VI- Expression du génome
|
|
|
- Michele Larochelle
- il y a 10 ans
- Total affichages :
Transcription

1 VI- Expression du génome

2 VI-1.- EXPRESSION DU GÉNOME- PRINCIPES GÉNÉRAUX

3 DOGME CENTRAL Les gènes et l information génétique sont conservés sous forme d acides nucléiques La perpétuation à l identique de l information génétique est assurée par la réplication Les gènes exercent leurs fonctions sous la forme de protéines. Le message qu ils contiennent doit donc être traduit en protéines : expression des gènes

4 EXPRESSION DE L INFORMATION GÉNÉTIQUE Processus en deux phases : 1. La transcription è ARN simple brin de séquence identique à l un des brins d ADN. Plusieurs types d ARN ARN messager ou ARNm ARN de transfert ou ARNt ARN ribosomal ou ARNr

5 EXPRESSION DE L INFORMATION (2) Processus en deux phases : La transcription Seul un des brins d ADN est transcrit en ARNm Brin matrice ou brin anti-sens : dirige la synthèse de l ARN par appariement de bases complémentaires Brin sens ou brin codant : même séquence que l ARNm (U remplace la T)

6 EXPRESSION DE L INFORMATION (3) Processus en deux phases : 2. La traduction è conversion séquence nucléotidique de l ARN en une chaîne d acides aminés qui constituent un peptide puis une protéine fonctionnelle L ARNm est traduit ARNt et ARNr font partie de la machinerie de synthèse protéique

7 EXPRESSION DE L INFORMATION (4) C est la région codante de l ARNm qui est traduite ; dans cette région chaque triplet de nucléotides (codon) correspond à un acide aminé.
 correspond à un acide")
8 EXPRESSION DE L INFORMATION (5) Chez les eucaryotes, le produit de la transcription (transcrit primaire) n est pas directement traduit il subit une série de modifications appelée maturation qui est constituée d une série d ajouts et d excisions. Puis migre dans le cytoplasme où il est traduit

9 EXPRESSION DE L INFORMATION (6) Nécessité d une correspondance entre séquence nucléotidique et séquence d acides aminés (entre l ADN et la protéine) : code génétique Procaryotes : transcription et traduction ont lieu dans le même compartiment et sont synchrones; Eucaryotes : transcription dans le noyau, traduction dans le cytoplasme

10 EXPRESSION DE L INFORMATION (7) L expression de l information est unidirectionnelle dans la quasitotalité des cas ADN ARN Protéines Irréversible

11 EXPRESSION DE L INFORMATION (8) L expression de l information est unidirectionnelle : exception chez les rétrovirus ADN Transcription inverse ARN génomique simple brin ARNm Protéines

12 VI-2.- EXPRESSION DU GÉNOME TRANSCRIPTION CHEZ LES PROCARYOTES (E. COLI)

13 VI-2.- EXPRESSION DU GÉNOME TRANSCRIPTION CHEZ LES PROCARYOTES (E. COLI) A- LES PRINCIPAUX ACTEURS
 A- LES PRINCIPAUX")
14 A) L ARN POLYMÉRASE Plusieurs milliers de copies par cellule L enzyme complète ou holoenzyme est constituée de plusieurs sous-unités : α 2 ββ σ sous-unités β et β : centre catalytique de l enzyme, sont impliquées dans toutes les étapes de la transcription

15 A) L ARN POLYMÉRASE (2) sous-unité α : assemblage du noyau de l enzyme, aucun rôle connu dans le mécanisme de la transcription rôle dans la reconnaissance du promoteur et dans l interaction de l ARN polymérase et de certains facteurs de régulation.

16 A) L ARN POLYMÉRASE (3) sous-unité σ : impliquée spécifiquement dans la reconnaissance des promoteurs,

17 A) L ARN POLYMÉRASE (4) noyau de l'enzyme : oligomère α 2 ββ capable de synthétiser de l ARN à partir d une matrice ADN mais ne peut initier la transcription au niveau des sites corrects, Peut se fixer à n importe quel ADN quelque soit sa séquence (attraction électrostatique)

18 A) L ARN POLYMÉRASE (5) le facteur σ : qui se lie spécifiquement au promoteur.

19 B) L ADN MATRICE Brin antisens Gène non morcelé On y distingue différentes séquences : séquences codantes et non codantes, point de départ, séquences de terminaison de la transcription

20 C) LES RÉGIONS PROMOTRICES OU PROMOTEUR Séquences d ADN nécessaires à la fixation de l ARN polymérase sur la matrice et à l exécution de la réaction d initiation

21 C) LES RÉGIONS PROMOTRICES OU PROMOTEUR (2) La séquence du promoteur est très importante Des études de comparaison de séquences de différents promoteurs ont été réalisées afin d identifier leurs caractéristiques nécessaires à leur reconnaissance par l ARN polymérase
22 C) LES RÉGIONS PROMOTRICES OU PROMOTEUR (3) Ces études ont retrouvé des séquences : variant peu d un promoteur à l autre présentes dans pratiquement tous les promoteurs Ces séquences sont dites conservées (faibles variations) A partir de ces études, des séquences consensus (séquences types) ont été retenues
23 C) LES RÉGIONS PROMOTRICES OU PROMOTEUR (4) Séquence consensus (appelées aussi boîte) : succession de bases pour laquelle l homologie est maximale pour différents promoteurs (pour chaque position, c est la base la plus fréquemment retrouvée qui a été retenue)
24 C) LES RÉGIONS PROMOTRICES OU PROMOTEUR (5) Il y a quatre séquences conservées chez E. coli : Le point de départ La séquence située en -10 La séquence 35 La distance entre les séquences 10 et -35
25 C) LES RÉGIONS PROMOTRICES OU PROMOTEUR (5) Le point de départ En général une purine On lui attribue la valeur +1 Avant le point de départ : séquences dites en amont (positionnement représenté par un nombre négatif) Après le point de départ : séquences en aval (nombre positif)
26 C) LES RÉGIONS PROMOTRICES OU PROMOTEUR (6) La séquence 10 : la séquence consensus ou boîte TATAAT ou TATAAT box Juste en amont du point de départ (environ 10 pb en amont) Région de 6 pb Présente dans la plupart des promoteurs
27 C) LES RÉGIONS PROMOTRICES OU PROMOTEUR (7) La séquence 35 : 35 pb en amont du point de départ Région de 6 pb Séquence consensus : TTGACA
28 C) LES RÉGIONS PROMOTRICES OU PROMOTEUR (8) La distance entre les sites 35 et -10 Varie de 16 à 19 pb Sa séquence n a aucun rôle mais sa longueur est primordiale
29 C) LES RÉGIONS PROMOTRICES OU PROMOTEUR (9) La distance entre les sites 35 et -10 Longueur permet de maintenir une distance appropriée entre région 10 et région 35 pour se conformer à la géométrie de l ARN polymérase
30 PROMOTEUR TYPE DES PROCARYOTES Point de départ - 35 Boîte TTGACA à 19 pb Boîte 5 à 9 pb TATAAT +1
31 VI-2.- EXPRESSION DU GÉNOME TRANSCRIPTION CHEZ LES PROCARYOTES (E. COLI) B- LES PRINCIPALES ÉTAPES
32 PRINCIPE DE LA TRANSCRIPTION La première paire de bases transcrite en ARN est le point de départ de la transcription. A partir de ce point, l ARN polymérase se déplace le long de la matrice en synthétisant l ARN jusqu à ce qu elle atteigne une séquence de terminaison (un terminateur).
33 PRINCIPE DE LA TRANSCRIPTION (2) Cette activité définit une unité de transcription (ADN) qui s étend du promoteur au terminateur. Une unité de transcription peut contenir plusieurs gènes.
34 PRINCIPE DE LA TRANSCRIPTION (3) Les séquences sont conventionnellement écrites de sorte que la transcription se déroule de gauche à droite. Ceci revient à écrire l ARNm dans le sens habituel 5 3.
35 LES DIFFÉRENTES ÉTAPES 1. La reconnaissance de la matrice et la formation de la bulle de transcription 2. L initiation 3. L élongation 4. La terminaison
36 A) LA RECONNAISSANCE DE LA MATRICE Débute par la liaison de l ARN polymérase sur un promoteur de l ADN double brin Puis séparation provisoire des brins d ADN è appariement du brin matrice avec les ribonucléotides
37 A) LA RECONNAISSANCE DE LA MATRICE (2) Création de la bulle de transcription par un déroulement local qui commence au niveau du site de liaison de l ARN polymérase sur le promoteur
38 B) L INITIATION L initiation représente la formation des premières liaisons nucléotidiques dans l ARN. L enzyme reste fixée au promoteur pendant la synthèse des 9 premières liaisons nucléotidiques environ.
39 B) L INITIATION (2) La phase d initiation s arrête lorsque l enzyme arrive à synthétiser une chaîne de plus de 9 bases et se détache du promoteur..
40 C) L ÉLONGATION L enzyme se déplace le long de l ADN et allonge la chaîne d ARN en croissance. En se déplaçant, l enzyme déroule l hélice d ADN pour exposer une nouvelle partie de la matrice dans un état simple-brin.
41 C) L ÉLONGATION (2) Les nucléotides sont ajoutés par des liaisons covalentes à l extrémité 3 de la chaîne naissante, formant un hybride ADN-ARN dans la région déroulée.
42 C) L ÉLONGATION (3) A l arrière de cette région déroulée, le brin matrice d ADN se ré-apparie avec son partenaire d origine pour reformer la double hélice
43 C) L ÉLONGATION (4) L ARN sort de l enzyme sous forme simple brin. Lors de l élongation, la bulle de transcription se déplace grâce à une perturbation de la structure d ADN, dans laquelle le brin matrice de la région transitoirement déroulée est appariée avec l ARN naissant au niveau du point d élongation.
44 D) LA TERMINAISON La terminaison est la reconnaissance de la région à partir de laquelle aucune base supplémentaire ne doit être ajoutée à la chaîne d ARN. Pour que la transcription se termine, la formation de liaisons phosphodiesters doit cesser et le complexe de transcription doit se détacher.
45 D) LA TERMINAISON (2) Lorsque la dernière base est ajoutée à la chaîne d ARN, la bulle de transcription se décroche en même temps que l ADN-ARN se sépare, le duplex d ADN se reforme puis l enzyme et l ARN sont tous deux libérés.
46 D) LA TERMINAISON (3) Les séquences d ADN nécessaires au déclenchement de ces réactions constituent des terminateurs.
47 D) LA TERMINAISON (4) La terminaison dépend de séquences d ADN donc également de celles de l ARN synthétisé. Elle est soumise à un contrôle spécifique.
48 VI-2.- EXPRESSION DU GÉNOME TRANSCRIPTION CHEZ LES PROCARYOTES (E. COLI) C- LES TERMINATEURS
49 2 types de terminateurs : Terminateurs intrinsèques : ces séquences sont suffisantes par elle mêmes pour mettre un terme à la transcription Terminateurs extrinsèques ont besoin d un facteur auxiliaire pour achever la transcription
50 A) TERMINATEURS INTRINSÈQUES ne nécessitent aucun facteur auxiliaire. ces séquences terminatrices se traduisent au niveau de l extrémité de l ARN par la présence d une structure en épingle à cheveux suivie d une succession de 6 résidus U : l épingle à cheveux et la séquence poly-u sont généralement séparés de 7 à 9 b.
51 TERMINATEURS INTRINSÈQUES (2) La structure en épingle à cheveux de l ARN est formée par l appariement de séquences répétées inversées (qui forment la tige) séparées par une courte séquence (qui forme la boucle). Longueur variable de la boucle. L épingle à cheveux ralentit la polymérase voire entraîne son arrêt. De plus, la formation des liaisons intra-chaines à l origine de l épingle à cheveux entre en compétition avec la formation de liaisons hydrogène entre l ADN et l ARN et donc déstabilise l hybride ADN-ARN
52 TERMINATEURS INTRINSÈQUES (3) La séquence poly-u entraîne la dissociation de l enzyme de la matrice pendant qu elle est arrêtée par l épingle à cheveux. La séquence poly-u de l ARN correspond à des séquences riches en A-T dans l ADN.

