I / L organisation du génome nucléaire
|
|
|
- Joëlle Marier
- il y a 8 ans
- Total affichages :
Transcription
1 Définition du sujet et limites : Le génome contient l information biologique nécessaire au développement et à la réalisation de toutes les fonctions des organismes. Il correspond à l ensemble du contenu d ADN (acide désoxyribonucléique). L ADN étant le support universel de l information génétique, et donc pas des seuls Eucaryotes, sa structure sera présentée rapidement sans développer les expériences historiques qui ont permis de l élucider et de montrer son rôle. En revanche, pourront être évoquées les expériences qui ont permis le décryptage du génome. Dans les cellules eucaryotes, celui-ci est pour l essentiel situé dans le noyau et définit le génome nucléaire, porté par les chromosomes. De l ADN s observe également dans les organites semi-autonomes, mitochondries et chloroplastes, constituant respectivement les génomes mitochondriaux et chloroplastiques. L accomplissement des fonctions biologiques repose sur l intervention de nombreuses protéines, dont la séquence est codée par l information génétique de l ADN. Ces protéines représentent ainsi l expression du génome, du moins des parties codantes de celui-ci, transcrites en ARNm. Une large part du génome ne conduit cependant pas à l expression de protéines. L ADN code par exemple un certain nombre d ARN qui ont par eux-mêmes une fonction et qui ne sont pas traduits en protéines : c est le cas des ARN ribosomiques, des ARN de transfert et de nombreux petits ARN catalytiques qui peuvent par exemple intervenir dans la maturation des ARN messagers. L ensemble de toutes ces parties transcrites constitue les gènes. D autres parties de l ADN, pourtant développées et parfois présentes en séquences répétitives, ne sont le siège d aucune activité de transcription. Problématique : L étude de l organisation du génome suppose que soient envisagées la part relative des différents types de séquences, leur distribution dans l ADN et dans les structures chromosomiques et leur signification informative. Elle peut être précédée d un bref rappel de la structure de la molécule en dégageant ce qui lui confère cette extraordinaire capacité d encodage de l information génétique et sa reproductibilité mais l organisation de l ADN en soi est hors sujet. La dimension du génome eucaryote découpé en chromosomes linéaires impose une compaction nécessaire à son stockage autant qu à son activité, il s agira d en préciser les modalités. L étude de l expression est d abord celle de la transcription, puis celle de la maturation des ARN, qui représentent deux étapes clés dans la production et la régulation des flux d informations au sein d une cellule. Chez les eucaryotes, la localisation nucléaire du génome introduit une étape supplémentaire dans la régulation de son expression, régulation qui sera présentée à chaque étape de l expression du génome. I / L organisation du génome nucléaire A / Organisation chromosomique et taille du génome 1 / Organisation chromosomique. Chaque chromosome d Eucaryote ne contient qu une seule molécule d ADN linéaire. Le nombre et la structure des chromosomes sont caractéristiques de chaque espèce et définissent le caryotype de celle-ci. Chaque chromosome peut présenter quatre types d espèces moléculaires : l ADN double brin, des protéines d empaquetage : les histones, différents types d ARN messagers, de transfert et ribosomiques résultant de la transcription, les protéines intervenant dans la transcription, la réplication et la réparation de l ADN. On ne traitera pas précisément des mécanismes d empaquetage. Il importe de rappeler que l ADN est structuré en chromatine par des protéines. On mentionnera la différence existant entre euchromatine (zones faiblement colorées) et hétérochromatine (zones fortement colorées).
