
UE2 : Structure générale de la cellule Chapitre 7 : Structure de la cellule Le noyau cellulaire Professeur Michel SEVE Année universitaire 2010/2011 Université Joseph Fourier de Grenoble - Tous droits
DÉFICITS IMMUNITAIRE COMMUN VARIABLE
 DÉFICITS IMMUNITAIRE COMMUN VARIABLE Le présent livret a été rédigé à l attention des patients et de leurs familles. Il ne doit pas remplacer les conseils d un spécialiste en immunologie. 1 Egalement Disponible
DÉFICITS IMMUNITAIRE COMMUN VARIABLE Le présent livret a été rédigé à l attention des patients et de leurs familles. Il ne doit pas remplacer les conseils d un spécialiste en immunologie. 1 Egalement Disponible
COUSIN Fabien KERGOURLAY Gilles. 19 octobre 2007. de l hôte par les. Master 2 MFA Responsable : UE Incidence des paramètres environnementaux
 COUSIN Fabien KERGOURLAY Gilles 19 octobre 2007 Inhibition des défenses de l hôte par les bactéries pathogènes Master 2 MFA Responsable : UE Incidence des paramètres environnementaux Gwennola ERMEL I Détection
COUSIN Fabien KERGOURLAY Gilles 19 octobre 2007 Inhibition des défenses de l hôte par les bactéries pathogènes Master 2 MFA Responsable : UE Incidence des paramètres environnementaux Gwennola ERMEL I Détection
L immunoenzymologie. Technique puissante couramment utilisée e en recherche et en diagnostic cificité des anticorps pour leurs nes
 L immunoenzymologie Technique puissante couramment utilisée e en recherche et en diagnostic Basée e sur la très s grande spécificit cificité des anticorps pour leurs antigènes nes Test qualitatif Détection
L immunoenzymologie Technique puissante couramment utilisée e en recherche et en diagnostic Basée e sur la très s grande spécificit cificité des anticorps pour leurs antigènes nes Test qualitatif Détection
Structure quantique cohérente et incohérente de l eau liquide
 Structure quantique cohérente et incohérente de l eau liquide Prof. Marc HENRY Chimie Moléculaire du Solide Institut Le Bel, 4, Rue Blaise Pascal 67070 Strasbourg Cedex, France Tél: 03.68.85.15.00 e-mail:
Structure quantique cohérente et incohérente de l eau liquide Prof. Marc HENRY Chimie Moléculaire du Solide Institut Le Bel, 4, Rue Blaise Pascal 67070 Strasbourg Cedex, France Tél: 03.68.85.15.00 e-mail:
Contrôle de l'expression génétique : Les régulations post-transcriptionnelles
 Contrôle de l'expression génétique : Les régulations post-transcriptionnelles http://perso.univ-rennes1.fr/serge.hardy/ utilisateur : biochimie mot de passe : 2007 L'ARNm, simple intermédiaire entre le
Contrôle de l'expression génétique : Les régulations post-transcriptionnelles http://perso.univ-rennes1.fr/serge.hardy/ utilisateur : biochimie mot de passe : 2007 L'ARNm, simple intermédiaire entre le
www.gbo.com/bioscience 1 Culture Cellulaire Microplaques 2 HTS- 3 Immunologie/ HLA 4 Microbiologie/ Bactériologie Containers 5 Tubes/ 6 Pipetage
 2 HTS 3 Immunologie / Immunologie Informations Techniques 3 I 2 ELISA 96 Puits 3 I 4 ELISA 96 Puits en Barrettes 3 I 6 en Barrettes de 8 Puits 3 I 7 en Barrettes de 12 Puits 3 I 8 en Barrettes de 16 Puits
2 HTS 3 Immunologie / Immunologie Informations Techniques 3 I 2 ELISA 96 Puits 3 I 4 ELISA 96 Puits en Barrettes 3 I 6 en Barrettes de 8 Puits 3 I 7 en Barrettes de 12 Puits 3 I 8 en Barrettes de 16 Puits
Compléments - Chapitre 5 Spectroscopie
 ompléments - hapitre 5 Spectroscopie Spectroscopie par résonance magnétique nucléaire (RMN 13 ) Tandis que la spectroscopie RMN 1 H fournit des données sur la disposition des atomes d'hydrogène dans une
ompléments - hapitre 5 Spectroscopie Spectroscopie par résonance magnétique nucléaire (RMN 13 ) Tandis que la spectroscopie RMN 1 H fournit des données sur la disposition des atomes d'hydrogène dans une
Rôle des acides biliaires dans la régulation de l homéostasie du glucose : implication de FXR dans la cellule bêta-pancréatique
 Rôle des acides biliaires dans la régulation de l homéostasie du glucose : implication de FXR dans la cellule bêta-pancréatique Tuteur : Anne Muhr-Tailleux cardiovasculaires et diabète (Equipe 1) Institut
Rôle des acides biliaires dans la régulation de l homéostasie du glucose : implication de FXR dans la cellule bêta-pancréatique Tuteur : Anne Muhr-Tailleux cardiovasculaires et diabète (Equipe 1) Institut
Les tests thyroïdiens
 Les tests thyroïdiens Mise à jour de juin 2007 par S. Fatio Réf: les images et les graphiques sont tirés de www.endotext.org (textbook d endocrinologie online et libre d accès) QuickTime et un décompresseur
Les tests thyroïdiens Mise à jour de juin 2007 par S. Fatio Réf: les images et les graphiques sont tirés de www.endotext.org (textbook d endocrinologie online et libre d accès) QuickTime et un décompresseur
EXERCICES : MECANISMES DE L IMMUNITE : pages 406 407 408 409 410
 EXERCICES : MECANISMES DE L IMMUNITE : pages 406 407 408 409 410 EXERCICE 1 PAGE 406 : EXPERIENCES A INTERPRETER Question : rôles respectifs du thymus et de la moelle osseuse dans la production des lymphocytes.