53 Séquences de 2 terminateurs intrinsèques
54 B) TERMINATEURS EXTRINSÈQUES Des terminateurs rho-dépendants qui nécessitent la présence du facteur rho. Rho est une protéine complexe Pas de séquences consensus clairement définies : grande diversité mais caractéristiques communes
55 B) TERMINATEURS EXTRINSÈQUES composés de deux éléments distincts répartis dans une séquence ADN de 150 à 200 pb environ (Lau et al., 1983; Richardson, 1990; Zalatan et al., 1993) Une séquence essentielle nommée rut (pour "Rho utilization") et située en amont du site de terminaison. Le site de terminaison proprement dit, où intervient le relargage du transcrit, aussi appelé tsp (pour "transcription stop point").
56 B) TERMINATEURS EXTRINSÈQUES La séquence rut La séquence rut de l ADN code pour le site d'entrée de Rho sur l'arn. Rho possède une forte affinité pour certains transcrits ARN Elle a 50 à 90 pb de long et, dans l ARN, est riche en résidus C (41%) et pauvre en G (14%). Les séquences terminatrices sont moins bien définies que les séquences rut.
57 TERMINATEURS EXTRINSÈQUES (2) Rho se fixe à l ARN et se déplace le long de l ARN «aux trousses» de l enzyme. Quand l ARN polymérase ralentit au niveau du site de terminaison, rho la rattrape Rho déroule l hybride ADN ARN dans la bulle de transcription è terminaison : ARN polymérase, rho et ARN sont libérés.
58 VI-2.- EXPRESSION DU GÉNOME TRANSCRIPTION CHEZ LES PROCARYOTES (E. COLI) D- LE CONTENU DE L ARNM
59 LE CONTENU DE L ARNM 2 types d ARNm Les ARN monocistroniques : = ARNm représentant un seul gène ARN polycistroniques La plupart des ARNs portent des séquences qui codent pour plusieurs protéines. Dans ce cas un seul ARNm est transcrit à partir d un groupe de gènes adjacents (impliqués souvent dans la même voie métabolique).
60 LE CONTENU DE L ARNM (2) Tous les ARNm contiennent deux types de régions Régions codantes Régions non codantes
61 LE CONTENU DE L ARNM (3) ARN monocistronique : une région codante constituée d une succession de codons représentant la séquence d acides aminés de la protéine qui débute en général par AUG et s achève par un codon de terminaison. Des régions supplémentaires non codantes. A l extrémité 5 se situe la tête qui précède le début de la région codante. La queue suit le codon de terminaison
62 LE CONTENU DE L ARNM (4) ARN polycistronique : Des régions intercistroniques entre les régions codantes ont des tailles très variables (1 ou 2 nt à 30 nt entre codon de terminaison de l un et codon initiateur de l autre). Possibilité de chevauchement de gènes : dernière base d une région codante = première base de la région codante suivante. nt = nucléotide
63 ARN monocistronique 5 3 Tête Région codante Queue ARN polycistronique 5 3 Tête Région codante 1 RIC Région codante 2 RIC Région codante 3 Queue RIC : Région intercistronique
64 VI-2.- EXPRESSION DU GÉNOME TRANSCRIPTION CHEZ LES PROCARYOTES (E. COLI) E- LE CYCLE DE L ARNM
65 LE CYCLE DE L ARNM Chez les procaryotes, les ribosomes se fixent à l ARNm avant la fin de la transcription. L ARNm bactérien est instable et n est traduit en protéines que pendant une courte durée de quelques minutes. Il est dégradée dans le sens 5-3 donc au fur à mesure qu il est traduit en protéine.
66 VI-2.- EXPRESSION DU GÉNOME TRANSCRIPTION CHEZ LES PROCARYOTES (E. COLI) F- LA RÉGULATION DE LA TRANSCRIPTION
67 La régulation de l initiation de la transcription est le niveau principal de régulation de l expression des gènes
68 JACOB ET MONOD (1961) La transcription est régulée par : des séquences agissant en cis : N importe quelle séquence d ADN qui agit en tant que séquence (n utilise pas d intermédiaire protéique) Qui ne touche que l ADN auquel elle est physiquement liée (ne peut réguler que les séquences qui lui sont contiguës) Exemple : le promoteur
69 JACOB ET MONOD (1961) La transcription est aussi régulée par : des facteurs agissant en trans : molécules synthétisées par un gène qui diffusent hors de leur lieu de synthèse pour aller agir sur un gène cible. Ces facteurs (protéine le plus souvent, parfois ARN) agissant en trans sont le produit d un gène régulateur.
70 GÈNE DE STRUCTURE Gène qui code pour : Des protéines structurales Des protéines à activité catalytique
71 GÈNE RÉGULATEUR Gène qui code pour une protéine impliquée dans la régulation d autres gènes Les protéines régulatrices de la transcription la contrôlent en se fixant à un ou plusieurs sites spécifiques de l ADN, en général situé juste en amont du gène cible
72 Les gènes bactériens qui codent pour des protéines apparentées (ex : enzymes participant aux différentes réactions d une même voie catabolique) sont transcrits en un ARNm polycistronique à partir d un seul promoteur. Le contrôle de ce promoteur régule l expression de la voie métabolique entière.
73 L unité de régulation qui contient les gènes structuraux et les éléments agissant en cis est appelée opéron.
74 L initiation de la transcription est régulée par des interactions qui se produisent au voisinage du promoteur. L initiation de la transcription par l ARN polymérase au niveau du promoteur est rendue possible ou impossible par d autres protéines.
75 Des gènes qui sont actifs sauf si des facteurs empêchent leur expression sont dits sous contrôle négatif. Des gènes qui ne sont transcrits que s ils sont spécifiquement activés sont dits sous contrôle positif (prédominant chez les eucaryotes).
76 Une protéine répresseur empêche à l ARN polymérase de se fixer au promoteur ou d activer la transcription.
77 Le répresseur se fixe sur une séquence cible, l opérateur, qui se trouve en général autour ou en amont du point de départ. La fixation d un répresseur sur son opérateur est régulée par une petite molécule : l inducteur empêche au répresseur de se fixer, le co-répresseur l active et lui permet cette fixation.
78 Certains promoteurs ne peuvent être reconnus par l ARN polymérase que si une protéine activatrice est présente. Les protéines activatrices peuvent également être régulées par de petites molécules.
79 EXEMPLE DE RÉGULATION CHEZ LES PROCARYOTES : L OPÉRON LACTOSE 3 gènes structuraux : lacz, lacy, laca Codent pour des protéines permettant de capter et de métaboliser des β-galactosides comme le lactose
80 EXEMPLE DE RÉGULATION CHEZ LES PROCARYOTES : L OPÉRON LACTOSE (2) lacz è β-galactosidase : coupe un β-galactoside et libère les sucres qui la composent lacy è β-galactoside perméase : transporte les β-galactosides dans la cellule laca è β-galactoside transacétylase : transfert d un résidu acétyl sur les β-galactosides
81 EXEMPLE DE RÉGULATION CHEZ LES PROCARYOTES : L OPÉRON LACTOSE (3) laczya : transcription contrôlée par une protéine régulatrice synthétisée à partir du gène laci laci peut se trouver à proximité des gènes de structure mais constitue une unité de transcription indépendante (transrégulateur)
82 EXEMPLE DE RÉGULATION CHEZ LES PROCARYOTES : L OPÉRON LACTOSE (3) laczya : régulation négative (transcrits sauf si protéine régulatrice) Produit de laci =répresseur lac Agit en se fixant sur opérateur Olac situé au début du groupe laczya
83 EXEMPLE DE RÉGULATION CHEZ LES PROCARYOTES : L OPÉRON LACTOSE (4) Opérateur Olac situé entre le promoteur (P lac ) et les gènes structuraux Fixation du répresseur sur Olac è blocage initiation de la transcription
84 EXEMPLE DE RÉGULATION CHEZ LES PROCARYOTES : L OPÉRON LACTOSE (5) Les bactéries évitent de synthétiser les enzymes d une voie métabolique en l absence des substrats adéquats Elles doivent être prêtes à tout moment à produire les enzymes si le substrat réapparaît
85 EXEMPLE DE RÉGULATION CHEZ LES PROCARYOTES : L OPÉRON LACTOSE (6) La voie du lactose fonctionne par induction Inducteur : β-galactoside Empêche au répresseur de se fixer sur Olac (l inactive) è production de la β-galactosidase (lacz) è métabolisme des β-galactosides Bactéries dans milieu sans β-galactoside: répresseur actif sur opérateur
86 EXEMPLE DE RÉGULATION CHEZ LES PROCARYOTES : L OPÉRON LACTOSE (7) Sur le répresseur 1 site de fixation à l ADN 1 site de fixation à l inducteur
87 VI-3.- EXPRESSION DU GÉNOME TRANSCRIPTION CHEZ LES EUCARYOTES
88 VI-3.- EXPRESSION DU GÉNOME TRANSCRIPTION CHEZ LES EUCARYOTES A- LES PRINCIPAUX ACTEURS
89 A) LES ARN POLYMÉRASES Trois ARN polymérases différentes : - ARN polymérase I qui transcrit l ARNr dont l activité est prédominante - ARN polymérase II qui transcrit l ARNm : elle transcrit les gènes structuraux et synthétise des produits très divers. - - ARN polymérase III qui transcrit l ARNt et d autres petits ARNs
90 A) LES ARN POLYMÉRASES (2) Les trois enzymes ont des structures similaires avec deux grandes sous-unités et de nombreuses petites sous-unités (8 à 14 sous-unités); elles ont quelques sous-unités communes.
91 A) LES ARN POLYMÉRASES (3) Aucune d elles ne reconnaît directement le promoteur comme c est le cas chez les procaryotes.
92 A) LES ARN POLYMÉRASES (4) Ce sont les facteurs de transcription qui reconnaissent les séquences caractéristiques d un promoteur particulier. De plus, ils servent à leur tour à fixer l ARN polymérase et à la positionner correctement au niveau du point de départ.
93 B) LES PROMOTEURS Les ARN polymérase I et III reconnaissent chacune un groupe de promoteurs relativement restreints et sont assistées par un petit nombre de facteurs auxiliaires..
94 B ) LE PROMOTEUR DE L ARN POLYMÉRASE II Pour l ARN polymérase II, les éléments de séquence reconnus par les facteurs de transcription sont situés en amont du point de départ. Ces sites agissent en cis et sont généralement dispersés dans une région qui comporte 200 pb. Ce sont la séquence et la distance entre ces différents éléments qui sont importantes..
95 LES SÉQUENCES PROMOTRICES DE L ARN POLYMÉRASES II Initiateur ou Inr : Py 2 CApy 5 entre les positions 3 et +6 (la première base de l ARNm est toujours un A)
96 LES SÉQUENCES PROMOTRICES DE L ARN POLYMÉRASES II TATA box : -25 en amont. Chez tous les eucaryotes. : séquence de 8 pb formée exclusivement de paires de bases A-T. Entourée des régions riches en paires G-C. Elle est proche de la séquence 10 des promoteurs bactériens.
97 LES SÉQUENCES PROMOTRICES DE L ARN POLYMÉRASES II (2) TATA box : -25 en amont. sur ce site, se fixe la protéine TBP (protéine de liaison à la TATA box = TATA binding protein). Le TBP est nécessaire à l initiation par les trois ARN polymérases. Tous les promoteurs n ont pas une TATA box.
98 LES SÉQUENCES PROMOTRICES DE L ARN POLYMÉRASES II (2) Inr et TATA box sont responsables de la sélection du point de départ exact des promoteurs de l ARN polymérase II.
99 LES SÉQUENCES PROMOTRICES DE L ARN POLYMÉRASES II (3) L efficacité et la spécificité avec lesquelles un promoteur est reconnu dépendent de courtes séquences situées plus en amont (environ 100 pb en amont du point de départ): la CAAT box en 75 è efficacité du promoteur la GC box en 90 contient la séquence GGGCGG
100 LES SÉQUENCES PROMOTRICES DE L ARN POLYMÉRASES II (4) Ces différentes boîtes diffèrent dans chaque promoteur par leur positionnement, leur nombre, leur orientation.
101 C) LES FACTEURS DE TRANSCRIPTION Ce sont des protéines Certains de ces facteurs sont courants, d autres sont spécifiques. On les regroupe en 3 catégories
102 C) LES FACTEURS DE TRANSCRIPTION (2) Facteurs généraux (TFIIX) nécessaires à l initiation de la synthèse d ARN au niveau de tous les promoteurs è appareil de transcription élémentaire
103 C) LES FACTEURS DE TRANSCRIPTION (3) Facteurs en amont : protéines dont l expression n est pas régulée. Augmentent l efficacité de l initiation è niveau convenable de fonctionnement du promoteur.
104 C) LES FACTEURS DE TRANSCRIPTION (4) Facteurs inductibles ont un rôle régulateur. Sont synthétisés ou activés à des moments spécifiques ou dans des tissus particuliers. Se fixent sur des éléments de réponse ou RE.
105 C) LES FACTEURS DE TRANSCRIPTION (5) Contrairement aux procaryotes, l initiation de la transcription chez les eucaryotes nécessite la fixation d un grand nombre de facteurs qui se fixent à différentes séquences agissant en cis.
106 Le promoteur est défini comme la région contenant tous ces sites de liaison et capable d assurer à la transcription son efficacité normale et un contrôle adéquat.
107 L ARN polymérase ne se lie pas à ces sites, elle se fixe à proximité du point de départ. En général, mode de régulation positif.
108 Une unité de transcription eucaryote contient généralement un gène et la terminaison a lieu après la fin de la région codante. Le mode de terminaison de l ARN polymérase II n est pas clair. Les ARN polymérases I et III terminent la transcription au niveau de séquences précises par des réactions bien définies
109 D) AUTRES SÉQUENCES AGISSANT EN CIS Les promoteurs peuvent être stimulés par des enhancers (séquences activatrices) qui sont des séquences qui sont définies par le fait qu elles :
110 AUTRES SÉQUENCES AGISSANT EN CIS augmentent considérablement le taux de transcription du gène auquel elles sont associées ; leur effet est indépendant de leur orientation dans le gène, de leur position ou de leur distance : elles peuvent être localisées en 5, en 3, dans un intron ou dans une région non traduite du génome ou du gène ;
111 D) AUTRES SÉQUENCES AGISSANT EN CIS (3) Elles gardent leur caractère activateur lorsqu on les déplace, leur effet étant maximal à un point donné En général, S étendent sur 200 pb Sous une forme de chromatine active (sensibilité à l ADNaseI) Entourée de régions d ADN typiques Leur mécanisme d action est en cours d élucidation Forment une boucle qui les rapproche du promoteur Seraient transcrites en un ARN qui ne serait pas traduit en protéine (earn) Stabilisation de l interaction enhancer/promoteur par cet earn (Kolovos et al, 2012)
112 D) AUTRES SÉQUENCES AGISSANT EN CIS (4). Des séquences identiques mais à effet opposé ont été mises en évidence : elles sont appelées silencers ou séquences extinctrices.
113 VI-3.- EXPRESSION DU GÉNOME TRANSCRIPTION CHEZ LES EUCARYOTES B- L ARN MESSAGER DES EUCARYOTES
114 Chez les eucaryotes supérieurs et parfois chez les eucaryotes inférieurs, les gènes sont morcelés. Le transcrit primaire a la même organisation que le gène, il est parfois appelé préarnm. Il va subir une maturation.
115 A) LE TRANSCRIT REÇOIT UNE COIFFE À L EXTRÉMITÉ 5 Le premier nucléotide (le plus souvent A) du transcrit a son groupement 5 triphosphate 5 pppapnpnpn. La guanylyl transférase è addition de G à l extrémité 5 selon une orientation inverse / autres nucléotides La coiffe est méthylée
116 A) LE TRANSCRIT REÇOIT UNE COIFFE À L EXTRÉMITÉ 5 (2) 5 Gppp + 5 pppapnpnp 5 _5 GpppApNpNp + pp + p Coiffe (cap)