2 Des expériences de cartographie des chromosomes montrent que la plupart des gènes actifs qui sont identifiables grâce à des mutations sont situés dans l euchromatine. Celle-ci se colore moins intensément parce qu elle est empaquetée de façon plus lâche. L idée généralement acceptée est qu il s agit de l état le plus compatible avec la transcription et l activité des gènes. Chez la plupart des organismes, l hétérochromatine se trouve surtout de part et d autre des centromères. 2 / Taille du génome et nombre de gènes Mesure par densitométrie après coloration Feulgen et maintenant par fluorométrie (ou cytométrie à flux) où l ADN est marqué par du DAPI, une molécule fluorescente qui se lie fixement à l ADN et émet une fluorescence bleue brillante lorsqu elle est éclairée par des UV. Discussion des relations entre nombre de gènes, taille du génome et complexité des organismes. Portion de l ADN finalement codant. - Estimation du nombre de gènes : Précédemment, pour estimer le nombre de gènes d une espèce, on utilisait les résultats génétiques ou on comparait la quantité d ADN présente dans cette espèce à celle d autres espèces connues. Actuellement, les estimations issues de projets génomes proviennent des programmes informatiques de recherche des gènes dans les séquences d ADN. Dans certains cas, la première méthode a pu conduire à des sous-estimations du nombre de gènes lorsque des gènes ne présentaient pas de forme mutante identifiable par un phénotype particulier. La génomique a permis d entreprendre le séquençage du matériel génétique de nombreux organismes eucaryotes : Saccharomyces cerevisiae, Caenorhabditis elegans, Drosophila melanogaster, Arabidopsis thaliana, génome humain. Actuellement le génome humain est estimé à environ gènes. Drosophile : environ à gènes. Caenorhabditis : environ gènes Saccharomyces : environ 6000 gènes Arabidopsis : environ gènes - La taille du génome peut être exprimée en nombre de bases : Homme : ( soit 3000 Mb) Drosophile : 1, Caenorhabditis : 9, Arabidopsis : 10 8 De manière plus générale, la taille du génome des Eucaryotes s étend de 10 Mb (Saccharomyces : 12 Mb) à Mb (Fritillaire). Certains végétaux ont un génome important (Triticum : Mb). - Les relations entre nombre de gènes, taille du génome et complexité des organismes sont difficiles à discuter : - La taille du génome n est pas directement en relation avec le nombre de gènes : il y a plus de gènes chez Caenorhabditis que chez la Drosophile, alors que son génome est plus petit. - Le génome de la levure est 250 fois plus petit que celui de l homme et pourtant son nombre de gènes n est pas 100 mais de près de Le génome humain est 30 fois plus petit que celui de certains végétaux et amphibiens et 200 fois plus petit que celui d une espèce d amibe. - La teneur en ADN de génomes d organismes similaires (ex : poissons osseux, amphibiens) peut varier de plusieurs centaines de fois alors que le génome contient grossièrement le même nombre de gènes. - Des espèces très apparentées et ayant des tailles de génome identique peuvent avoir un nombre très différent de chromosomes de tailles différentes : il n y a pas de relations simples entre le nombre de chromosomes, la complexité des espèces et la taille totale du génome. Ces valeurs suggèrent l existence de parties plus ou moins importantes non exprimées (génome de la levure : moins de gènes morcelés : 239 introns pour l ensemble du génome alors qu on peut observer
 On peut utiliser ici les résultats d")
3 chez des Eucaryotes plus complexes, jusqu à 100 introns dans un seul gène ; moins de séquences répétitives). - L ADN finalement codant chez l homme ne représente que 3 % du génome. B / Les différents types de séquences Résultats d expériences de réassociation de l ADN, % de réassociation en fonction de Cot (quantité totale d ADN x temps) On peut utiliser ici les résultats d expériences de réassociation de l ADN : des échantillons d ADN provenant de noyaux eucaryotes sont chauffés pour provoquer leur dénaturation, puis refroidis lentement. On suit alors la cinétique de réassociation de séquences complémentaires, initialement dissociées et qui se retrouvent grâce aux mouvements aléatoires des molécules en solution. On observe que l ADN se réassocie beaucoup plus rapidement qu il n était prévu dans l hypothèse de brins séparés de gènes uniques. Ces résultats ne s expliquent qu en postulant l existence de séquences d ADN présentes en de nombreux exemplaires dans le génome : ces expériences conduisent à mettre en évidence l ADN répétitif (avec des séquences moyennement et fortement répétées). On distinguera ainsi des séquences en un seul exemplaire, qui correspondent à des gènes fonctionnels et des séquences répétitives, qui peuvent correspondre à des séquences fonctionnelles ou à des séquences dont la fonction reste mal connue. C / Les séquences répétitives 1 / Les séquences répétitives codantes a / les familles de gènes dispersées Plusieurs types de protéines sont codées par des familles de gènes homologues, dispersés dans tout le génome. De telles familles peuvent ne comporter que peu de gènes ou au contraire beaucoup : - Actines : de 5 à 30 gènes. - Kératines : plus de 20 gènes. - Chaîne lourde de la myosine : 5 à 10 gènes. - Histones : 100 à 1000 gènes. Les séquences des gènes d une famille peuvent diverger et les gènes homologues avoir acquis des fonctions légèrement différentes. Certains gènes ont cessé d être fonctionnels et sont devenus des pseudogènes. Un pseudogène est une séquence nucléotidique de l ADN qui ressemble beaucoup à celle d un gène fonctionnel mais qui contient de nombreuses mutations (par délétion) empêchant sa propre expression. La plupart des pseudogènes proviennent de la duplication d un gène fonctionnel suivie de l accumulation de mutations qui ont endommagé la copie. Gènes le plus souvent morcelés : exons et introns b / les familles de gènes alignés en tandem