EXERCICES : MECANISMES DE L IMMUNITE : pages 406 407 408 409 410 EXERCICE 1 PAGE 406 : EXPERIENCES A INTERPRETER Question : rôles respectifs du thymus et de la moelle osseuse dans la production des lymphocytes.
Les Champs Magnétiques
 Les Champs Magnétiques Guillaume Laurent To cite this version: Guillaume Laurent. Les Champs Magnétiques. École thématique. Assistants de prévention, Paris, France. 2014, pp.31. HAL Id:
Les Champs Magnétiques Guillaume Laurent To cite this version: Guillaume Laurent. Les Champs Magnétiques. École thématique. Assistants de prévention, Paris, France. 2014, pp.31. HAL Id:
Thierry DELZESCAUX. «biopicsel» group, URA CNRS-CEA 2210 Service MIRCen, I²BM, CEA Fontenay-aux-Roses, France. Thierry.Delzescaux@cea.
 Thierry DELZESCAUX «biopicsel» group, URA CNRS-CEA 2210 Service MIRCen, I²BM, CEA Fontenay-aux-Roses, France Thierry.Delzescaux@cea.fr Thierry DELZESCAUX Ingénieur chercheur CEA Experience Since 2008:
Thierry DELZESCAUX «biopicsel» group, URA CNRS-CEA 2210 Service MIRCen, I²BM, CEA Fontenay-aux-Roses, France Thierry.Delzescaux@cea.fr Thierry DELZESCAUX Ingénieur chercheur CEA Experience Since 2008:
Le grand écart de la recherche fondamentale vers l application biomédicale
 Le grand écart de la recherche fondamentale vers l application biomédicale Ludger Johannes «Chemical Biology of Membranes and Therapeutic Delivery» unit U1143 INSERM UMR3666 CNRS «Endocytic Trafficking
Le grand écart de la recherche fondamentale vers l application biomédicale Ludger Johannes «Chemical Biology of Membranes and Therapeutic Delivery» unit U1143 INSERM UMR3666 CNRS «Endocytic Trafficking
Sur le grossissement des divers appareils pour la mesure des angles par la réflexion d un faisceau lumineux sur un miroir mobile
 Sur le grossissement des divers appareils pour la mesure des angles par la réflexion d un faisceau lumineux sur un miroir mobile W. Lermantoff To cite this version: W. Lermantoff. Sur le grossissement
Sur le grossissement des divers appareils pour la mesure des angles par la réflexion d un faisceau lumineux sur un miroir mobile W. Lermantoff To cite this version: W. Lermantoff. Sur le grossissement
Le trajet des aliments dans l appareil digestif.