117 Structure de la coiffe d'un ARN. Ce nucléotide à guanine est relié à la chaîne nucléotidique par une liaison inhabituelle : au lieu d'être reliés par une liaison ester-phosphorique entre le groupement OH porté par le carbone 3' du ribose du nucléotide à guanine et l'acide phosphorique alpha du premier nucléotide de l'arn natif, les deux nucléotides sont reliés par une liaison entre les acides phosphoriques des deux nucléotides. Par la suite, le cycle imidazole de la guanine terminale est méthylé sur son azote 7. Par ailleurs, le ribose du premier nucléotide de l'arn natif est méthylé sur l'oxygène porté par le carbone 2'. Ce peut également être le cas du nucléotide suivant. Document de Françoise Ibarrondo, Gilles Camus
118 B) POLYADÉNYLATION À L EXTRÉMITÉ 3 La plupart des ARNm des eucaryotes Le préarnm subit un clivage à son extrémité 3 avant la polyadénylation Ce clivage se fait au niveau d un site précis : le dinucléotide CA qui est entouré de deux autres séquences : AAUAAA très conservée située 10 à 30 nucléotides en amont du site de clivage (à ne pas retenir), La séquence DES riche en U ou en GU situé une trentaine de nucléotides en aval du site de clivage (à ne pas retenir),
119 B) POLYADÉNYLATION À L EXTRÉMITÉ 3 Le clivage est assuré par un complexe de clivage composé de plusieurs protéines Après clivage, la poly (A) polymérase ajoute environ 200 nucléotides (= queue poly(a)) Les ARNm qui portent une queue polya sont dits ARNm poly(a) +
120 B) POLYADÉNYLATION À L EXTRÉMITÉ 3 (2) Fixation de la protéine de liaison aux poly(a) (PABP) Rôle des poly(a) Stabilisation ARNm? Favoriserait la traduction?
121 C) L ÉPISSAGE DU TRANSCRIT PRIMAIRE Le processus par lequel les introns du préarnm sont enlevés et les exons resoudés est appelé épissage A lieu dans le noyau
122 C) L ÉPISSAGE DU TRANSCRIT PRIMAIRE (2) Les sites d épissage (les 2 frontières exonintron) Site gauche ou donneur Site droit (3 ) (5 ) Exon AG GUAAGU 12PyNCAG N Exon Intron : règle GU AG
123 C) L ÉPISSAGE DU TRANSCRIT PRIMAIRE (3) 1- coupure au niveau du site 5 2- structure en lasso de la molécule intron-exon de droite : extrémité 5 produite est reliée à une base dans l intron : base A appartenant à une séquence de branchement
124 C) L ÉPISSAGE DU TRANSCRIT PRIMAIRE (4) 3- coupure au niveau du site 3 è libération de l intron sous la forme d un lasso et ligature des 2 exons 4- débranchement du lasso è intron linéaire excisé

125
126 C) L ÉPISSAGE DU TRANSCRIT PRIMAIRE (5) Epissage réalisé par un gros complexe : particule d épissage (ou spliceosome) composé de petits ARNs et de nucléoprotéines
127 L ARN est ensuite transporté à travers les pores nucléaires vers le cytoplasme où il est reconnu par les ribosomes et traduit. Les demi-vies des ARNms des cellules animales varient entre 4 et 24 heures.
128 Les ARNm des eucaryotes existent sous une forme associée à des protéines : les ribonucléoprotéines. (RNPsm).
129 VI-4.- EXPRESSION DU GÉNOME LES ARN DE TRANSFERT
130 ARN DE TRANSFERT OU ARNT Chez les procaryotes, le transcrit primaire peut être clivé pour donner des produits matures : les ARNr et les ARNt. Chez les eucaryotes, ces ARNt sont transcrits à partir de gènes spécifiques et à l aide de l ARN polymérase III.
131 ARN DE TRANSFERT OU ARNT (2) L ARNt est l adaptateur entre l ARNm et les acides aminés lors de la traduction. C est une petite molécule de 75 à 85 bases de long.
132 ARN DE TRANSFERT OU ARNT (3) Il a deux propriétés essentielles : il représente un seul acide aminé auquel il se lie par covalence il contient une séquence de trois nucléotides, l anticodon, qui est complémentaire du codon représentant son acide aminé. L anticodon permet à l ARNt de reconnaître le codon grâce à l appariement des bases complémentaires.
133 ARN DE TRANSFERT OU ARNT (4) La structure secondaire de l ARNt peut être représentée sous la forme d une feuille de trèfle dans laquelle l appariement de bases complémentaires conduit à la formation de tiges ayant à leur extrémité des boucles simple brin.
134 ARN DE TRANSFERT OU ARNT (5) Ces structures constituent les bras ou boucles de l ARNt : de 3 vers 5 bras accepteur (site de fixation de l acide aminé), bras TψC (ψ = pseudouridine), bras supplémentaire ou bras variable. bras de l anticodon bras D (contient de la dihydrouridine),

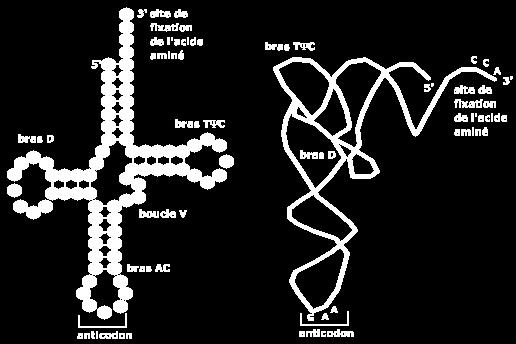
135
136 ARN DE TRANSFERT OU ARNT (6) Les acides aminés sont fixés sur l ARNt correspondant par l aminoacyl-arnt synthétase (20 différentes). L ARNt chargé de son acide aminé est appelé aminoacyl-arnt.
137 VI-5.- EXPRESSION DU GÉNOME LES ARNS RIBOSOMAUX
138 A) ARNR CHEZ LES PROCARYOTES Il y a 7 opérons non contigus codant l ARNr de E. coli : opérons rrna-g. ayant la même organisation : ils contiennent tous trois molécules d ARNr dans l ordre : 16S-23S-5S dans une zone intermédiaire une séquence codant pour un ARNt spécifique
139 A) ARNR CHEZ LES PROCARYOTES (2) Ces opérons sont transcrits à partir du même ARN précurseur, sédimentant à 30S, Clivage précurseur è : les différentes molécules d ARNr un ARNt enzyme impliquée : ARNase III. Maturation des produits de clivage par l ARNase III
140 B) ARNR CHEZ LES EUCARYOTES Mieux connus chez les mammifères. Le transcrit primaire chez les mammifères est un ARN 45S fortement méthylé, contenant les séquences des ARNrs 18S et 28S et du petit ARN 5,8S.
141 B) ARNR CHEZ LES EUCARYOTES (2) La méthylation servirait à distinguer les régions du transcrit primaire destinées à subir la maturation en ARNr. Le transcrit subit sa maturation alors qu il est associé à des ribonucléoprotéines.
142 VI-6.- EXPRESSION DU GÉNOME LA TRADUCTION
143 VI-6.- EXPRESSION DU GÉNOME LA TRADUCTION A- LE CODE GÉNÉTIQUE
144 LE CODE GÉNÉTIQUE Le code génétique est déchiffré par un système complexe situé entre acides nucléiques et protéines Au niveau de l ADN, il est lu par groupe de trois nucléotides ou codon. A un codon correspond un acide aminé
145 LE CODE GÉNÉTIQUE (2) Chaque gène = suite de codons lus séquentiellement d un point de départ situé à une extrémité du gène jusqu à un point de terminaison situé à l autre extrémité Crick et coll., 1961 : le code génétique est lu sous la forme de triplets non chevauchants
146 LE CODE GÉNÉTIQUE (3) Triplets non chevauchants = trois façons de traduire une même séquence nucléotidique en protéine selon le point de départ : les cadres de lecture
147 LE CODE GÉNÉTIQUE (4) CADRES DE LECTURES Une séquence : A C G A C G A C G A C G A C G A C G Trois cadres de lecture possibles ACG ACG ACG ACG ACG CGA CGA CGA CGA CGA GAC GAC GAC GAC GAC
148 LE CODE GÉNÉTIQUE (5) CADRES DE LECTURES Les mutations qui insèrent ou enlèvent une base è changement du cadre de lecture en aval de la mutation è décalage du cadre de lecture Conséquence de ces mutations : séquence différente d acides aminés = protéine différente è perte probable de la fonction de la protéine
149 LE CODE GÉNÉTIQUE (6) 64 codons possibles (4 3 ) 61 codons correspondent à des acides aminés 3 entraînent l arrêt de la synthèse protéique L ARNt est l intermédiaire qui traduit chaque triplet de nucléotides en un acide aminé
150 LE CODE GÉNÉTIQUE (7) 64 codons et 20 acides aminés : chaque acide aminé est représenté par plusieurs codons sauf la méthionine et le tryptophane Les codons représentant le même acide aminé diffèrent en général par la troisième base : le code génétique est dégénéré