4 Les gènes des ARN structuraux (ARNTt) Les gènes des histones L organisateur nucléolaire Les cellules ont besoin de grandes quantités de produits de certains gènes et les familles de ces gènes ont évolué sous forme de séquences en tandem. Exemple de l organisateur nucléolaire : l organisateur nucléolaire des chromosomes X et Y de la drosophile contient respectivement 250 et 150 copies en tandem des gènes d ARNr. Une telle redondance est un moyen de garantir une grande quantité d ARNr par cellule. Les gènes des histones de certaines espèces dont la drosophile ou l oursin sont alignés en tandem, avec de multiples copies identiques (ce qui suppose un mécanisme assurant l invariance des copies). 2 / Les séquences répétitives en tandem et non codantes : l ADN satellite 20 % du génome humain constitué de séquences répétitives non fonctionnelles. a / Identification de l ADN satellite Ultracentrifugation dans un gradient de CsCl2, se distingue par une densité moindre (1,7) (densité de GC 40,3%) Constitué de très longs alignements (plusieurs centaines de kb) qui se retrouvent au niveau des centromères, dans les régions télomériques et dans une partie du chromosome Y des Mammifères. ADN satellite : le terme d ADN satellite provient de la manière dont celui-ci a été isolé pour la première fois. Une technique de purification de l ADN consiste à procéder à une ultracentrifugation dans une solution de chlorure de césium. Sous l effet de la centrifugation, ce sel se répartit dans un gradient de concentration croissante depuis le haut jusqu au bas du tube. Les fragments d ADN migrent dans la région du tube qui correspond à leur propre densité. Celle-ci est d autant plus forte que l ADN est riche en C-G. Il se forme une bande principale d ADN correspondant à la densité conférée par le contenu en C- G moyen caractéristique de l espèce étudiée. Des bandes de densité différente, appelées satellites parce qu elles entourent la bande majoritaire, peuvent être observées. Si la composition en C-G de ce motif dévie de la composition moyenne en C-G de l organisme considéré, il s ensuit une densité différente de ces fragments. Exemple de l ADN humain : rupture de l ADN en fragments de 50 à 100 kb. Une bande principale avec une densité moyenne de 1,701 et qui correspond à une teneur moyenne en séquences CG de 40,3 %. 3 bandes satellites de densité 1,687 ; 1,693 ; 1,697 qui traduisent des zones de séquences en CG différentes. Les régions d ADN hautement répétitives issues d une chromosome eucaryote, sont généralement identifiables par leur composition particulière en nucléotides. L ADN satellite n est pas transcrit et n a pas de fonction connue. (Rmq : Pas d ADN satellites chez les Procaryotes). b / L ADN centromérique hautement répétitif La séquence consensus de cet ADN satellite contient un enchaînement de 17 pb, la boîte CENP-B qui se lie spécifiquement aux protéines CENP du centromère. Région dépourvue d OR, la dernière à être répliquée. 5% du génome total Chez les Vertébrés, la région centromérique contient plusieurs centaines de kb d ADN satellite constitué de séquences d ADN répété. L ADN satellite est présent au niveau de la constriction primaire de tous les
5 chromosomes humains. La séquence consensus de cet ADN satellite contient un enchaînement nucléotidique de 17 paires de bases, appelé boîte CENP-B qui se lie spécifiquement à une protéine du centromère, la protéine CENP (CENtromere Protein). La nature particulière de l ADN centromérique est reflétée par la présence d au moins 7 protéines qui ne sont retrouvées nulle part ailleurs dans le chromosome. L une d elles est la CENP-A, très voisine de l histone H3. On pense qu elle peut remplacer celle-ci dans les nucléosomes centromériques sans que l on comprenne pour l instant les modifications qu elle y engendrerait. Une hypothèse découle de l observation que la région centromérique est la dernière région à être répliquée. L ADN de cette région serait ainsi dépourvu de séquences d origine de réplication, ce qui assure la réplication retardée de cette zone. Les séquences répétitives de l ADN seraient un moyen de garantir l absence d origines de réplication. c / Les minisatellites Longueur autour de 10 à 30 pb. Ex ADN télomérique Dispersés dans le génome se distinguent par la taille des motifs qui se répètent de 10 à 100 fois. Peut être en nb variable ds différentes positions chromosomiques et chez des membres distincts d une même espèce, répétitions en tandem : VNTR Les minisatellites forment des fragments de longueur voisine de 20 kb, avec des unités répétées d environ 10 à 30 pb. Un exemple de minisatellite est représenté par l ADN télomérique. Les télomères contiennent des alignements en tandem de séquences simples d ADN, qui ne codent pas d ARN ni de protéines mais qui ont des fonctions bien précises. (Cilié Tetrahymena : répétition de la séquence TTGGGG.) (Homme : répétition de la séquence TTAGGG.) Les répétitions observées au niveau des télomères jouent un rôle bien défini dans la réplication : on sait que le brin tardif atteint un endroit au-delà duquel le système d amorçage par l ARN ne peut plus fonctionner et il reste un fragment non polymérisé ce qui devrait aboutir à la formation d un chromosome tronqué. Une enzyme, la télomérase, ajoute les unités répétées simples aux extrémités du chromosome. La télomérase est une enzyme de la classe des transcriptases inverses. Elle fournit une courte molécule d ARN dont une partie sert de matrice à la polymérisation de l unité télomérique répétée. (Chez Tetrahymena, l ARN a la séquence 3 AACCCC-5 et sert de matrice pour l unité répétée 5 -TTGGGG-3 ) (Les télomères sont indispensables à l intégrité des chromosomes : en leur absence, les chromosomes deviennent instables et fusionnent avec d autres chromosomes ayant subi le même événement). En plus des minisatellites télomériques, certains génomes eucaryotes contiennent d autres fractions d ADN minisatellites, dont le rôle n a pas encore été élucidé. L ADN minisatellite peut être présent en nombre variable dans différentes positions chromosomiques et chez des membres distincts d une même espèce : on appelle ce type de répétitions, répétitions en tandem en nombre variable : VNTR : variable number tandem repeats. Chez l homme, les locus des VNTR sont des séquences de 1 à 5 kb contenant un nombre de répétition de 15 à 100 nucléotides. En raison de la variabilité du nombre de répétitions en tandem d un individu à l autre, l ensemble des fragments qui apparaissent sur l autoradiographie est hautement spécifique de chaque individu. Ces profils de bandes constituent les empreintes génétiques (DNA fingerprints). d / Les microsatellites Ce sont des fragments courts, inférieurs à 150 bp et les unités répétées 10 à 20 fois sont de 4 paires de bases ou moins. Chez l homme par exemple, l ensemble des microsatellites de motif CA représente au total 15 Mb, soit 0,5 % du génome.