 La digestion. La digestion, c est la transformation des aliments en nutriments assimilables par l organisme. Dans le tube digestif, les aliments subissent une série de dégradations mécaniques et chimiques
La digestion. La digestion, c est la transformation des aliments en nutriments assimilables par l organisme. Dans le tube digestif, les aliments subissent une série de dégradations mécaniques et chimiques
Transplantation pulmonaire et mucoviscidose. Optimiser la prise en charge médicale
 Transplantation pulmonaire et mucoviscidose Optimiser la prise en charge médicale Dr Isabelle Danner-Boucher CRCM adulte de Nantes Unité de Transplantation Thoracique 11èmes Journées Scientifiques de la
Transplantation pulmonaire et mucoviscidose Optimiser la prise en charge médicale Dr Isabelle Danner-Boucher CRCM adulte de Nantes Unité de Transplantation Thoracique 11èmes Journées Scientifiques de la
Université Louis Pasteur Strasbourg I Ecole Doctorale des Sciences de la Vie et de la Santé THESE. En vue de l obtention du grade de
 Université Louis Pasteur Strasbourg I Ecole Doctorale des Sciences de la Vie et de la Santé THESE En vue de l obtention du grade de Docteur de l Université Louis Pasteur de Strasbourg Discipline : Aspects
Université Louis Pasteur Strasbourg I Ecole Doctorale des Sciences de la Vie et de la Santé THESE En vue de l obtention du grade de Docteur de l Université Louis Pasteur de Strasbourg Discipline : Aspects
Explorations des réponses Immunitaires. L3 Médecine
 2012 Explorations des réponses Immunitaires L3 Médecine Rappel sur les réponses Immunitaires DIFFERENTS TYPES DE REPONSES IMMUNITAIRES Naturelle = innée Adaptative Non spécifique Spécifique Immédiate Barrière
2012 Explorations des réponses Immunitaires L3 Médecine Rappel sur les réponses Immunitaires DIFFERENTS TYPES DE REPONSES IMMUNITAIRES Naturelle = innée Adaptative Non spécifique Spécifique Immédiate Barrière
Les marqueurs biologiques des tumeurs endocrines digestives
 Les marqueurs biologiques des tumeurs endocrines digestives Michèle d Herbomez DIU de Chirurgie Endocrinienne Lille 2009 Bilan biologique participe au Diagnostic Pronostic Suivi Trois types de marqueurs:
Les marqueurs biologiques des tumeurs endocrines digestives Michèle d Herbomez DIU de Chirurgie Endocrinienne Lille 2009 Bilan biologique participe au Diagnostic Pronostic Suivi Trois types de marqueurs:
Chapitre II La régulation de la glycémie
 Chapitre II La régulation de la glycémie Glycémie : concentration de glucose dans le sang valeur proche de 1g/L Hypoglycémie : perte de connaissance, troubles de la vue, voire coma. Hyperglycémie chronique
Chapitre II La régulation de la glycémie Glycémie : concentration de glucose dans le sang valeur proche de 1g/L Hypoglycémie : perte de connaissance, troubles de la vue, voire coma. Hyperglycémie chronique
Christian TREPO, MD, PhD
 PEG INTERFERON MONOTHERAPI E Christian TREPO, MD, PhD Unmet medical needs in chronic HBV infection Inhibition of viral replication Normalisation of ALT Improvement in liver necroinflammation Improvement
PEG INTERFERON MONOTHERAPI E Christian TREPO, MD, PhD Unmet medical needs in chronic HBV infection Inhibition of viral replication Normalisation of ALT Improvement in liver necroinflammation Improvement
CONTROVERSE : IDR OU QUANTIFERON LORS D'UN CONTAGE EN EHPAD?
 CONTROVERSE : IDR OU QUANTIFERON LORS D'UN CONTAGE EN EHPAD? Hélène MANGEARD François MALADRY Tuberculose : infection mycobactérienne Infection mycobactérienne chronique (M. Tuberculosis ++ ou bacille
CONTROVERSE : IDR OU QUANTIFERON LORS D'UN CONTAGE EN EHPAD? Hélène MANGEARD François MALADRY Tuberculose : infection mycobactérienne Infection mycobactérienne chronique (M. Tuberculosis ++ ou bacille
MAB Solut. vos projets. MABLife Génopole Campus 1 5 rue Henri Desbruères 91030 Evry Cedex. www.mabsolut.com. intervient à chaque étape de
 Mabsolut-DEF-HI:Mise en page 1 17/11/11 17:45 Page1 le département prestataire de services de MABLife de la conception à la validation MAB Solut intervient à chaque étape de vos projets Création d anticorps
Mabsolut-DEF-HI:Mise en page 1 17/11/11 17:45 Page1 le département prestataire de services de MABLife de la conception à la validation MAB Solut intervient à chaque étape de vos projets Création d anticorps
Le stress oxydant provoqué par l'exercice : une fatalité?
 Le stress oxydant provoqué par l'exercice : une fatalité? L e métabolisme aérobie est un moyen très efficace de produire de l'énergie en oxydant la matière organique, mais l'utilisation d'oxygène entraîne
Le stress oxydant provoqué par l'exercice : une fatalité? L e métabolisme aérobie est un moyen très efficace de produire de l'énergie en oxydant la matière organique, mais l'utilisation d'oxygène entraîne
Activité 38 : Découvrir comment certains déchets issus de fonctionnement des organes sont éliminés de l organisme
 Activité 38 : Découvrir comment certains déchets issus de fonctionnement des organes sont éliminés de l organisme 1. EXTRAITS REFERENTIELS DU BO Partie du programme : Fonctionnement de l organisme et besoin
Activité 38 : Découvrir comment certains déchets issus de fonctionnement des organes sont éliminés de l organisme 1. EXTRAITS REFERENTIELS DU BO Partie du programme : Fonctionnement de l organisme et besoin
1ST2S Biophysiopathologie : Motricité et système nerveux. 2012-2013. La physiologie neuro-musculaire :
 La physiologie neuro-musculaire : Introduction : Le neurone possède 3 propriétés indispensables pour assurer sa fonction au sein de l organisme : -l excitabilité : capacité à répondre à un stimulus -la
La physiologie neuro-musculaire : Introduction : Le neurone possède 3 propriétés indispensables pour assurer sa fonction au sein de l organisme : -l excitabilité : capacité à répondre à un stimulus -la
Utilisation des substrats énergétiques
 Utilisation des substrats énergétiques Collège des Enseignants de Nutrition Date de création du document 2010-2011 Table des matières I Les organes et les substrats... 3 I.1 Les substrats énergétiques...