151
152 LE CODE GÉNÉTIQUE (8) Le code génétique est universel : Pas de changement dans les génomes nucléaires Mais changement au cours de l évolution dans le génome mitochondrial
153 VI-6.- EXPRESSION DU GÉNOME LA TRADUCTION B- LES ACTEURS DE LA TRADUCTION
154 LES ACTEURS DE LA TRADUCTION L ARNm : codons Les aminoacyl-arnts avec les anticodons Les différentes enzymes Les ribosomes Les facteurs auxiliaires qui assistent le ribosome à chaque étape GTP : son hydrolyse fournit l énergie
155 LES RIBOSOMES Particules ribonucléoprotéiques : Protéines ARNr La plupart de ces protéines et ARNr ont un rôle structural strict et ne sont pas impliqués dans la synthèse des protéines
156 LES RIBOSOMES (2) Protéines ribosomales : protéines r Les ribosomes sont bien caractérisés chez les procaryotes Tous les ribosomes d une même bactérie sont identiques
157 LES RIBOSOMES (3) CHEZ E. COLI Ribosome complet (66% d ARNs) : densité de 70S Petite sous-unité de 30 S 1 ARNr de 16S 21 protéines Grande sous-unité de 50S 1 ARNr de 23S un petit ARN 5S 31 protéines
158 LES RIBOSOMES (4) CHEZ LES EUCARYOTES SUPÉRIEURS Ribosome complet (60% d ARNs) : densité de 80S Petite sous-unité de 40 S 1 ARNr de 18S 33 protéines Grande sous-unité de 60S 1 ARNr de 28S un petit ARN 5S Un petit ARN de 5,8 S 49 protéines
159 LES RIBOSOMES (5) Se comportent comme un facteur mobile qui se déplace le long d une matrice Fournit l environnement qui permet de contrôler l interaction ARNm / Aminoacyl- ARNt
160 LES RIBOSOMES (6) Différents sites actifs : Site P ou site donneur (site polypeptidique) à cheval sur les 2 sous-unités Site A : site accepteur (de l aminoacyl-arnt) à cheval sur les 2 sous-unités Site E : site par lequel l ARNt déchargé de son acide aminé quitte le ribosome
161 LES RIBOSOMES (6) Différents sites actifs : Site peptidyl transférase Un domaine de sortie du peptide Site de liaison de l ARNm
162 VI-6.- EXPRESSION DU GÉNOME LA TRADUCTION C- LES ÉTAPES DE LA TRADUCTION
163 L ARNm est associé à la petite sous-unité sur environ 30 bases à tout moment A tout moment, 2 molécules d ARNt sont actives dans la synthèse protéique Ces 2 molécules d ARNt s insèrent au niveau de sites internes du ribosome
164 Un aminoacyl-arnt entrant se fixe sur le site A Avant l entrée de l aminoacyl-arnt, le site expose le codon correspondant au prochain acide aminé qui devra être ajouté à la chaîne polypeptidique
165 Le codon représentant l acide aminé qui vient juste d être fixé à la chaîne polypeptidique se trouve dans le site P. Ce site est occupé par un peptidyl-arnt : un ARNt qui porte la chaîne polypeptidique naissante
166 LES DIFFÉRENTS ÉTAPES L initiation L élongation La terminaison
167 L INITIATION Met en jeu les réactions qui précèdent la formation de la liaison peptidique entre les deux premiers acides aminés de la chaine peptidique Nécessite la fixation du ribosome à l ARNm è complexe d initiation qui contient le premier aminoacyl-arnt Etape lente
168 L INITIATION (2) L initiation se produit au niveau d une séquence particulière de l ARNm : site de liaison des ribosomes
169 L INITIATION (3) Ces sites de liaison de 30 bases environ ont 2 caractéristiques : Le codon initiateur AUG (plus rarement GUG ou UUG chez les bactéries) est toujours à l intérieur du site de liaison A moins de 10 bases en amont de l AUG, une séquence qui correspond totalement ou en partie à l hexamère 5..A G G A G G..3 : séquence de Shine-Dalgarno (complémentaire de l extrémité 3 de l ARNr 16S)
170 L INITIATION (4) La reconnaissance de l ARNm : fixation d une petite sous-unité sur l ARNm è complexe d initiation : nécessité de facteurs d initiation (IFs) Libération des IFs è Puis une grande sous-unité rejoint le complexe è ribosome complet Bactéries : IF-1, 2 et 3.
171 L INITIATION (5) La synthèse de toutes les protéines débute par la méthionine Signal d initiation = codon d initiation : AUG, (GUG, UUG plus rarement chez les bactéries) Deux types d ARNt peuvent porter la méthionine selon qu elle soit initiatrice ou localisée à l intérieur de la chaine peptidique
172 L INITIATION (6) Chez les bactéries et dans les organites eucaryotes, l ARNt initiateur porte un résidu méthionine formylé sur son groupement amine è molécule de N-formyl-méthionyl- ARNt (ARNt Met f ou fmet-arnt) Chez les eucaryotes, l aminoacylarnt initiateur n est pas formylé, il contient d autres modifications qui permettent de le distinguer des met-arnt intrachaine Il peut être intégré directement au site P
173 L INITIATION (7) Chez les eucaryotes, la petite sous-unité reconnaît un ARNm grâce à sa coiffe en 5, migre jusqu à l AUG où elle sera rejointe par la grande sous-unité
174 L INITIATION (8) Lorsque la grande sous-unité rejoint le complexe, l aminoacyl-arnt initiateur se trouve dans le site P Le site A est disponible pour l entrée de l aminoacyl-arnt complémentaire du deuxième codon du gène
175 L ÉLONGATION Comprend toutes les réactions entre la synthèse de la première liaison peptidique et l addition du dernier acide aminé Les acides aminés sont ajoutés un à un à la chaîne polypeptidique Ajout d un acide aminé : étape très rapide
176 L ÉLONGATION (2) Le ribosome est complet au niveau du site d initiation Démarrage d un cycle : 1 aminoacyl-arnt entre dans le site A grâce à un facteur d élongation (EF- Tu chez les bactéries, eef-1α chez les eucaryotes) en présence de GTP GTP permet la liaison au site A N importe quel aminoacyl-arnt peut entrer dans le site A
177 L ÉLONGATION (3) Démarrage d un cycle : Le site P est occupé soit Soit par l aminoacyl-arnt initiateur Soit par un peptidyl-arnt
178 L ÉLONGATION (4) Transfert du polypeptide ou de de la méthionine formylée attaché à l ARNt du site P sur l aminoacyl-arnt présent dans le site A Synthèse de la liaison peptidique par l activité peptidyl transférase de la grande sous-unité ribosomale
179 L ÉLONGATION (5) Puis survient la translocation du ribosome qui se déplace de trois nucléotides sur l ARNm è expulsion du site P de l ARNt non chargé è entrée d un nouveau peptidyl-arnt è site A vide è positionnement de l aminoacyl-arnt correspondant au codon suivant Nécessité du GTP Facteur d élongation EF-G (eef-2 chez les eucaryotes)
180 LA TERMINAISON Englobe les réactions nécessaires pour libérer la chaîne polypeptidique complète; Dans le même temps le ribosome se dissocie de l ARNm
181 LA TERMINAISON (2) 3 codons de terminaison ou codons stop: UAG, UAA, UGA Pas d ARNt correspondant Ces codons sont reconnus directement par des facteurs protéiques : les facteurs de libération (RF) RF-1 et 2 chez E. coli
182 LA TERMINAISON (3) Provoque la libération du polypeptide achevé L expulsion du dernier ARNt du ribosome La dissociation de l ARNm du ribosome
183 RAPPEL : LE CODE GÉNÉTIQUE CADRES DE LECTURES Une séquence : A C G A C G A C G A C G A C G A C G Trois cadres de lecture possibles ACG ACG ACG ACG ACG CGA CGA CGA CGA CGA GAC GAC GAC GAC GAC
184 CADRE DE LECTURE OUVERT Cadre de lecture qui commence par le codon AUG et se poursuit en triplets représentant des acides aminés jusqu au codon de terminaisonè correspond à une séquence traduite en protéine ORF en anglais
185 CADRE DE LECTURE OUVERT (2) En général les deux autres cadres de lecture sont bloqués par de fréquents codons stop è ne synthétisent pas de protéines
186 MODIFICATIONS POST TRADUCTIONNELLES DES PROTÉINES Peuvent être multiples qualitativement et/ou quantitativement dans une même molécule : Glycosylations Phosphorylation Acétylation
187 VI-7.- EXPRESSION DU GÉNOME LA RÉGULATION DE L EXPRESSION DES GÈNES CHEZ LES EUCARYOTES (ARN POLYMÉRASE II)
188 Cinq points de contrôle potentiels Activation de la structure d un gène Initiation de la transcription Maturation du transcrit Transport vers le cytoplasme Traduction de l ARNm
189 VI-7.- EXPRESSION DU GÉNOME LA RÉGULATION DE L EXPRESSION DES GÈNES CHEZ LES EUCARYOTES (ARN POLYMÉRASE II) A- ACTIVATION DE LA STRUCTURE DU GÈNE
190 ACTIVATION DE LA STRUCTURE DU GÈNE Les gènes peuvent exister dans deux états structuraux : actif inactif Les gènes sont à l état actif dans les cellules qui les expriment Le changement de structure indique que le gène est susceptible d être transcrit L acquisition de la structure active est la première étape incontournable de l expression des gènes
191 ACTIVATION DE LA STRUCTURE DU GÈNE (2) Activation du gène par : Exclusion des nucléosomes Niveau de méthylation
192 ACTIVATION DE LA STRUCTURE DU GÈNE : EXCLUSION DES NUCLÉOSOMES Il y aurait une compétition entre les histones (octamères d histones ou nucléosomes) et les facteurs de transcription pour se fixer sur l ADN Sites sensibles à l ADNase I: Régions où nucléosomes exclus par la présence d autres protéines se liant à l ADN (ADN ou ARN polymérases, facteurs transcriptionnels, ) Régions hypersensibles : en général se situent dans les régions 5 non transcrites des gènes : promoteurs des gènes actifs
193 ACTIVATION DE LA STRUCTURE DU GÈNE : LA MÉTHYLATION DE L ADN CHEZ LES EUCARYOTES Chez les eucaryotes, la méthylation de l ADN est principalement liée au contrôle de la transcription 2 à 7% des cytosines de l ADN des cellules animales sont méthylées La plupart des groupements méthyle sont associés à des dinucléotides CG (ilots CpG) 5 m CpG 3 3 GpC m 5 Le gène méthylé est inactif Le gène non méthylé ou sous-méthylé est actif
194 ACTIVATION DE LA STRUCTURE DU GÈNE : LA MÉTHYLATION DE L ADN CHEZ LES EUCARYOTES (2) L ADN subit des épisodes de déméthylation globale aux stades précoces du développement Surviendraient essentiellement au cours de 2 étapes clés du développement : dans les cellules germinales et chez l embryon avant l implantation Rôle : maintien de la totipotence des gènes = tous les gènes fonctionnent Ces déméthylations sont suivies d une méthylation de novo par des ADN méthyltranférases Ensuite, dans les différents tissus, seuls les gènes tissu-spécifiques subissent des déméthylations et vont s exprimer Tan et Shi, 2012; (R Chatterjee, and C Vinson, 2012;, Jeffrey R. Shearstone et al, 2011;
195 VI-7.- EXPRESSION DU GÉNOME LA RÉGULATION DE L EXPRESSION DES GÈNES CHEZ LES EUCARYOTES (ARN POLYMÉRASE II) B- RÉGULATION DE L INITIATION DE LA TRANSCRIPTION
196 LA RÉGULATION EN CIS Assurée par des séquences influençant l initiation et le taux de la transcription Eléments à localisation fixe : les promoteurs Eléments à localisation variables : les séquences stimulatrices (enhancers) et les séquences extinctrices (silencers)
197 LA RÉGULATION EN CIS (2) Ces séquences sont reconnues par des protéines spécifiques Régulation positive : protéine présente uniquement si le gène doit être exprimé
198 LA RÉGULATION EN CIS (3) Promoteurs ou enhancers Certaines séquences confèrent une spécificité tissulaire d expression (= gènes exprimés spécifiquement dans certains tissus) D autres sont la cible de facteurs transcriptionnels sous le contrôle de stimuli extra ou intracellulaires (hormones, ions, choc thermique,..) Ce sont les Eléments de réponse (Response element ou RE)
199 LA RÉGULATION EN CIS (4) Exemples de RE GRE : élément de réponse aux glucocorticoïdes IRE : élément de réponse au fer
200 LA RÉGULATION EN TRANS : LES FACTEURS TRANSCRIPTIONNELS Très nombreux Contiennent : Un domaine de fixation à l ADN : différents types Un domaine d activation de la transcription = domaine de liaison à d autres protéines (ARN polymérase,..)
201 LA RÉGULATION EN TRANS : LES FACTEURS TRANSCRIPTIONNELS (2) Rôle ++++ de la disposition relative dans l espace des différentes protéines (plus importante que leur distance relative par rapport au promoteur) Rôle de la structure dans l espace et des interactions protéines-protéines dans la régulation
202 VI-7.- EXPRESSION DU GÉNOME LA RÉGULATION DE L EXPRESSION DES GÈNES CHEZ LES EUCARYOTES (ARN POLYMÉRASE II) C- RÉGULATION POSTTRANSCRIPTIONNELLE
203 EPISSAGE ALTERNATIF Gène Exon 1 Exon 2 Exon 3 Exon 4 Transcription Transcrit Prim Exon 1 Exon 2 Exon 3 ARNm A Protéine A Exon 1 Exon 2 Exon 4 ARNm B Protéine B