6 La fonction de ces microsatellites reste inconnue. Le nombre de répétitions peut varier d un individu à l autre, ce qui en fait de bons marqueurs génétiques. La variabilité du nombre de copies résulte de glissements qui se produisent au cours de la réplication. Bilan microsatellites / minisatellites Les motifs répétés sont appelés microsatellites si la taille du motif est restreinte à 2 ou 3 nucléotides (par exemple (CA)n ou (CGA)n). Ils sont appelés minisatellites si les motifs répétés comportent un plus grand nombre de nucléotides. Celui-ci peut aller jusqu à quelques dizaines. Le nombre de motifs qui constituent les microsatellites et les minisatellites est variable. Ces variations de taille résultent soit de glissement de l ADN polymérase au cours de la réplication, soit de recombinaison entre deux motifs en positions différentes. 3 / Les séquences répétitives transposées Barbara Mc Clintock éléments sauteurs (Nobel en 83) 40% du génome chez l H Séquence d ADN capable de se déplacer de façon autonome dans le génome et de s y multiplier, ces séquences codent l ensemble des protéines nécessaires à leur déplacement. Une forte proportion du génome eucaryote est constituée d éléments répétitifs qui se sont propagés dans le génome en fabriquant des copies d eux-mêmes, capables de gagner d autres positions. Les éléments génétiques transposables sont encore plus courants dans les chromosomes eucaryotes que dans les chromosomes bactériens. 25 à 40 % des chromosomes de Mammifères sont constitués d éléments transposables qui se sont accumulés au cours de l évolution (homme 30 % d éléments transposables). La moitié des mutations spontanées observées chez la drosophile sont attribuées au déplacement et à l insertion d éléments transposables. Il existe différents types de séquences transposées a / Les transposons Se déplacent selon un mode couper-coller ou copier-coller s il y a réplication avant transposition. Se déplacent grâce à une transposase codée par le transposon. Ils portent à leur extrémité des courtes séquences répétées encadrant la séquence codant l enzyme. La transposition se fait sous forme d ADN et n implique pas de passage par l ARN (ce qui caractérise les rétrotransposons). Chez les Eucaryotes, les transposons sont moins fréquents que les rétrotransposons. Ils ont cependant été découverts sur le maïs et ont été reconnus chez la drosophile. Ils peuvent déterminer des transferts horizontaux de gènes entre espèces (exemple : élément mariner reconnu chez la drosophile et retrouvé chez d autres espèces dont l homme). Les transposons à ADN sont encore appelés transposons de type bactérien. Les transposons contiennent un ou plusieurs gènes encadrés par de courtes répétitions terminales inversées (RTI) à leurs extrémités. Un de ces gènes code pour une transposase qui opère les coupures de l ADN permettant la libération du transposon de son emplacement de départ et son insertion dans la nouvelle position. L élément P de la drosophile et l élément Ac/Ds du maïs sont des exemples bien étudiés de transposons à ADN b / Les rétrotransposons Classe 1 se déplacent selon un mode copier-coller et sont spécifiques des Eucaryotes ; Après transcription par l ARN pol II, l ARN est rétrotranscrit en ADN par la transcriptase réverse codée par le transposon. L ADN est intégré dans le génome par un intégrase. Transposons à LTR, importantes similitudes avec les rétrovirus. - les rétrotransposons de type rétroviral : LTR elements. Ceux-ci renferment généralement deux gènes (gag et pol) et sont encadrés par de longues répétitions terminales d une taille de quelques centaines de nucléotides, appelées LTR, Long Terminal Repeat). Les transcrits de ces gènes peuvent être recopiés en ARN grâce à une transcriptase inverse codée par le gène pol. L ADN ainsi obtenu peut lui-même aller s intégrer dans une nouvelle position