Utilisation des substrats énergétiques Collège des Enseignants de Nutrition Date de création du document 2010-2011 Table des matières I Les organes et les substrats... 3 I.1 Les substrats énergétiques...
BTS BAT 1 Notions élémentaires de chimie 1
 BTS BAT 1 Notions élémentaires de chimie 1 I. L ATOME NOTIONS EÉLEÉMENTAIRES DE CIMIE Les atomes sont des «petits grains de matière» qui constituent la matière. L atome est un système complexe que l on
BTS BAT 1 Notions élémentaires de chimie 1 I. L ATOME NOTIONS EÉLEÉMENTAIRES DE CIMIE Les atomes sont des «petits grains de matière» qui constituent la matière. L atome est un système complexe que l on
Dr Pascale Vergne-Salle Service de Rhumatologie, CHU de Limoges. Membre enseignant chercheur EA 4021
 Dr Pascale Vergne-Salle Service de Rhumatologie, CHU de Limoges Membre enseignant chercheur EA 4021 Rhumatisme inflammatoire chronique Prévalence des SA ies en Europe = 1 à 2% Age moy : 26 ans, ratio homme/femme:
Dr Pascale Vergne-Salle Service de Rhumatologie, CHU de Limoges Membre enseignant chercheur EA 4021 Rhumatisme inflammatoire chronique Prévalence des SA ies en Europe = 1 à 2% Age moy : 26 ans, ratio homme/femme:
Respiration Mitochondriale
 Université Pierre et Marie Curie Respiration Mitochondriale Objectifs au cours de Révisions Biochimie PCEM2 Révisions Biochimie Métabolique 2004-2005 Pr. A. Raisonnier (alain.raisonnier@upmc.fr) Mise à
Université Pierre et Marie Curie Respiration Mitochondriale Objectifs au cours de Révisions Biochimie PCEM2 Révisions Biochimie Métabolique 2004-2005 Pr. A. Raisonnier (alain.raisonnier@upmc.fr) Mise à
TUTORAT UE 5 2012-2013 Anatomie Correction Séance n 6 Semaine du 11/03/2013
 TUTORAT UE 5 2012-2013 Anatomie Correction Séance n 6 Semaine du 11/03/2013 Appareil respiratoire-organogénèse de l appareil digestif Cours du Pr. PRUDHOMME Séance préparée par Anissa BARAKAT, Caroline
TUTORAT UE 5 2012-2013 Anatomie Correction Séance n 6 Semaine du 11/03/2013 Appareil respiratoire-organogénèse de l appareil digestif Cours du Pr. PRUDHOMME Séance préparée par Anissa BARAKAT, Caroline
TUMEURS DU BAS APPAREIL URINAIRE
 1 Plan de cours I - TUMEURS PRIMITIVES DE LA VESSIE c1 - Tumeurs papillaires non infiltrantes c2 - Tumeurs papillaires infiltrantes c3 - Carcinome in-situ en muqueuse plane D - Pronostic : II - TUMEURS
1 Plan de cours I - TUMEURS PRIMITIVES DE LA VESSIE c1 - Tumeurs papillaires non infiltrantes c2 - Tumeurs papillaires infiltrantes c3 - Carcinome in-situ en muqueuse plane D - Pronostic : II - TUMEURS
Résistance du VIH-1 aux antirétroviraux dans les compartiments anatomiques et cellulaires
 Résistance du VIH-1 aux antirétroviraux dans les compartiments anatomiques et cellulaires Jade GHOSN Laboratoire de Virologie CHU Necker-Enfants Malades EA MRT 3620 Paris 5 Réservoirs anatomiques du VIH:
Résistance du VIH-1 aux antirétroviraux dans les compartiments anatomiques et cellulaires Jade GHOSN Laboratoire de Virologie CHU Necker-Enfants Malades EA MRT 3620 Paris 5 Réservoirs anatomiques du VIH:
UE2 CANCEROLOGIE Place de la Médecine Nucléaire
 UE2 CANCEROLOGIE Place de la Médecine Nucléaire Imagerie Imagerie de Gamma Caméra Tomographie d émission monophotoniqueou TEMP: radiopharmaceutiqueémetteurs de rayonnement Gamma Imagerie de Caméra TEP
UE2 CANCEROLOGIE Place de la Médecine Nucléaire Imagerie Imagerie de Gamma Caméra Tomographie d émission monophotoniqueou TEMP: radiopharmaceutiqueémetteurs de rayonnement Gamma Imagerie de Caméra TEP
DIAPOSITIVE 1 Cette présentation a trait à la réglementation sur les thérapies cellulaires.