204 EPISSAGE ALTERNATIF (2) En fonction des conditions ou des besoins, la protéine A ou la protéine B sera synthétisée: Certains cas, fonction du sexe D autres cas, fonction des tissus
205 STABILISATION DE L ARNM : EXEMPLE DE LA TRANSFERRINE Transferrine : molécule qui transporte le fer L ARNm de la transferrine contient une séquence appelée IRE IRE : région 3 non codante de l ARNm Fixation d une protéine avec une affinité contrôlée par la concentration en fer Présence de fer è stabilisation de l ARNm
CHAPITRE 3 LA SYNTHESE DES PROTEINES
 CHAITRE 3 LA SYNTHESE DES ROTEINES On sait qu un gène détient dans sa séquence nucléotidique, l information permettant la synthèse d un polypeptide. Ce dernier caractérisé par sa séquence d acides aminés
CHAITRE 3 LA SYNTHESE DES ROTEINES On sait qu un gène détient dans sa séquence nucléotidique, l information permettant la synthèse d un polypeptide. Ce dernier caractérisé par sa séquence d acides aminés
Module 5 La maturation de l ARN et le contrôle post-transcriptionnel chez les eucaryotes
 Module 5 La maturation de l ARN et le contrôle post-transcriptionnel chez les eucaryotes Où trouver l'information complémentaire? MCB -11, GVII-5, 22, 23. La maturation des ARNm chez les eucaryotes Les
Module 5 La maturation de l ARN et le contrôle post-transcriptionnel chez les eucaryotes Où trouver l'information complémentaire? MCB -11, GVII-5, 22, 23. La maturation des ARNm chez les eucaryotes Les
Contrôle de l'expression génétique : Les régulations post-transcriptionnelles
 Contrôle de l'expression génétique : Les régulations post-transcriptionnelles http://perso.univ-rennes1.fr/serge.hardy/ utilisateur : biochimie mot de passe : 2007 L'ARNm, simple intermédiaire entre le
Contrôle de l'expression génétique : Les régulations post-transcriptionnelles http://perso.univ-rennes1.fr/serge.hardy/ utilisateur : biochimie mot de passe : 2007 L'ARNm, simple intermédiaire entre le
ULBI 101 Biologie Cellulaire L1. Le Système Membranaire Interne
 ULBI 101 Biologie Cellulaire L1 Le Système Membranaire Interne De la nécessité d un SMI Le volume augmente comme le cube de la dimension linéaire, alors que la surface n'est augmentée que du carré Une
ULBI 101 Biologie Cellulaire L1 Le Système Membranaire Interne De la nécessité d un SMI Le volume augmente comme le cube de la dimension linéaire, alors que la surface n'est augmentée que du carré Une
TD de Biochimie 4 : Coloration.
 TD de Biochimie 4 : Coloration. Synthèse de l expérience 2 Les questions posées durant l expérience 2 Exposé sur les méthodes de coloration des molécules : Générique Spécifique Autres Questions Pourquoi
TD de Biochimie 4 : Coloration. Synthèse de l expérience 2 Les questions posées durant l expérience 2 Exposé sur les méthodes de coloration des molécules : Générique Spécifique Autres Questions Pourquoi
Contrôle de l'expression génétique :
 Contrôle de l'expression génétique : Les régulations post-transcriptionnelles L'ARNm, simple intermédiaire entre le génome et les protéines? gène protéine L'ARNm, simple intermédiaire entre le génome et
Contrôle de l'expression génétique : Les régulations post-transcriptionnelles L'ARNm, simple intermédiaire entre le génome et les protéines? gène protéine L'ARNm, simple intermédiaire entre le génome et
Séquence 2. L expression du patrimoine génétique. Sommaire
 Séquence 2 L expression du patrimoine génétique Sommaire 1. La synthèse des protéines 2. Phénotypes, génotypes et environnement Synthèse de la séquence 2 Exercices de la séquence 2 Glossaire des séquences
Séquence 2 L expression du patrimoine génétique Sommaire 1. La synthèse des protéines 2. Phénotypes, génotypes et environnement Synthèse de la séquence 2 Exercices de la séquence 2 Glossaire des séquences
Travaux dirigés de Microbiologie Master I Sciences des Génomes et des Organismes Janvier 2015
 Andrew Tolonen [email protected] Travaux dirigés de Microbiologie Master I Sciences des Génomes et des Organismes Janvier 2015 A- Généralités I- La vie sur terre telle que nous la connaissons ne
Andrew Tolonen [email protected] Travaux dirigés de Microbiologie Master I Sciences des Génomes et des Organismes Janvier 2015 A- Généralités I- La vie sur terre telle que nous la connaissons ne
Chapitre 7 : Structure de la cellule Le noyau cellulaire
 UE2 : Structure générale de la cellule Chapitre 7 : Structure de la cellule Le noyau cellulaire Professeur Michel SEVE Année universitaire 2010/2011 Université Joseph Fourier de Grenoble - Tous droits
UE2 : Structure générale de la cellule Chapitre 7 : Structure de la cellule Le noyau cellulaire Professeur Michel SEVE Année universitaire 2010/2011 Université Joseph Fourier de Grenoble - Tous droits
Dr E. CHEVRET UE2.1 2013-2014. Aperçu général sur l architecture et les fonctions cellulaires
 Aperçu général sur l architecture et les fonctions cellulaires I. Introduction II. Les microscopes 1. Le microscope optique 2. Le microscope à fluorescence 3. Le microscope confocal 4. Le microscope électronique
Aperçu général sur l architecture et les fonctions cellulaires I. Introduction II. Les microscopes 1. Le microscope optique 2. Le microscope à fluorescence 3. Le microscope confocal 4. Le microscope électronique
Tableau 1. Liste (non exhaustive) des protéines se localisant dans les P-Bodies
 des protéines se localisant dans les P-Bodies") NOM FONCTION EFFET DE L ABSENCE OU DE REFERENCES LA SUREXPRESSION SUR LES P- BODIES XRN1 exonucléase 5 3 Absence : augmente la taille et le Bashkirov et al. 1997 Shet et Parker 2003 Cougot nombre des P-bodies
NOM FONCTION EFFET DE L ABSENCE OU DE REFERENCES LA SUREXPRESSION SUR LES P- BODIES XRN1 exonucléase 5 3 Absence : augmente la taille et le Bashkirov et al. 1997 Shet et Parker 2003 Cougot nombre des P-bodies
Université d Evry-Val d Essonne Ecole Doctorale des Génomes Aux Organismes. Thèse
 Université d Evry-Val d Essonne Ecole Doctorale des Génomes Aux Organismes Thèse Présentée pour obtenir le grade de Docteur en sciences de l université d Evry-Val d Essonne Spécialité Bioinformatique par
Université d Evry-Val d Essonne Ecole Doctorale des Génomes Aux Organismes Thèse Présentée pour obtenir le grade de Docteur en sciences de l université d Evry-Val d Essonne Spécialité Bioinformatique par
Univers Vivant Révision. Notions STE
 Univers Vivant Révision Notions STE Chap. 13) L Écologie 1) a) Qu est-ce que l empreinte écologique? L empreinte écologique correspond à la surface terrestre et aquatique totale nécessaire à un individu,
Univers Vivant Révision Notions STE Chap. 13) L Écologie 1) a) Qu est-ce que l empreinte écologique? L empreinte écologique correspond à la surface terrestre et aquatique totale nécessaire à un individu,
IMMUNOLOGIE. La spécificité des immunoglobulines et des récepteurs T. Informations scientifiques
 IMMUNOLOGIE La spécificité des immunoglobulines et des récepteurs T Informations scientifiques L infection par le VIH entraîne des réactions immunitaires de l organisme qui se traduisent par la production
IMMUNOLOGIE La spécificité des immunoglobulines et des récepteurs T Informations scientifiques L infection par le VIH entraîne des réactions immunitaires de l organisme qui se traduisent par la production
Biomarqueurs en Cancérologie
 Biomarqueurs en Cancérologie Définition, détermination, usage Biomarqueurs et Cancer: définition Anomalie(s) quantitative(s) ou qualitative(s) Indicative(s) ou caractéristique(s) d un cancer ou de certaines
Biomarqueurs en Cancérologie Définition, détermination, usage Biomarqueurs et Cancer: définition Anomalie(s) quantitative(s) ou qualitative(s) Indicative(s) ou caractéristique(s) d un cancer ou de certaines
ACIDES BASES. Chap.5 SPIESS
 ACIDES BASES «Je ne crois pas que l on me conteste que l acide n ait des pointes Il ne faut que le goûter pour tomber dans ce sentiment car il fait des picotements sur la langue.» Notion d activité et
ACIDES BASES «Je ne crois pas que l on me conteste que l acide n ait des pointes Il ne faut que le goûter pour tomber dans ce sentiment car il fait des picotements sur la langue.» Notion d activité et
Les OGM. 5 décembre 2008. Nicole Mounier
 Les OGM 5 décembre 2008 Nicole Mounier Université Claude Bernard Lyon 1 CGMC, bâtiment Gregor Mendel 43, boulevard du 11 Novembre 1918 69622 Villeurbanne Cedex OGM Organismes Génétiquement Modifiés Transfert
Les OGM 5 décembre 2008 Nicole Mounier Université Claude Bernard Lyon 1 CGMC, bâtiment Gregor Mendel 43, boulevard du 11 Novembre 1918 69622 Villeurbanne Cedex OGM Organismes Génétiquement Modifiés Transfert
3: Clonage d un gène dans un plasmide
 3: Clonage d un gène dans un plasmide Le clonage moléculaire est une des bases du génie génétique. Il consiste à insérer un fragment d'adn (dénommé insert) dans un vecteur approprié comme un plasmide par
3: Clonage d un gène dans un plasmide Le clonage moléculaire est une des bases du génie génétique. Il consiste à insérer un fragment d'adn (dénommé insert) dans un vecteur approprié comme un plasmide par
De la physico-chimie à la radiobiologie: nouveaux acquis (I)
") De la physico-chimie à la radiobiologie: nouveaux acquis (I) Collaboration: - Laboratoire de Radiotoxicologie et Oncologie (L. Sabatier) CEA, DSV - Laboratoire de Génotoxicité et Modulation de l Expression
De la physico-chimie à la radiobiologie: nouveaux acquis (I) Collaboration: - Laboratoire de Radiotoxicologie et Oncologie (L. Sabatier) CEA, DSV - Laboratoire de Génotoxicité et Modulation de l Expression
IlliS. Le ribosome bactérien : structure et fonctions SYNTHÈSE
 IlliS SYNTHÈSE médecine! sciences 1989 ; 5: 662-69 Le ribosome bactérien : structure et fonctions Le ribosome est une particule ribonucléoprotéique, complexe jouant le rôle d'«unité centrale» de la synthèse
IlliS SYNTHÈSE médecine! sciences 1989 ; 5: 662-69 Le ribosome bactérien : structure et fonctions Le ribosome est une particule ribonucléoprotéique, complexe jouant le rôle d'«unité centrale» de la synthèse
Chapitre II La régulation de la glycémie
 Chapitre II La régulation de la glycémie Glycémie : concentration de glucose dans le sang valeur proche de 1g/L Hypoglycémie : perte de connaissance, troubles de la vue, voire coma. Hyperglycémie chronique
Chapitre II La régulation de la glycémie Glycémie : concentration de glucose dans le sang valeur proche de 1g/L Hypoglycémie : perte de connaissance, troubles de la vue, voire coma. Hyperglycémie chronique
Cellules procaryotes Service histologie Pr.k.mebarek
 Cellules procaryotes Service histologie Pr.k.mebarek I) Les cellules procaryotes II) Les cellules eucaryotes o 1) Caractéristiques générales des cellules eucaryotes o 2) Organisation des cellules eucaryotes
Cellules procaryotes Service histologie Pr.k.mebarek I) Les cellules procaryotes II) Les cellules eucaryotes o 1) Caractéristiques générales des cellules eucaryotes o 2) Organisation des cellules eucaryotes
INTRODUCTION À L'ENZYMOLOGIE
 INTRODUCTION À L'ENZYMOLOGIE Les enzymes sont des macromolécules spécialisées qui - catalysent les réactions biologiques - transforment différentes formes d'énergie. Les enzymes diffèrent des catalyseurs
INTRODUCTION À L'ENZYMOLOGIE Les enzymes sont des macromolécules spécialisées qui - catalysent les réactions biologiques - transforment différentes formes d'énergie. Les enzymes diffèrent des catalyseurs
Université de Montréal. Développement d outils pour l analyse de données de ChIP-seq et l identification des facteurs de transcription
 Université de Montréal Développement d outils pour l analyse de données de ChIP-seq et l identification des facteurs de transcription par Eloi Mercier Département de bioinformatique Faculté de médecine
Université de Montréal Développement d outils pour l analyse de données de ChIP-seq et l identification des facteurs de transcription par Eloi Mercier Département de bioinformatique Faculté de médecine
Bases moléculaires des mutations Marc Jeanpierre
 Bases moléculaires des mutations Marc Jeanpierre Chaque enfant qui naît hérite de 10 à 30 nouvelles mutations ponctuelles. L essentiel des ces mutations sont heureusement des variations neutres de séquence