7 chromosomique grâce au gène gag. Le nom de transposon de type rétroviral est justifié par le fait qu ils se rapprochent de la famille des rétrovirus. Ceux-ci possèdent des gènes homologues. La seule différence est que, contrairement aux rétrovirus, les rétrotransposons ne possèdent pas de forme extracellulaire connue et ne sont pas infectieux. Exemple : élément Ty1 de la levure : 35 copies par génome. Ces éléments Ty provoquent des mutations par insertion dans différents gènes du chromosome. Exemple : élément de type copia chez la drosophile avec au moins 7 familles, de taille variant de 5 à 8,5 kb. Les membres de chaque famille apparaissent en 10 à 100 positions dans le génome de drosophile. Chaque membre porte une longue répétition terminale ainsi qu une courte répétition inversée. Certaines mutations classiques (couleur white-apricot de l oeil) résultent de l insertion d éléments de type copia Rétrotransposons sans LTR : Ce sont les transposons les plus fréquents rencontrés chez les Mammifères. Ces éléments présentent une région polya à une extrémité. - élément LINE : long interspersed nuclear elements : longs éléments dispersés. Les éléments LINE possèdent les gènes équivalents aux gènes gag et pol. Ils contiennent un gène codant pour la transcriptase réverse. Chez l homme, il existe des dizaines de milliers de copies d éléments LINE. Exemple : LINE-1 : 6,1 kb copies sous la forme la plus longue et plusieurs centaines de milliers de copies tronquées codent les enzymes nécessaires à la réverse transcription et à l intégration également - éléments SINE : short interspersed nuclear elements : courts éléments dispersés. Ils ne possèdent pas de gène codant la transcriptase réverse, ils ont donc perdu leur autonomie. La transposition se fait en empruntant les transcriptases synthétisées par d autres éléments. Le SINE le plus commun dans le génome humain est Alu, présent à près d un million de copies. Cette séquence répétitive est ainsi appelée car elle contient un site de coupure spécifique de l enzyme de restriction Alu. Le génome humain en contient des centaines de milliers de séquences entières ou tronquées, dispersées entre les gènes et même dans les introns qui constituent environ 5% de l ADN humain. La séquence Alu complète est longue d environ 200 nucléotides et est retrouvée en moyenne tous les 5 kb. Les transposons sont les acteurs de la dynamique du génome à deux niveaux : - immédiat, dans le déplacement d un élément - à long terme : une dynamique liée à la nature répétées de ces éléments Les conséquences dépendent de la région où se fait l insertion. - SINE : Bilan Organisation de l ADN eucaryote :
 qui est ce qui reste après que toutes les catégories précédentes ont été identifiées.")
8 Un grand nombre de séquences répétitives ont des fonctions inconnues et l on estime globalement qu environ 20 % du génome humain est constitué de séquences répétitives non fonctionnelles d un type ou d un autre. Une dernière classe d ADN est l ADN intercalaire (spacer) qui est ce qui reste après que toutes les catégories précédentes ont été identifiées. Cet ADN est mal étudié et sans fonction connue. Le concept de la sélection naturelle impliquait l élimination de l ADN non fonctionnel et on aurait pensé que cet ADN non fonctionnel était un fardeau génétique ne serait-ce qu en raison de l énergie nécessaire à sa réplication. Rôles hypothétiques : - «Ballast génétique» : donne aux chromosomes la masse critique requise pour une ségrégation efficace lors de la division cellulaire. - Séparation des éléments fonctionnels (les gènes), condition pour permettre une régulation efficace. II / L organisation des génomes extranucléaires Les génomes extranucléaires sont situés dans les mitochondries et les chloroplastes. A / Organisation générale ADN circulaires, souvent en plusieurs exemplaires et donc quantitativement importants dans la cellule eucaryote. Présence de nucléoïdes comme pour les chromosomes bactériens. Origine endosymbiotique. Des différences : code génétique et nombre et nature des ARNt (22/32 gènes). La plupart des chromosomes de ces organites apparaissent comme des molécules circulaires d ADN. Toutefois, on a reconnu aussi la présence de molécules linéaires. Les chromosomes d organites sont par ailleurs présents en de nombreux exemplaires. Chaque mitochondrie humaine contient de 2 à 10 molécules identiques avec un nombre de mitochondries par cellule variant d un type cellulaire à l autre. Les fibroblastes humains contiennent ainsi plusieurs centaines de molécules d ADNmt tandis que les ovocytes humains en possèdent près de Une cellule type de levure haploïde peut contenir de 1 à 45 mitochondries, chacune contenant de 40 à 100 molécules d ADN. Les chloroplastes contiennent des zones spécifiques qui réagissent fortement aux colorants de l ADN : on appelle ces régions des nucléoïdes (éléments retrouvés aussi dans les mitochondries).