 Produits de thérapie cellulaire DIAPOSITIVE 1 Cette présentation a trait à la réglementation sur les thérapies cellulaires. DIAPOSITIVE 2 La fabrication des thérapies cellulaires est examinée par la Division
Produits de thérapie cellulaire DIAPOSITIVE 1 Cette présentation a trait à la réglementation sur les thérapies cellulaires. DIAPOSITIVE 2 La fabrication des thérapies cellulaires est examinée par la Division
La Dysplasie Ventriculaire Droite Arythmogène
 1 La Dysplasie Ventriculaire Droite Arythmogène Document rédigé par l équipe pluridisciplinaire du centre de référence pour les maladies cardiaques héréditaires (Paris), en collaboration avec des patients
1 La Dysplasie Ventriculaire Droite Arythmogène Document rédigé par l équipe pluridisciplinaire du centre de référence pour les maladies cardiaques héréditaires (Paris), en collaboration avec des patients
Hépatite B. Le virus Structure et caractéristiques 07/02/2013
 Hépatite B Le virus Structure et caractéristiques o o o Famille des Hepadnaviridae Genre orthohepadnavirus Enveloppé, capside icosaédrique, 42 nm 1 Le virus Structure et caractéristiques En microscopie
Hépatite B Le virus Structure et caractéristiques o o o Famille des Hepadnaviridae Genre orthohepadnavirus Enveloppé, capside icosaédrique, 42 nm 1 Le virus Structure et caractéristiques En microscopie
Calcaire ou eau agressive en AEP : comment y remédier?
 Calcaire ou eau agressive en AEP : comment y remédier? Les solutions techniques Principes et critères de choix Par Sébastien LIBOZ - Hydrogéologue Calcaire ou eau agressive en AEP : comment y remédier?
Calcaire ou eau agressive en AEP : comment y remédier? Les solutions techniques Principes et critères de choix Par Sébastien LIBOZ - Hydrogéologue Calcaire ou eau agressive en AEP : comment y remédier?
M2.6. 2012 Cancer du colon et du rectum. Coordination : Pr Hahnloser
 M2.6. 2012 Cancer du colon et du rectum Coordination : Pr Hahnloser Pathologie Dr. Maryse Fiche Cancers colo-rectaux Précurseurs : Images et figures : Robbins 8è Edition Objectifs d apprentissage : SCLO
M2.6. 2012 Cancer du colon et du rectum Coordination : Pr Hahnloser Pathologie Dr. Maryse Fiche Cancers colo-rectaux Précurseurs : Images et figures : Robbins 8è Edition Objectifs d apprentissage : SCLO
Formavie 2010. 2 Différentes versions du format PDB...3. 3 Les champs dans les fichiers PDB...4. 4 Le champ «ATOM»...5. 6 Limites du format PDB...
 Formavie 2010 Les fichiers PDB Les fichiers PDB contiennent les informations qui vont permettre à des logiciels de visualisation moléculaire (ex : RasTop ou Jmol) d afficher les molécules. Un fichier au
Formavie 2010 Les fichiers PDB Les fichiers PDB contiennent les informations qui vont permettre à des logiciels de visualisation moléculaire (ex : RasTop ou Jmol) d afficher les molécules. Un fichier au
Travaux dirigés de Microbiologie Master I Sciences des Génomes et des Organismes Janvier 2015
 Andrew Tolonen atolonen@genoscope.cns.fr Travaux dirigés de Microbiologie Master I Sciences des Génomes et des Organismes Janvier 2015 A- Généralités I- La vie sur terre telle que nous la connaissons ne
Andrew Tolonen atolonen@genoscope.cns.fr Travaux dirigés de Microbiologie Master I Sciences des Génomes et des Organismes Janvier 2015 A- Généralités I- La vie sur terre telle que nous la connaissons ne
P.L.E.A.S.E. Painless Laser Epidermal System Needle-free drug delivery. Arne Heinrich arne.heinrich@pantec-biosolutions.com
 P.L.E.A.S.E. Painless Laser Epidermal System Needle-free drug delivery Arne Heinrich arne.heinrich@pantec-biosolutions.com Groupe Pantec: 19 ans de tradition en ingénierie avec spécialisation en mécatronique
P.L.E.A.S.E. Painless Laser Epidermal System Needle-free drug delivery Arne Heinrich arne.heinrich@pantec-biosolutions.com Groupe Pantec: 19 ans de tradition en ingénierie avec spécialisation en mécatronique
Hémochromatose génétique non liée à HFE-1 : quand et comment la rechercher? Cécilia Landman 11 décembre 2010
 Hémochromatose génétique non liée à HFE-1 : quand et comment la rechercher? Cécilia Landman 11 décembre 2010 Métabolisme du fer : hepcidine Fer absorbé par les entérocytes des villosités duodénales : transporteur
Hémochromatose génétique non liée à HFE-1 : quand et comment la rechercher? Cécilia Landman 11 décembre 2010 Métabolisme du fer : hepcidine Fer absorbé par les entérocytes des villosités duodénales : transporteur
Biomarqueurs en Cancérologie
 Biomarqueurs en Cancérologie Définition, détermination, usage Biomarqueurs et Cancer: définition Anomalie(s) quantitative(s) ou qualitative(s) Indicative(s) ou caractéristique(s) d un cancer ou de certaines
Biomarqueurs en Cancérologie Définition, détermination, usage Biomarqueurs et Cancer: définition Anomalie(s) quantitative(s) ou qualitative(s) Indicative(s) ou caractéristique(s) d un cancer ou de certaines
Moyens d étude de la peau
 Moyens d étude de la peau Dr. Yannick Le Corre yalecorre@chu-angers.fr Service de Dermatologie Vénéréologie Pr. L. Martin 2011-2012 CHU Angers UE Revêtement cutané Dermoscopie ou Dermatoscopie Examen de
Moyens d étude de la peau Dr. Yannick Le Corre yalecorre@chu-angers.fr Service de Dermatologie Vénéréologie Pr. L. Martin 2011-2012 CHU Angers UE Revêtement cutané Dermoscopie ou Dermatoscopie Examen de
AGROBASE : un système de gestion de données expérimentales
 AGROBASE : un système de gestion de données expérimentales Daniel Wallach, Jean-Pierre RELLIER To cite this version: Daniel Wallach, Jean-Pierre RELLIER. AGROBASE : un système de gestion de données expérimentales.