Bases moléculaires des mutations Marc Jeanpierre Chaque enfant qui naît hérite de 10 à 30 nouvelles mutations ponctuelles. L essentiel des ces mutations sont heureusement des variations neutres de séquence
MYRIAD. l ADN isolé n est à présent plus brevetable!
 MYRIAD La Cour Suprême des Etats-Unis revient sur plus de 30 ans de pratique : l ADN isolé n est à présent plus brevetable! Mauvaise passe pour les inventions en biotechnologies sur le territoire américain.
MYRIAD La Cour Suprême des Etats-Unis revient sur plus de 30 ans de pratique : l ADN isolé n est à présent plus brevetable! Mauvaise passe pour les inventions en biotechnologies sur le territoire américain.
Formavie 2010. 2 Différentes versions du format PDB...3. 3 Les champs dans les fichiers PDB...4. 4 Le champ «ATOM»...5. 6 Limites du format PDB...
 Formavie 2010 Les fichiers PDB Les fichiers PDB contiennent les informations qui vont permettre à des logiciels de visualisation moléculaire (ex : RasTop ou Jmol) d afficher les molécules. Un fichier au
Formavie 2010 Les fichiers PDB Les fichiers PDB contiennent les informations qui vont permettre à des logiciels de visualisation moléculaire (ex : RasTop ou Jmol) d afficher les molécules. Un fichier au
ORIGINE ET DIFFERENTS TYPES DE RYTHMES BIOLOGIQUES. Carine Bécamel [email protected] 04 67 14 29 83
 ORIGINE ET DIFFERENTS TYPES DE RYTHMES BIOLOGIQUES Carine Bécamel [email protected] 04 67 14 29 83 INTRODUCTION Rythmes biologiques: définition Tout être vivant (humain, animal, végétal) est soumis
ORIGINE ET DIFFERENTS TYPES DE RYTHMES BIOLOGIQUES Carine Bécamel [email protected] 04 67 14 29 83 INTRODUCTION Rythmes biologiques: définition Tout être vivant (humain, animal, végétal) est soumis
Hépatite chronique B Moyens thérapeutiques
 Hépatite chronique B Moyens thérapeutiques Dr Olfa BAHRI Laboratoire de Virologie Clinique Institut Pasteur de Tunis INTRODUCTION Plus de 300. 10 6 porteurs chroniques de VHB dans le monde Hépatite chronique
Hépatite chronique B Moyens thérapeutiques Dr Olfa BAHRI Laboratoire de Virologie Clinique Institut Pasteur de Tunis INTRODUCTION Plus de 300. 10 6 porteurs chroniques de VHB dans le monde Hépatite chronique
Conférence technique internationale de la FAO
 Décembre 2009 ABDC-10/7.2 F Conférence technique internationale de la FAO Biotechnologies agricoles dans les pays en développement: choix et perspectives pour les cultures, les forêts, l élevage, les pêches
Décembre 2009 ABDC-10/7.2 F Conférence technique internationale de la FAO Biotechnologies agricoles dans les pays en développement: choix et perspectives pour les cultures, les forêts, l élevage, les pêches
2 C est quoi la chimie?
 PARTIE 1 AVANT LA CHIMIE VERTE... 2 C est quoi la chimie? L inconnu étant source d angoisse, nous allons essayer de définir les grands domaines de la chimie pour mieux la connaître, l appréhender et donc
PARTIE 1 AVANT LA CHIMIE VERTE... 2 C est quoi la chimie? L inconnu étant source d angoisse, nous allons essayer de définir les grands domaines de la chimie pour mieux la connaître, l appréhender et donc
Annales de Biologie Cellulaire QCM (niveau SVT 1 er année)
") Annales de Biologie Cellulaire QCM (niveau SVT 1 er année) Equipe pédagogique Université Bordeaux-1 Didier Morin, Michel Moenner, Sophie North, Gérard Tramu et IJsbrand Kramer Contact : [email protected]
Annales de Biologie Cellulaire QCM (niveau SVT 1 er année) Equipe pédagogique Université Bordeaux-1 Didier Morin, Michel Moenner, Sophie North, Gérard Tramu et IJsbrand Kramer Contact : [email protected]
LA MITOSE CUEEP - USTL DÉPARTEMENT SCIENCES BAHIJA DELATTRE
 Biologie LA MITOSE CUEEP - USTL DÉPARTEMENT SCIENCES BAHIJA DELATTRE Février 2006 I. L'INTRODUCTION Chaque cellule d'un organisme supérieur provient de la multiplication d'une cellule préexistante (cellule
Biologie LA MITOSE CUEEP - USTL DÉPARTEMENT SCIENCES BAHIJA DELATTRE Février 2006 I. L'INTRODUCTION Chaque cellule d'un organisme supérieur provient de la multiplication d'une cellule préexistante (cellule
Utilisation des substrats énergétiques
 Utilisation des substrats énergétiques Collège des Enseignants de Nutrition Date de création du document 2010-2011 Table des matières I Les organes et les substrats... 3 I.1 Les substrats énergétiques...
Utilisation des substrats énergétiques Collège des Enseignants de Nutrition Date de création du document 2010-2011 Table des matières I Les organes et les substrats... 3 I.1 Les substrats énergétiques...
Transport des gaz dans le sang
 UE3-2 - Physiologie Physiologie Respiratoire Chapitre 9 : Transport des gaz dans le sang Docteur Sandrine LAUNOIS-ROLLINAT Année universitaire 2011/2012 Université Joseph Fourier de Grenoble - Tous droits
UE3-2 - Physiologie Physiologie Respiratoire Chapitre 9 : Transport des gaz dans le sang Docteur Sandrine LAUNOIS-ROLLINAT Année universitaire 2011/2012 Université Joseph Fourier de Grenoble - Tous droits
Partie Observer : Ondes et matière CHAP 04-ACT/DOC Analyse spectrale : Spectroscopies IR et RMN
 Partie Observer : Ondes et matière CHAP 04-ACT/DOC Analyse spectrale : Spectroscopies IR et RMN Objectifs : Exploiter un spectre infrarouge pour déterminer des groupes caractéristiques Relier un spectre
Partie Observer : Ondes et matière CHAP 04-ACT/DOC Analyse spectrale : Spectroscopies IR et RMN Objectifs : Exploiter un spectre infrarouge pour déterminer des groupes caractéristiques Relier un spectre
4 : MÉTHODES D ANALYSE UTILISÉES EN ÉCOLOGIE MICROBIENNE
 4 : MÉTHODES D ANALYSE UTILISÉES EN ÉCOLOGIE MICROBIENNE L écologie microbienne (ou étude des micro-organismes de l environnement) étudie : les relations entre les différentes populations de micro-organismes
4 : MÉTHODES D ANALYSE UTILISÉES EN ÉCOLOGIE MICROBIENNE L écologie microbienne (ou étude des micro-organismes de l environnement) étudie : les relations entre les différentes populations de micro-organismes
Effets électroniques-acidité/basicité
 Université du Maine Faculté des Sciences Retour Révisions de cours Effets électroniquesacidité/basicité Il est très important dans un cours de himie organique de connaitre ces notions qui vont intervenir
Université du Maine Faculté des Sciences Retour Révisions de cours Effets électroniquesacidité/basicité Il est très important dans un cours de himie organique de connaitre ces notions qui vont intervenir
Interactions des rayonnements avec la matière
 UE3-1 : Biophysique Chapitre 2 : Interactions des rayonnements avec la matière Professeur Jean-Philippe VUILLEZ Année universitaire 2011/2012 Université Joseph Fourier de Grenoble - Tous droits réservés.
UE3-1 : Biophysique Chapitre 2 : Interactions des rayonnements avec la matière Professeur Jean-Philippe VUILLEZ Année universitaire 2011/2012 Université Joseph Fourier de Grenoble - Tous droits réservés.
CHAPITRE 2 : Structure électronique des molécules
 CHAPITRE 2 : Structure électronique des molécules I. La liaison covalente 1) Formation d une liaison covalente Les molécules sont des assemblages d atomes liés par des liaisons chimiques résultant d interactions
CHAPITRE 2 : Structure électronique des molécules I. La liaison covalente 1) Formation d une liaison covalente Les molécules sont des assemblages d atomes liés par des liaisons chimiques résultant d interactions
EXERCICES : MECANISMES DE L IMMUNITE : pages 406 407 408 409 410
 EXERCICES : MECANISMES DE L IMMUNITE : pages 406 407 408 409 410 EXERCICE 1 PAGE 406 : EXPERIENCES A INTERPRETER Question : rôles respectifs du thymus et de la moelle osseuse dans la production des lymphocytes.
EXERCICES : MECANISMES DE L IMMUNITE : pages 406 407 408 409 410 EXERCICE 1 PAGE 406 : EXPERIENCES A INTERPRETER Question : rôles respectifs du thymus et de la moelle osseuse dans la production des lymphocytes.
1.2 Coordinence. Notion de liaison de coordinence : Cas de NH 3. et NH 4+ , 3 liaisons covalentes + 1 liaison de coordinence.
 Règle de l octet : tendance qu on les atomes à s entourer de 8 électrons dans l édifice moléculaire. Ce n est pas une règle générale. Composés respectant la règle de l octet Composés ne respectant pas
Règle de l octet : tendance qu on les atomes à s entourer de 8 électrons dans l édifice moléculaire. Ce n est pas une règle générale. Composés respectant la règle de l octet Composés ne respectant pas
Respiration Mitochondriale
 Université Pierre et Marie Curie Respiration Mitochondriale Objectifs au cours de Révisions Biochimie PCEM2 Révisions Biochimie Métabolique 2004-2005 Pr. A. Raisonnier ([email protected]) Mise à
Université Pierre et Marie Curie Respiration Mitochondriale Objectifs au cours de Révisions Biochimie PCEM2 Révisions Biochimie Métabolique 2004-2005 Pr. A. Raisonnier ([email protected]) Mise à
CATALOGUE DES PRESTATIONS DE LA
 1/23 La plate-forme Biopuces et Séquençage de Strasbourg est équipée des technologies Affymetrix et Agilent pour l étude du transcriptome et du génome sur puces à ADN. SOMMAIRE ANALYSE TRANSCRIPTIONNELLE...
1/23 La plate-forme Biopuces et Séquençage de Strasbourg est équipée des technologies Affymetrix et Agilent pour l étude du transcriptome et du génome sur puces à ADN. SOMMAIRE ANALYSE TRANSCRIPTIONNELLE...
Les outils de génétique moléculaire Les techniques liées aux acides nucléiques
 Les outils de génétique moléculaire Les techniques liées aux acides nucléiques Sommaire Preparation des acides nucléiques Extraction / purification Les enzymes agissant sur les acides nucléiques Les enzymes
Les outils de génétique moléculaire Les techniques liées aux acides nucléiques Sommaire Preparation des acides nucléiques Extraction / purification Les enzymes agissant sur les acides nucléiques Les enzymes
Compétence 3-1 S EXPRIMER A L ECRIT Fiche professeur
 Compétence 3-1 S EXPRIMER A L ECRIT Fiche professeur Nature de l activité : Réaliser 3 types de productions écrites (réécriture de notes, production d une synthèse de documents, production d une argumentation)
Compétence 3-1 S EXPRIMER A L ECRIT Fiche professeur Nature de l activité : Réaliser 3 types de productions écrites (réécriture de notes, production d une synthèse de documents, production d une argumentation)
chez les eucaryotes, source de diversification et de modulation de l'expression des gènes
 L'initiation de la traduction chez les eucaryotes, source de diversification et de modulation de l'expression des gènes Olivier Jean-Jean, Michel Cassan, Jean-Pierre Rousset Société Française de Génétique
L'initiation de la traduction chez les eucaryotes, source de diversification et de modulation de l'expression des gènes Olivier Jean-Jean, Michel Cassan, Jean-Pierre Rousset Société Française de Génétique
Transport des gaz dans le sang
 UE3-2 - Physiologie Physiologie Respiratoire Chapitre 9 : Transport des gaz dans le sang Docteur Sandrine LAUNOIS-ROLLINAT Année universitaire 2010/2011 Université Joseph Fourier de Grenoble - Tous droits
UE3-2 - Physiologie Physiologie Respiratoire Chapitre 9 : Transport des gaz dans le sang Docteur Sandrine LAUNOIS-ROLLINAT Année universitaire 2010/2011 Université Joseph Fourier de Grenoble - Tous droits
Le rôle de l endocytose dans les processus pathologiques
 UE7 Cours n 9 C. LAMAZE 24.11.11 Elise GODEAU (partie1) Guillaume MERGENTHALER (partie2) Le rôle de l endocytose dans les processus pathologiques SOMMAIRE : I. L endocytose à récepteurs : la voie des clathrines
UE7 Cours n 9 C. LAMAZE 24.11.11 Elise GODEAU (partie1) Guillaume MERGENTHALER (partie2) Le rôle de l endocytose dans les processus pathologiques SOMMAIRE : I. L endocytose à récepteurs : la voie des clathrines
Génétique et génomique Pierre Martin
 Génétique et génomique Pierre Martin Principe de la sélections Repérage des animaux intéressants X Accouplements Programmés Sélection des meilleurs mâles pour la diffusion Index diffusés Indexation simultanée
Génétique et génomique Pierre Martin Principe de la sélections Repérage des animaux intéressants X Accouplements Programmés Sélection des meilleurs mâles pour la diffusion Index diffusés Indexation simultanée