9 Chaque chloroplaste de betterave contient de 4 à 8 nucléoïdes, eux-mêmes constitués individuellement de 4 à 18 molécules d ADN, soit environ 140 copies d ADN. B / Les génomes mitochondriaux Petite taille, organisation compacte (H 17 kb), faible nb de gènes (H 37 gènes) Les génomes mitochondriaux sont généralement de petite taille, avec une organisation compacte, les gènes étant situés près les uns des autres. Génome mitochondrial humain : pb (soit environ17 kb) ; Saccharomyces cerevisiae : 75 kb ; Arabidopsis thaliana : 367 kb Le nombre de gènes reste faible : Homme : 37 ; Saccharomyces cerevisiae : 35 ; Arabidopsis thaliana : 52 Deux fonctions essentielles du génome mitochondrial : - Il code certaines des protéines constituant le système de phosphorylation oxydative. - Il code des ARNt, ARNr et certaines protéines, utilisés dans la synthèse des protéines mitochondriales. Rmq : les autres protéines du système de phosphorylation oxydative sont codées par des gènes nucléaires. L ARNm est traduit hors de la mitochondrie par les ribosomes cytosoliques. Les protéines synthétisées sont transportées dans la mitochondrie et le système est assemblé dans la membrane mitochondriale interne. Le génome des mitochondries humaines comprend 13 gènes codant pour les protéines du système de phosphorylation oxydative, 22 gènes codant pour les ARNt mitochondriaux (et 2 gènes pour des ARNr). Ces ARNt assurent toute la traduction réalisée dans les mitochondries. Leur nombre est inférieur aux 32 ARNt requis pour traduire les ARNt fabriqués dans le noyau. Cette économie est permise par un flottement encore plus net de l appariement des anticodons des ARNt. Les codons représentatifs sont alors différents de ceux du code génétique nucléaire. On observe également des introns dans plusieurs gènes mitochondriaux de levure. C / Les génomes chloroplastiques Taille de 120 à 200 kb 136 gènes (4 ARNr, 31 ARNt, 90 protéines PS et CTE) Hérédité cytoplasmique, transmission horizontale des gènes, beaucoup son passé dans le génome nucléaire. La taille des molécules est comprise entre 120 et 200 kb. La molécule de Marchantia polymorpha (hépatique des fontaines) a pour taille 121 kb. Elle comprend 136 gènes codant entre autres quatre sortes d ARNr, 31 sortes d ARNt et environ 90 protéines. 20 protéines concernent les fonctions de photosynthèse et de transport des électrons. III / L expression du génome : la transcription Intervention des différentes molécules d ARN. - ARNm : traduits en protéines. - ARN fonctionnels qui ne sont jamais traduits en protéines et remplissent leurs fonctions uniquement en tant qu ARN. - Deux classes sont présentes dans toutes les cellules, procaryotes et eucaryotes : - ARN de transfert. - ARN ribosomaux.
10 - Trois classes sont spécifiques aux Eucaryotes : - petits ARN nucléaires : ARNsn : ils participent à l épissage des transcrits primaires en ARN messagers dans le noyau. Des protéines spécifiques se combinent aux ARNsn pour former de petites particules ribonucléosomiques (RNPsn) qui servent de plateformes aux réactions d épissage - petits ARN nucléolaires (ARNsno) : (ou guide de méthylation) : ils sont utilisés pour la maturation ou la modification chimique des ARNr. - petits ARN cytoplasmiques (ARNsc) : ils commandent le trafic des protéines. Ils garantissent par exemple que des polypeptides destinés à être sécrétés hors de la cellule soient insérés dans le réticulum endoplasmique. Ils interviennent aussi dans d autres processus : synthèse des télomères, inactivation du chromosome X. Les fonctions de tous les ADN et les ARN reposent sur deux éléments clés : - Les bases complémentaires dans les chaînes nucléotidiques simple-brin peuvent contracter des liaisons hydrogènes pour former des structures double-brin. - Des séquences de bases particulières dans les acides nucléiques simple ou double brin peuvent être reconnues par des protéines qui se fixent aux acides nucléiques. A / Mise en évidence expérimentale dʼun intermédiaire dʼarn Expérience de marquage avec des précurseurs radioactifs de l ARN : Uridine tritiée (chasse isotopique ou pulse chase) : apparition de l ARN d abord dans le noyau puis dans le cytoplasme. B / Les différentes étapes de la transcription Chaque segment transcrit d ADN est une unité de transcription. Chez les eucaryotes, une unité de transcription typique contient les informations d un seul gène et code pour une seule molécule d ARN ou une seule protéine (ou groupe de protéines apparentées si le transcrit d ARN initial est épissé de plusieurs façons et produit différents ARNm) (Rmq : Chez les bactéries, un groupe de gènes adjacents est souvent transcrit sous forme d une seule unité ; la molécule d ARN qui en résulte porte donc les informations de diverses protéines distinctes (opérons ; ARN polycistroniques)). 