AGROBASE : un système de gestion de données expérimentales Daniel Wallach, Jean-Pierre RELLIER To cite this version: Daniel Wallach, Jean-Pierre RELLIER. AGROBASE : un système de gestion de données expérimentales.
Cordarone et Thyroïde par François Boustani
 Physiologie de la thyroïde : l hormonosynthèse thyroïdienne se fait à partir de l iode essentiellement d origine digestive et de la thyroglobuline qui est une protéine synthétisée par la thyroïde à partir
Physiologie de la thyroïde : l hormonosynthèse thyroïdienne se fait à partir de l iode essentiellement d origine digestive et de la thyroglobuline qui est une protéine synthétisée par la thyroïde à partir
Parrainage par Monsieur Philippe PAREIGE de notre classe, presentation des nanotechnologies.
 LUDIVINE TASSERY 1ere S 5 Parrainage par Monsieur Philippe PAREIGE de notre classe, presentation des nanotechnologies. Lors de la seconde visite, Monsieur PAREIGE, nous a parlé des nanotechnologies et
LUDIVINE TASSERY 1ere S 5 Parrainage par Monsieur Philippe PAREIGE de notre classe, presentation des nanotechnologies. Lors de la seconde visite, Monsieur PAREIGE, nous a parlé des nanotechnologies et
Advanced Fluorescence Microscopy to Study Plasma Membrane Protein Dynamics
 Advanced Fluorescence Microscopy to Study Plasma Membrane Protein Dynamics THÈSE N O 4869 (2010) PRÉSENTÉE LE 5 NOVEMBRE 2010 À LA FACULTÉ SCIENCES DE BASE LABORATOIRE DE CHIMIE PHYSIQUE DES POLYMÈRES
Advanced Fluorescence Microscopy to Study Plasma Membrane Protein Dynamics THÈSE N O 4869 (2010) PRÉSENTÉE LE 5 NOVEMBRE 2010 À LA FACULTÉ SCIENCES DE BASE LABORATOIRE DE CHIMIE PHYSIQUE DES POLYMÈRES
2 C est quoi la chimie?
 PARTIE 1 AVANT LA CHIMIE VERTE... 2 C est quoi la chimie? L inconnu étant source d angoisse, nous allons essayer de définir les grands domaines de la chimie pour mieux la connaître, l appréhender et donc
PARTIE 1 AVANT LA CHIMIE VERTE... 2 C est quoi la chimie? L inconnu étant source d angoisse, nous allons essayer de définir les grands domaines de la chimie pour mieux la connaître, l appréhender et donc
1 ère partie : tous CAP sauf hôtellerie et alimentation CHIMIE ETRE CAPABLE DE. PROGRAMME - Atomes : structure, étude de quelques exemples.