ANTIVIRAUX ET INTERFERONS LES ANTIVIRAUX
 ANTIVIRAUX ET INTERFERONS LES ANTIVIRAUX La thérapeutique antivirale progresse avec lenteur car elle s'attaque à des micro-organismes ne se multipliant qu'à l'intérieur des cellules vivantes dont ils détournent
ANTIVIRAUX ET INTERFERONS LES ANTIVIRAUX La thérapeutique antivirale progresse avec lenteur car elle s'attaque à des micro-organismes ne se multipliant qu'à l'intérieur des cellules vivantes dont ils détournent
MAB Solut. vos projets. MABLife Génopole Campus 1 5 rue Henri Desbruères 91030 Evry Cedex. www.mabsolut.com. intervient à chaque étape de
 Mabsolut-DEF-HI:Mise en page 1 17/11/11 17:45 Page1 le département prestataire de services de MABLife de la conception à la validation MAB Solut intervient à chaque étape de vos projets Création d anticorps
Mabsolut-DEF-HI:Mise en page 1 17/11/11 17:45 Page1 le département prestataire de services de MABLife de la conception à la validation MAB Solut intervient à chaque étape de vos projets Création d anticorps
COUSIN Fabien KERGOURLAY Gilles. 19 octobre 2007. de l hôte par les. Master 2 MFA Responsable : UE Incidence des paramètres environnementaux
 COUSIN Fabien KERGOURLAY Gilles 19 octobre 2007 Inhibition des défenses de l hôte par les bactéries pathogènes Master 2 MFA Responsable : UE Incidence des paramètres environnementaux Gwennola ERMEL I Détection
COUSIN Fabien KERGOURLAY Gilles 19 octobre 2007 Inhibition des défenses de l hôte par les bactéries pathogènes Master 2 MFA Responsable : UE Incidence des paramètres environnementaux Gwennola ERMEL I Détection
Relation entre deux variables : estimation de la corrélation linéaire
 CHAPITRE 3 Relation entre deux variables : estimation de la corrélation linéaire Parmi les analyses statistiques descriptives, l une d entre elles est particulièrement utilisée pour mettre en évidence
CHAPITRE 3 Relation entre deux variables : estimation de la corrélation linéaire Parmi les analyses statistiques descriptives, l une d entre elles est particulièrement utilisée pour mettre en évidence
Séquence 1. Reproduction conforme de la cellule et réplication de l ADN Variabilité génétique et mutation de l ADN
 Séquence 1 Reproduction conforme de la cellule et réplication de l ADN Variabilité génétique et mutation de l ADN Sommaire 1. Reproduction conforme de la cellule et réplication de l ADN 2. Variabilité
Séquence 1 Reproduction conforme de la cellule et réplication de l ADN Variabilité génétique et mutation de l ADN Sommaire 1. Reproduction conforme de la cellule et réplication de l ADN 2. Variabilité
Compléments - Chapitre 5 Spectroscopie
 ompléments - hapitre 5 Spectroscopie Spectroscopie par résonance magnétique nucléaire (RMN 13 ) Tandis que la spectroscopie RMN 1 H fournit des données sur la disposition des atomes d'hydrogène dans une
ompléments - hapitre 5 Spectroscopie Spectroscopie par résonance magnétique nucléaire (RMN 13 ) Tandis que la spectroscopie RMN 1 H fournit des données sur la disposition des atomes d'hydrogène dans une
Détection des duplications en tandem au niveau nucléique à l'aide de la théorie des flots
 Université Toulouse 3 Paul Sabatier(UT3 Paul Sabatier) Informatique Spécialité Bioinformatique Eric AUDEMARD lundi 28 novembre 2011 Détection des duplications en tandem au niveau nucléique à l'aide de
Université Toulouse 3 Paul Sabatier(UT3 Paul Sabatier) Informatique Spécialité Bioinformatique Eric AUDEMARD lundi 28 novembre 2011 Détection des duplications en tandem au niveau nucléique à l'aide de
Chapitre 6. Réactions nucléaires. 6.1 Généralités. 6.1.1 Définitions. 6.1.2 Lois de conservation
 Chapitre 6 Réactions nucléaires 6.1 Généralités 6.1.1 Définitions Un atome est constitué d électrons et d un noyau, lui-même constitué de nucléons (protons et neutrons). Le nombre de masse, noté, est le
Chapitre 6 Réactions nucléaires 6.1 Généralités 6.1.1 Définitions Un atome est constitué d électrons et d un noyau, lui-même constitué de nucléons (protons et neutrons). Le nombre de masse, noté, est le
Partie 1. Addition nucléophile suivie d élimination (A N + E) 1.1. Réactivité électrophile des acides carboxyliques et groupes dérivés
 1.1. Réactivité électrophile des acides carboxyliques et groupes dérivés") Molécules et matériaux organiques Partie 1. Addition nucléophile suivie d élimination (A N + E) 1.1. Réactivité électrophile des acides carboxyliques et groupes dérivés bjectifs du chapitre Notions à connaître
Molécules et matériaux organiques Partie 1. Addition nucléophile suivie d élimination (A N + E) 1.1. Réactivité électrophile des acides carboxyliques et groupes dérivés bjectifs du chapitre Notions à connaître
Résonance Magnétique Nucléaire : RMN
 21 Résonance Magnétique Nucléaire : RMN Salle de TP de Génie Analytique Ce document résume les principaux aspects de la RMN nécessaires à la réalisation des TP de Génie Analytique de 2ème année d IUT de
21 Résonance Magnétique Nucléaire : RMN Salle de TP de Génie Analytique Ce document résume les principaux aspects de la RMN nécessaires à la réalisation des TP de Génie Analytique de 2ème année d IUT de
Comprendre l Univers grâce aux messages de la lumière
 Seconde / P4 Comprendre l Univers grâce aux messages de la lumière 1/ EXPLORATION DE L UNIVERS Dans notre environnement quotidien, les dimensions, les distances sont à l échelle humaine : quelques mètres,
Seconde / P4 Comprendre l Univers grâce aux messages de la lumière 1/ EXPLORATION DE L UNIVERS Dans notre environnement quotidien, les dimensions, les distances sont à l échelle humaine : quelques mètres,
Les composites thermoplastiques
 Les composites thermoplastiques Définition Par définition, un thermoplastique (anglais :thermoplast) est un matériau à base de polymère (composé de macromolécules) qui peut être mis en forme, à l état
Les composites thermoplastiques Définition Par définition, un thermoplastique (anglais :thermoplast) est un matériau à base de polymère (composé de macromolécules) qui peut être mis en forme, à l état
Critères pour les méthodes de quantification des résidus potentiellement allergéniques de protéines de collage dans le vin (OIV-Oeno 427-2010)
") Méthode OIV- -MA-AS315-23 Type de méthode : critères Critères pour les méthodes de quantification des résidus potentiellement allergéniques de protéines de collage (OIV-Oeno 427-2010) 1 Définitions des
Méthode OIV- -MA-AS315-23 Type de méthode : critères Critères pour les méthodes de quantification des résidus potentiellement allergéniques de protéines de collage (OIV-Oeno 427-2010) 1 Définitions des
IV- Comment fonctionne un ordinateur?
 1 IV- Comment fonctionne un ordinateur? L ordinateur est une alliance du hardware (le matériel) et du software (les logiciels). Jusqu à présent, nous avons surtout vu l aspect «matériel», avec les interactions
1 IV- Comment fonctionne un ordinateur? L ordinateur est une alliance du hardware (le matériel) et du software (les logiciels). Jusqu à présent, nous avons surtout vu l aspect «matériel», avec les interactions
Hémochromatose génétique non liée à HFE-1 : quand et comment la rechercher? Cécilia Landman 11 décembre 2010
 Hémochromatose génétique non liée à HFE-1 : quand et comment la rechercher? Cécilia Landman 11 décembre 2010 Métabolisme du fer : hepcidine Fer absorbé par les entérocytes des villosités duodénales : transporteur
Hémochromatose génétique non liée à HFE-1 : quand et comment la rechercher? Cécilia Landman 11 décembre 2010 Métabolisme du fer : hepcidine Fer absorbé par les entérocytes des villosités duodénales : transporteur
La reconnaissance moléculaire: la base du design rationnel Modélisation moléculaire: Introduction Hiver 2006
 La reconnaissance moléculaire: la base du design rationnel En 1890 Emil Fisher a proposé le modèle "serrure et clé" pour expliquer la façon de fonctionner des systèmes biologiques. Un substrat rentre et
La reconnaissance moléculaire: la base du design rationnel En 1890 Emil Fisher a proposé le modèle "serrure et clé" pour expliquer la façon de fonctionner des systèmes biologiques. Un substrat rentre et
BREVET DE TECHNICIEN SUPÉRIEUR QUALITÉ DANS LES INDUSTRIES ALIMENTAIRES ET LES BIO-INDUSTRIES
 ~--------------~~-----~- ----~-- Session 2009 BREVET DE TECNICIEN SUPÉRIEUR QUALITÉ DANS LES INDUSTRIES ALIMENTAIRES ET LES BIO-INDUSTRIES U22 - SCIENCES PYSIQUES Durée: 2 heures Coefficient : 3 Les calculatrices
~--------------~~-----~- ----~-- Session 2009 BREVET DE TECNICIEN SUPÉRIEUR QUALITÉ DANS LES INDUSTRIES ALIMENTAIRES ET LES BIO-INDUSTRIES U22 - SCIENCES PYSIQUES Durée: 2 heures Coefficient : 3 Les calculatrices
Tableau récapitulatif : composition nutritionnelle de la spiruline
 Tableau récapitulatif : composition nutritionnelle de la spiruline (Valeur énergétique : 38 kcal/10 g) Composition nutritionnelle Composition pour 10 g Rôle Protéines (végétales) 55 à 70 % Construction
Tableau récapitulatif : composition nutritionnelle de la spiruline (Valeur énergétique : 38 kcal/10 g) Composition nutritionnelle Composition pour 10 g Rôle Protéines (végétales) 55 à 70 % Construction
Molécules et Liaison chimique
 Molécules et liaison chimique Molécules et Liaison chimique La liaison dans La liaison dans Le point de vue classique: l approche l de deux atomes d hydrogd hydrogènes R -0,9-1 0 0,5 1 1,5,5 3 3,5 4 R
Molécules et liaison chimique Molécules et Liaison chimique La liaison dans La liaison dans Le point de vue classique: l approche l de deux atomes d hydrogd hydrogènes R -0,9-1 0 0,5 1 1,5,5 3 3,5 4 R
leurs produits respectifs, et production d'anticorps spécinques. Mémoire présenté Département de biochimie Université Laval
 Philippe Garneau Étude de la régulation de l'expression des gènes cyse et cyss de Batcillus subtitis : Surproduction et puriffcation de leurs produits respectifs, et production d'anticorps spécinques.
Philippe Garneau Étude de la régulation de l'expression des gènes cyse et cyss de Batcillus subtitis : Surproduction et puriffcation de leurs produits respectifs, et production d'anticorps spécinques.
Vous avez plusieurs prises. ou + B. Vous n avez pas de prise à 6 pôles. 1 Préparez le matériel nécessaire.
 Vous avez plusieurs prises B. Vous n avez pas de prise à 6 pôles Une habitation peut posséder plusieurs prises. La ligne Belgacom n entre toutefois chez vous que par une seule prise. Nous l appellerons
Vous avez plusieurs prises B. Vous n avez pas de prise à 6 pôles Une habitation peut posséder plusieurs prises. La ligne Belgacom n entre toutefois chez vous que par une seule prise. Nous l appellerons
Information pour le patient
 Information pour le patient Plus de qualité de vie avec des implants dentaires SWISS PRECISION AND INNOVATION. 1 Table des matières Quelle est l importance des dents pour notre qualité de vie? 4 Qu est-ce
Information pour le patient Plus de qualité de vie avec des implants dentaires SWISS PRECISION AND INNOVATION. 1 Table des matières Quelle est l importance des dents pour notre qualité de vie? 4 Qu est-ce
3 Charges électriques
 3 Charges électriques 3.1 Electrisation par frottement Expérience : Frottons un bâton d ébonite avec un morceau de peau de chat. Approchonsle de petits bouts de papier. On observe que les bouts de papier
3 Charges électriques 3.1 Electrisation par frottement Expérience : Frottons un bâton d ébonite avec un morceau de peau de chat. Approchonsle de petits bouts de papier. On observe que les bouts de papier
Génomique Comparative et intégrative
 Génomique Comparative et intégrative Introduction : Le big data : on peut traiter des données massives à présent, l'objectif à présent est d'éviter les transferts de données trop longs. On a tout à portée
Génomique Comparative et intégrative Introduction : Le big data : on peut traiter des données massives à présent, l'objectif à présent est d'éviter les transferts de données trop longs. On a tout à portée
5.5.5 Exemple d un essai immunologique