1 / L initiation Les régions de l ADN qui signalent l initiation de la transcription sont les promoteurs. L enzyme qui catalyse la transcription est l ARN polymérase. Chez les Procaryotes : un seul type d ARN polymérase transcrit tous les types d ARN. Chez les Eucaryotes, il y a trois ARN polymérases différentes : - l ARN polymérase I catalyse la synthèse des ARNr. - l ARN polymérase II catalyse la synthèse des ARNm. - l ARN polymérase III catalyse la synthèse des ARNt ainsi que les petites molécules d ARN nucléaires et cellulaires. Plusieurs autres différences : - Alors que l ARN polymérase bactérienne peut initier la transcription sur une matrice d ADN in vitro sans l aide d une autre protéine, l ARN polymérase des eucaryotes ne le peut pas et nécessite l intervention d un groupe important de protéines, les facteurs généraux de la transcription qui doivent s assembler au niveau du promoteur avec elle avant que ne commence la transcription. - L initiation de la transcription chez les eucaryotes doit faire face à l empaquetage de l ADN dans les nucléosomes. Le promoteur central est la région à laquelle se fixe l ARN polymérase II. Le promoteur central fait généralement référence à la région partant du site de début de la transcription, qui comprend la TATA box (boîte TATA), située environ 30 pb en amont du site d'initiation de la transcription. Ce promoteur central est incapable, seul, d'assurer une transcription
 fonctionne comme l'une de ces séquences agissant en cis, de même qu'un segment riche en GC.")

11 efficace. Certains éléments importants, les éléments proches du promoteur (ou éléments proximaux), sont distants de 100 à 200 ph du site d'initiation de la transcription. La CAT box (boîte CCAAT) fonctionne comme l'une de ces séquences agissant en cis, de même qu'un segment riche en GC. La région promotrice des Eucaryotes supérieurs 2 / L élongation L ARN est toujours synthétisé dans le sens 5-3. La chaîne s allonge par la formation d une liaison entre l extrémité 3 OH du brin en croissance et un nucléoside triphosphate, ce qui libère un pyrophosphate. L énergie est fournie par le clivage du triphosphate à haute énergie en monophosphate et diphosphate inorganique. NTP + (NMP)n --> (NMP)n+1 + PPi

12 3 / La terminaison (Chez les Procaryotes : deux mécanismes : - terminaison directe : séquence de terminaison sur l ADN comportant environ 40 pb se terminant par un segment riche en GC suivie de 6 A ou plus. Sur la séquence complémentaire d ARN, ce segment s organise en une boucle en épingle à cheveux, suivie d une succession de résidus U qui correspondent aux résidus A présents sur la matrice d ADN. La boucle en épingle à cheveux et la succession de résidus U semblent servir de signal pour la libération de l ARN polymérase et la terminaison de la transcription. - Intervention d un facteur protéique supplémentaire, rho, nécessaire à l ARN polymérase pour reconnaître les signaux de terminaison. Il n y a pas, dans ce cas, intervention de boucle en épingles à cheveux.) Chez les Eucaryotes, les phénomènes sont différents : il existe sur l ADN des signaux de coupure et de polyadénylation. Ces signaux sont transcrits sur l ARN lorsque l ARN polymérase II les traverse puis sont reconnus sous forme d ARN par une série de protéines de liaison à l ARN et d enzymes de maturation de l ARN (CstF : cleavage stimulation factor ; CPSF : cleavage and polyadenylation specificity factor). Lorsque ces deux facteurs sont fixés sur des séquences spécifiques de nucléotides de la molécule d ARN émergente, d autres protéines s assemblent à elles pour effectuer la maturation qui crée l extrémité 3 de l ARNm : l ARN m est coupé (intervention de facteurs complémentaires de coupure) puis une enzyme, la poly-a polymérase, ajoute un à un 200 nucléotides sur l extrémité 3 produite par clivage. Cette enzyme ne nécessite pas de matrice. L ARN polymérase II poursuit sa transcription au-delà du point de coupure puis de dissocie par un mécanisme mal compris. La partie d ARN en aval du site de clivage est dégradée dans le noyau de la cellule.


13 C / Contrôle et régulation de la transcription La transcription est un phénomène contrôlé. Les protéines régulatrices agissant en trans fonctionnent grâce à la spécificité des séquences d ADN reconnues, en se fixant à leurs séquences cibles régulatrices, qui, elles, agissent en cis. Action en cis : action de séquences adjacentes. Action en trans : action d un produit diffusible. 1 / Les séquences agissant en cis dans le contrôle et la régulation de la transcription L action de l'arn polymérase II fait intervenir une coopération entre de multiples éléments régulateurs agissant en cis. Trois classes d'éléments peuvent être distingués d'après leurs positions relatives : - Près du site d'initiation de la transcription se trouvent le promoteur central (la région à laquelle se fixe l ARN polymérase II) ainsi que des séquences agissant en cis proches du promoteur, auxquelles se fixent des protéines qui aident à leur tour à la fixation de l'arn polymérase II à son promoteur.