 Référentiel CAP Sciences Physiques Page 1/9 SCIENCES PHYSIQUES CERTIFICATS D APTITUDES PROFESSIONNELLES Le référentiel de sciences donne pour les différentes parties du programme de formation la liste
Référentiel CAP Sciences Physiques Page 1/9 SCIENCES PHYSIQUES CERTIFICATS D APTITUDES PROFESSIONNELLES Le référentiel de sciences donne pour les différentes parties du programme de formation la liste
Séquence 1. Glycémie et diabète. Sommaire
 Séquence 1 Glycémie et diabète Sommaire 1. Prérequis 2. La régulation de la glycémie 3. Le diabète 4. Synthè se 5. Glossaire 6. Exercices Exercices d apprentissage du chapitre 2 Exercices d apprentissage
Séquence 1 Glycémie et diabète Sommaire 1. Prérequis 2. La régulation de la glycémie 3. Le diabète 4. Synthè se 5. Glossaire 6. Exercices Exercices d apprentissage du chapitre 2 Exercices d apprentissage
Résistance du virus de l hépatite C aux nouveaux traitements anti-viraux
 14 ème forum SOS Hépatites, Lyon Résistance du virus de l hépatite C aux nouveaux traitements anti-viraux Dr. Sylvie LARRAT Laboratoire de Virologie CHU Grenoble Hépatite C chronique 180 millions de porteurs
14 ème forum SOS Hépatites, Lyon Résistance du virus de l hépatite C aux nouveaux traitements anti-viraux Dr. Sylvie LARRAT Laboratoire de Virologie CHU Grenoble Hépatite C chronique 180 millions de porteurs
5.5.5 Exemple d un essai immunologique
 5.5.5 Exemple d un essai immunologique Test de grossesse Test en forme de bâtonnet destiné à mettre en évidence l'hormone spécifique de la grossesse, la gonadotrophine chorionique humaine (hcg), une glycoprotéine.
5.5.5 Exemple d un essai immunologique Test de grossesse Test en forme de bâtonnet destiné à mettre en évidence l'hormone spécifique de la grossesse, la gonadotrophine chorionique humaine (hcg), une glycoprotéine.
Professeur Diane GODIN-RIBUOT
 UE3-2 - Physiologie rénale Chapitre 5 : Mesure de la fonction rénale : la clairance rénale Professeur Diane GODIN-RIBUOT Année universitaire 2011/2012 Université Joseph Fourier de Grenoble - Tous droits
UE3-2 - Physiologie rénale Chapitre 5 : Mesure de la fonction rénale : la clairance rénale Professeur Diane GODIN-RIBUOT Année universitaire 2011/2012 Université Joseph Fourier de Grenoble - Tous droits
1. LES ENTRAINEMENTS SPORTIFS
 ALLER PLUS HAUT POUR ÊTRE PLUS FORT? 1. LES ENTRAINEMENTS SPORTIFS Pour introduire le chapitre, voici 3 extraits d articles abordant les entrainements sportifs. Lis-les attentivement et réponds aux questions
ALLER PLUS HAUT POUR ÊTRE PLUS FORT? 1. LES ENTRAINEMENTS SPORTIFS Pour introduire le chapitre, voici 3 extraits d articles abordant les entrainements sportifs. Lis-les attentivement et réponds aux questions
Programme AcSé. Accès Sécurisé aux Innovations Thérapeutiques Deux études pilotes : AcSé - crizotinib et AcSé - vémurafenib
 Programme AcSé Accès Sécurisé aux Innovations Thérapeutiques Deux études pilotes : AcSé - crizotinib et AcSé - vémurafenib G. Vassal, J.Y. Blay, C. Cailliot, N. Hoog-Labouret, F. Denjean, M. Jimenez, A.L.
Programme AcSé Accès Sécurisé aux Innovations Thérapeutiques Deux études pilotes : AcSé - crizotinib et AcSé - vémurafenib G. Vassal, J.Y. Blay, C. Cailliot, N. Hoog-Labouret, F. Denjean, M. Jimenez, A.L.
Médicaments du futur : Tendances et enjeux. Nicolas PY, Debiopharm International forumofac.14 26/09/2014
 Médicaments du futur : Tendances et enjeux Nicolas PY, Debiopharm International forumofac.14 26/09/2014 Quelques mots sur Debiopharm Groupe fondé en 1979 Siège à Lausanne 350 collaborateurs Financièrement
Médicaments du futur : Tendances et enjeux Nicolas PY, Debiopharm International forumofac.14 26/09/2014 Quelques mots sur Debiopharm Groupe fondé en 1979 Siège à Lausanne 350 collaborateurs Financièrement
Traitement des hépatites h chroniques virales B et C
 Traitement des hépatites h chroniques virales B et C Dr Joanna POFELSKI Service d hépatod pato-gastroentérologierologie CH d Annecyd Particularités? Virus hépatotropes / cytopathogènes : lésions hépatiques
Traitement des hépatites h chroniques virales B et C Dr Joanna POFELSKI Service d hépatod pato-gastroentérologierologie CH d Annecyd Particularités? Virus hépatotropes / cytopathogènes : lésions hépatiques
Y a-t-il une place pour un vaccin thérapeutique contre l hépatite B? H. Fontaine, Unité d Hépatologie Médicale, Hôpital Cochin
 Y a-t-il une place pour un vaccin thérapeutique contre l hépatite B? H. Fontaine, Unité d Hépatologie Médicale, Hôpital Cochin Objectifs thérapeutiques Inhibition de la réplication virale Négativation
Y a-t-il une place pour un vaccin thérapeutique contre l hépatite B? H. Fontaine, Unité d Hépatologie Médicale, Hôpital Cochin Objectifs thérapeutiques Inhibition de la réplication virale Négativation
Étude des formes de pratiques de la gymnastique sportive enseignées en EPS à l école primaire
 Étude des formes de pratiques de la gymnastique sportive enseignées en EPS à l école primaire Stéphanie Demonchaux To cite this version: Stéphanie Demonchaux. Étude des formes de pratiques de la gymnastique
Étude des formes de pratiques de la gymnastique sportive enseignées en EPS à l école primaire Stéphanie Demonchaux To cite this version: Stéphanie Demonchaux. Étude des formes de pratiques de la gymnastique
TEST ELISA (ENZYME-LINKED IMMUNOSORBENT ASSEY)
") TEST ELISA (ENZYME-LINKED IMMUNOSORBENT ASSEY) Lise Vézina, technicienne de laboratoire Michel Lacroix, agronome-phytopathologiste Direction de l innovation scientifique et technologique Au Laboratoire
TEST ELISA (ENZYME-LINKED IMMUNOSORBENT ASSEY) Lise Vézina, technicienne de laboratoire Michel Lacroix, agronome-phytopathologiste Direction de l innovation scientifique et technologique Au Laboratoire
DYSKINESIE CILIAIRE PRIMITIVE BROCHURE D INFORMATION DESTINEE AUX PATIENTS ET LEUR FAMILLE QU EST-CE QUE LA DYSKINESIE CILIAIRE PRIMITIVE?