 5.5.5 Exemple d un essai immunologique Test de grossesse Test en forme de bâtonnet destiné à mettre en évidence l'hormone spécifique de la grossesse, la gonadotrophine chorionique humaine (hcg), une glycoprotéine.
5.5.5 Exemple d un essai immunologique Test de grossesse Test en forme de bâtonnet destiné à mettre en évidence l'hormone spécifique de la grossesse, la gonadotrophine chorionique humaine (hcg), une glycoprotéine.
Premiers Pas avec OneNote 2013
 Premiers Pas avec OneNote 2 Présentation de OneNote 3 Ouverture du logiciel OneNote 4 Sous Windows 8 4 Sous Windows 7 4 Création de l espace de travail OneNote 5 Introduction 5 Présentation des différentes
Premiers Pas avec OneNote 2 Présentation de OneNote 3 Ouverture du logiciel OneNote 4 Sous Windows 8 4 Sous Windows 7 4 Création de l espace de travail OneNote 5 Introduction 5 Présentation des différentes
Cours d électricité. Circuits électriques en courant constant. Mathieu Bardoux. 1 re année
 Cours d électricité Circuits électriques en courant constant Mathieu Bardoux [email protected] IUT Saint-Omer / Dunkerque Département Génie Thermique et Énergie 1 re année Objectifs du chapitre
Cours d électricité Circuits électriques en courant constant Mathieu Bardoux [email protected] IUT Saint-Omer / Dunkerque Département Génie Thermique et Énergie 1 re année Objectifs du chapitre
KIT SOLAIRE EVOLUTIF DE BASE
 PARALLELE ENERGIE KIT SOLAIRE EVOLUTIF DE BASE Ce kit solaire est un système solaire facile à utiliser. Il s agit d un «générateur solaire» qui convertit la lumière du soleil en électricité, et qui peut
PARALLELE ENERGIE KIT SOLAIRE EVOLUTIF DE BASE Ce kit solaire est un système solaire facile à utiliser. Il s agit d un «générateur solaire» qui convertit la lumière du soleil en électricité, et qui peut
- pellicule de fruits qui a un rôle de prévention contre l'évaporation, le développement de moisissures et l'infection par des parasites
 LES LIPIDES Quelles Sont les Idées Clés? Les lipides sont les huiles et les graisses de la vie courante. Ils sont insolubles dans l eau. Pour les synthétiser, une réaction : l Estérification. Pour les
LES LIPIDES Quelles Sont les Idées Clés? Les lipides sont les huiles et les graisses de la vie courante. Ils sont insolubles dans l eau. Pour les synthétiser, une réaction : l Estérification. Pour les
MABioVis. Bio-informatique et la
 MABioVis Modèles et Algorithmes pour la Bio-informatique et la Visualisation Visite ENS Cachan 5 janvier 2011 MABioVis G GUY MELANÇON (PR UFR Maths Info / EPI GRAVITE) (là, maintenant) - MABioVis DAVID
MABioVis Modèles et Algorithmes pour la Bio-informatique et la Visualisation Visite ENS Cachan 5 janvier 2011 MABioVis G GUY MELANÇON (PR UFR Maths Info / EPI GRAVITE) (là, maintenant) - MABioVis DAVID
CHAPITRE VI UN ESSAI DE SYNTHESE
 CHAPITRE VI UN ESSAI DE SYNTHESE CHAPITRE VI : UN ESSAI DE SYNTHESE Introduction 1) La relation dialectique entre dynamiques démographiques et développement 2) Les dynamiques démographiques et les théories
CHAPITRE VI UN ESSAI DE SYNTHESE CHAPITRE VI : UN ESSAI DE SYNTHESE Introduction 1) La relation dialectique entre dynamiques démographiques et développement 2) Les dynamiques démographiques et les théories
Production d une protéine recombinante
 99 Production d une protéine recombinante Lic. B. PIRSON Lic. J-M. SERONT ISICHt - Mons Production de la protéine recombinante GFP (Green Fluorescent Protein d Aequoria victoria) par une bactérie ( E.
99 Production d une protéine recombinante Lic. B. PIRSON Lic. J-M. SERONT ISICHt - Mons Production de la protéine recombinante GFP (Green Fluorescent Protein d Aequoria victoria) par une bactérie ( E.
I. La levure Saccharomyces cerevisiae: mode de vie
 LES LEVURES UE «levures» -5 avril: généralités (MN Simon) -6 avril: analyse génétique (MN Simon) -6 avril: Cycle cellulaire I: la réplication (E. bailly) -7 avril: Cycle cellulaire II: la mitose (E. Bailly)
LES LEVURES UE «levures» -5 avril: généralités (MN Simon) -6 avril: analyse génétique (MN Simon) -6 avril: Cycle cellulaire I: la réplication (E. bailly) -7 avril: Cycle cellulaire II: la mitose (E. Bailly)
Chapitre 5 : Noyaux, masse et énergie
 Chapitre 5 : Noyaux, masse et énergie Connaissances et savoir-faire exigibles : () () (3) () (5) (6) (7) (8) Définir et calculer un défaut de masse et une énergie de liaison. Définir et calculer l énergie
Chapitre 5 : Noyaux, masse et énergie Connaissances et savoir-faire exigibles : () () (3) () (5) (6) (7) (8) Définir et calculer un défaut de masse et une énergie de liaison. Définir et calculer l énergie
Chapitre 11: Réactions nucléaires, radioactivité et fission
 1re B et C 11 Réactions nucléaires, radioactivité et fission 129 Chapitre 11: Réactions nucléaires, radioactivité et fission 1. Définitions a) Nucléides (= noyaux atomiques) Les nucléides renferment les
1re B et C 11 Réactions nucléaires, radioactivité et fission 129 Chapitre 11: Réactions nucléaires, radioactivité et fission 1. Définitions a) Nucléides (= noyaux atomiques) Les nucléides renferment les
1. Principes de biochimie générale. A. Bioénergétique et dynamique. a) Intro: Les mitochondries passent leur temps à fabriquer de l énergie.
 Intro: Les mitochondries passent leur temps à fabriquer de l énergie.") RESPIRATION MITOCHONDRIALE 1. Principes de biochimie générale. A. Bioénergétique et dynamique. a) Intro: Les mitochondries passent leur temps à fabriquer de l énergie. b) Rappels: L énergie chimique sert
RESPIRATION MITOCHONDRIALE 1. Principes de biochimie générale. A. Bioénergétique et dynamique. a) Intro: Les mitochondries passent leur temps à fabriquer de l énergie. b) Rappels: L énergie chimique sert
TP3 Test immunologique et spécificité anticorps - déterminant antigénique
 TP3 Test immunologique et spécificité anticorps - déterminant antigénique Partie 1 : Spécificité d'un anticorps pour un déterminant antigénique du VIH La séropositivité pour le VIH correspond à la présence
TP3 Test immunologique et spécificité anticorps - déterminant antigénique Partie 1 : Spécificité d'un anticorps pour un déterminant antigénique du VIH La séropositivité pour le VIH correspond à la présence
Eléments constitutifs et synthèse des convertisseurs statiques. Convertisseur statique CVS. K à séquences convenables. Source d'entrée S1
 1 Introduction Un convertisseur statique est un montage utilisant des interrupteurs à semiconducteurs permettant par une commande convenable de ces derniers de régler un transfert d énergie entre une source
1 Introduction Un convertisseur statique est un montage utilisant des interrupteurs à semiconducteurs permettant par une commande convenable de ces derniers de régler un transfert d énergie entre une source
Windows Internet Name Service (WINS)
") Windows Internet Name Service (WINS) WINDOWS INTERNET NAME SERVICE (WINS)...2 1.) Introduction au Service de nom Internet Windows (WINS)...2 1.1) Les Noms NetBIOS...2 1.2) Le processus de résolution WINS...2
Windows Internet Name Service (WINS) WINDOWS INTERNET NAME SERVICE (WINS)...2 1.) Introduction au Service de nom Internet Windows (WINS)...2 1.1) Les Noms NetBIOS...2 1.2) Le processus de résolution WINS...2
CHAPITRE 3 LES TASQ COMME PLATEFORME POUR LA CATALYSE PSEUDO-ENZYMATIQUE
 L. Stefan Chapitre 3 CHAPITRE 3 LES TASQ COMME PLATEFORME POUR LA CATALYSE PSEUDO-ENZYMATIQUE Nous avons démontré dans le chapitre précédent que les TASQ hydrosolubles développés au cours de cette thèse
L. Stefan Chapitre 3 CHAPITRE 3 LES TASQ COMME PLATEFORME POUR LA CATALYSE PSEUDO-ENZYMATIQUE Nous avons démontré dans le chapitre précédent que les TASQ hydrosolubles développés au cours de cette thèse
THEME 2. LE SPORT CHAP 1. MESURER LA MATIERE: LA MOLE
 THEME 2. LE SPORT CHAP 1. MESURER LA MATIERE: LA MOLE 1. RAPPEL: L ATOME CONSTITUANT DE LA MATIERE Toute la matière de l univers, toute substance, vivante ou inerte, est constituée à partir de particules
THEME 2. LE SPORT CHAP 1. MESURER LA MATIERE: LA MOLE 1. RAPPEL: L ATOME CONSTITUANT DE LA MATIERE Toute la matière de l univers, toute substance, vivante ou inerte, est constituée à partir de particules
Luca : à la recherche du plus proche ancêtre commun universel Patrick Forterre, Simonetta Gribaldo, Céline Brochier
 MEDECINE/SCIENCES 2005 ; 21 :???-??? > L application des méthodes de la biologie moléculaire à l étude des premières étapes de l évolution a montré que le monde vivant pouvait être divisé en trois domaines
MEDECINE/SCIENCES 2005 ; 21 :???-??? > L application des méthodes de la biologie moléculaire à l étude des premières étapes de l évolution a montré que le monde vivant pouvait être divisé en trois domaines
Les bases de données transcriptionnelles en ligne
 Les bases de données transcriptionnelles en ligne Différents concepts en régulation transcriptionnelle sites de fixation - in vitro/vivo? - quelle technique? - degré de confiance? facteur de transcription
Les bases de données transcriptionnelles en ligne Différents concepts en régulation transcriptionnelle sites de fixation - in vitro/vivo? - quelle technique? - degré de confiance? facteur de transcription
L obligation de négocier sur la pénibilité dans les entreprises. Premiers éléments de bilan. Direction générale du travail
 CONSEIL D ORIENTATION DES RETRAITES Séance plénière du 21 novembre 2012 à 14 h 30 «Pénibilité. Transition emploi-retraite. Elaboration de cas-types pour les projections.» Document N 6 Document de travail,
CONSEIL D ORIENTATION DES RETRAITES Séance plénière du 21 novembre 2012 à 14 h 30 «Pénibilité. Transition emploi-retraite. Elaboration de cas-types pour les projections.» Document N 6 Document de travail,
UE : GENE 2208. Responsable : Enseignant : ECUE 1. Enseignant : ECUE 2. Dr COULIBALY Foungotin Hamidou
 UE : GENE 2208 Responsable : Enseignant : ECUE 1 Enseignant : ECUE 2 Dr COULIBALY Foungotin Hamidou Spécialités : Génétique Humaine Biologie de la Procréation Cytogénétique Spermiologie Essais Cliniques
UE : GENE 2208 Responsable : Enseignant : ECUE 1 Enseignant : ECUE 2 Dr COULIBALY Foungotin Hamidou Spécialités : Génétique Humaine Biologie de la Procréation Cytogénétique Spermiologie Essais Cliniques
INSTRUCTIONS D INSTALLATION ET D ENLÈVEMENT : LÈVE-VITRE
 ENLÈVEMENT/INSTALLATION DU LÈVE-VITRE (741-649) Dodge Durango 1998 2003, Dakota Quad Cab 2000 03 Conseils De Tech Général : Utiliser du ruban du peintre plutôt que conduit la bande à fenêtre sécurisée.
ENLÈVEMENT/INSTALLATION DU LÈVE-VITRE (741-649) Dodge Durango 1998 2003, Dakota Quad Cab 2000 03 Conseils De Tech Général : Utiliser du ruban du peintre plutôt que conduit la bande à fenêtre sécurisée.
Atelier B : Maintivannes
 Atelier B : Maintivannes BMA03 : Procédure de réglage de la vanne de régulation CAMFLEX BMA03 : Procédure de réglage de la vanne CAMFLEX Page 1 Buts de l exercice : - Réaliser la manutention d un élément
Atelier B : Maintivannes BMA03 : Procédure de réglage de la vanne de régulation CAMFLEX BMA03 : Procédure de réglage de la vanne CAMFLEX Page 1 Buts de l exercice : - Réaliser la manutention d un élément
Semestre 2 Spécialité «Analyse in silico des complexes macromolécules biologiques-médicaments»
 Master In silico Drug Design Semestre 2 Spécialité «Analyse in silico des complexes macromolécules biologiques-médicaments» 30NU01IS INITIATION A LA PROGRAMMATION (6 ECTS) Responsables : D. MESTIVIER,
Master In silico Drug Design Semestre 2 Spécialité «Analyse in silico des complexes macromolécules biologiques-médicaments» 30NU01IS INITIATION A LA PROGRAMMATION (6 ECTS) Responsables : D. MESTIVIER,
1 ère partie : tous CAP sauf hôtellerie et alimentation CHIMIE ETRE CAPABLE DE. PROGRAMME - Atomes : structure, étude de quelques exemples.
 Référentiel CAP Sciences Physiques Page 1/9 SCIENCES PHYSIQUES CERTIFICATS D APTITUDES PROFESSIONNELLES Le référentiel de sciences donne pour les différentes parties du programme de formation la liste
Référentiel CAP Sciences Physiques Page 1/9 SCIENCES PHYSIQUES CERTIFICATS D APTITUDES PROFESSIONNELLES Le référentiel de sciences donne pour les différentes parties du programme de formation la liste