 QU EST-CE QUE LA DYSKINESIE CILIAIRE PRIMITIVE? COMMENT SE TRANSMET-ELLE? COMMENT SE MANIFESTE-T-ELLE? DYSKINESIE CILIAIRE PRIMITIVE BROCHURE D INFORMATION DESTINEE AUX PATIENTS ET LEUR FAMILLE COMMENT
QU EST-CE QUE LA DYSKINESIE CILIAIRE PRIMITIVE? COMMENT SE TRANSMET-ELLE? COMMENT SE MANIFESTE-T-ELLE? DYSKINESIE CILIAIRE PRIMITIVE BROCHURE D INFORMATION DESTINEE AUX PATIENTS ET LEUR FAMILLE COMMENT
Cibles et mécanismes d action des traitements par cytokines et anti-cytokines
 Cibles et mécanismes d action des traitements par cytokines et anti-cytokines Jean Daniel Lelièvre, Yves Lévy, Pierre Miossec I-Introduction... 2 II-Les interférons... 2 II-1.L interféron... 3 II-1-a.
Cibles et mécanismes d action des traitements par cytokines et anti-cytokines Jean Daniel Lelièvre, Yves Lévy, Pierre Miossec I-Introduction... 2 II-Les interférons... 2 II-1.L interféron... 3 II-1-a.
A B C Eau Eau savonneuse Eau + détergent
 1L : Physique et chimie dans la cuisine Chapitre.3 : Chimie et lavage I. Les savons et les détergents synthétiques 1. Propriétés détergentes des savons Le savon est un détergent naturel, les détergents
1L : Physique et chimie dans la cuisine Chapitre.3 : Chimie et lavage I. Les savons et les détergents synthétiques 1. Propriétés détergentes des savons Le savon est un détergent naturel, les détergents
L univers vivant De la cellule à l être humain
 L univers vivant De la cellule à l être humain La cellule humaine Voir : http://www.biologieenflash.net/sommaire.html Voir : http://fr.wikipedia.org/ La cellule humaine Unité de base de tous les êtres
L univers vivant De la cellule à l être humain La cellule humaine Voir : http://www.biologieenflash.net/sommaire.html Voir : http://fr.wikipedia.org/ La cellule humaine Unité de base de tous les êtres
LA MITOSE CUEEP - USTL DÉPARTEMENT SCIENCES BAHIJA DELATTRE
 Biologie LA MITOSE CUEEP - USTL DÉPARTEMENT SCIENCES BAHIJA DELATTRE Février 2006 I. L'INTRODUCTION Chaque cellule d'un organisme supérieur provient de la multiplication d'une cellule préexistante (cellule
Biologie LA MITOSE CUEEP - USTL DÉPARTEMENT SCIENCES BAHIJA DELATTRE Février 2006 I. L'INTRODUCTION Chaque cellule d'un organisme supérieur provient de la multiplication d'une cellule préexistante (cellule
Chapitre 4 : cohabiter avec les micro-organismes. Contrat-élève 3 ème
 Chapitre 4 : cohabiter avec les micro-organismes Mais, que dois-je savoir? Pour rattraper un cours manquant, retrouve-le sur le site du collège dans la rubrique «enseignements» : http://colleges.acrouen.fr/courbet/spipuser/
Chapitre 4 : cohabiter avec les micro-organismes Mais, que dois-je savoir? Pour rattraper un cours manquant, retrouve-le sur le site du collège dans la rubrique «enseignements» : http://colleges.acrouen.fr/courbet/spipuser/