. Souvent, un enhancer ou un silenceur agit seulement dans un ou un petit nombre de types cellulaires chez un Eucaryote pluricellulaire.")
14 - D'autres éléments de séquence agissant en cis peuvent exercer leur fonction à des distances considérables : on appelle ces éléments des enhancers et des silenceurs (éléments répresseurs, silencers en anglais). Souvent, un enhancer ou un silenceur agit seulement dans un ou un petit nombre de types cellulaires chez un Eucaryote pluricellulaire. Les enhancers sont des séquences agissant en cis, qui peuvent fortement augmenter le taux de transcription à partir de promoteurs situés sur la même molécule d'adn. Ils peuvent donc activer (ou réguler de façon positive) la transcription. Les silenceurs exercent un effet inverse. Ce sont des séquences agissant en cis, auxquelles se fixent des répresseurs, inhibant ainsi les activateurs et réduisant le taux de transcription. Les enhancers et les silenceurs, comme les régions proches du promoteur, sont organisés en une succession de séquences agissant en cis, auxquelles se fixent des protéines régulatrices agissant en trans. Toutefois, ils se différencient des éléments proches du promoteur, par le fait qu'ils sont capables d'agir à distance, parfois à 50 kb ou plus, et d'exercer leur fonction en amont ou en aval du promoteur qu'ils contrôlent. Les enhancers et les silenceurs ont une structure très complexe. Les enhancers, eux-mêmes composés de multiples copies d'un élément d'adn auxquelles se fixent les protéines, sont courants. Des séquences d'adn différentes servent de sites cibles de reconnaissance pour des protéines régulatrices spécifiques, agissant en trans. Les mécanismes de l'action à distance : Comment des enhancers et des silenceurs situés à des milliers de paires de bases du site de transcription régulent-ils ce processus? La plupart des modèles de ce type d'action à distance comportent la formation d'une sorte de boucle dans l'adn (voir figure). Dans ce modèle, une boucle d'adn amène des protéines activatrices fixées sur des enhancers distants, à proximité de complexes protéiques associés aux séquences proximales du promoteur. Les promoteurs, les éléments proches des promoteurs et les éléments dont la fonction est indépendante de la distance, représentent donc tous des cibles de fixation pour différentes protéines qui agissent en trans en se fixant à l'adn 2 / Contrôle et régulation en trans de la transcription Un grand nombre de protéines régulatrices agissant en trans ont désormais été identifiées dans les cellules eucaryotes. Comme leurs équivalents chez les procaryotes, ces protéines régulatrices agissent en se fixant à des séquences cibles spécifiques d'adn.
15 Les protéines régulatrices qui se fixent au promoteur central et aux éléments proches du promoteur aident l'arn polymérase II à démarrer la transcription et forment avec celle-ci un complexe d'initiation (voir figure). Plusieurs complexes de facteurs de transcription différents (complexes TFII) interagissent avec l ARN polymérase II. Par exemple, le complexe TFIID est constitué d'une protéine se fixant à la TATA box (TBP pour TATA box-binding protein en anglais) et de plus de huit autres sous-unités. On appelle souvent les complexes TFII, des facteurs généraux de la transcription, car ils constituent le minimum nécessaire pour que l ARN polymérase II puisse débuter la transcription (généralement à un taux très faible) au niveau d'un promoteur. Les CAT et GC box sont reconnues par d'autres protéines de liaison à l'adn. Certaines des protéines qui se fixent à des éléments dont la fonction est indépendante de la distance ont également été identifiées. La protéine codée par le gène GCN4 de levure est un exemple de protéine agissant en trans qui se fixe à des enhancers. Elle se fixe à des enhancers appelés séquences activatrices en amont (UAS pour upstream activating sequences en anglais). GCN4 active la transcription de nombreux gènes de levure qui codent les enzymes des voies de biosynthèse des acides aminés. En réponse à une carence en acides aminés, la concentration de la protéine GCN4 augmente, et entraîne à son tour une augmentation du taux d'expression des gènes de biosynthèse des acides aminés. Les UAS reconnues par GCN4 contiennent l'élément de séquence de reconnaissance principal, ATGACTCAT. Structure des protéines régulatrices : Les protéines régulatrices de gènes reconnaissent de courts segments de la double hélice d ADN d'une séquence définie et déterminent ainsi quel gène, parmi les milliers de la cellule, sera transcrit. Des milliers de protéines régulatrices ont été identifiées dans un large éventail d'organismes. Même si chacune de ces protéines a une caractéristique propre, la plupart se lient à l ADN sous forme d'homodimères ou d'hétérodimères et reconnaissent l ADN par une des quelques conformations structurales. Les conformations les plus communes comprennent l'hélice-coude-hélice, les homéodomaines, les fermetures Éclair à leucine, les hélice-boucle-hélice et les doigts à zinc de divers types. La séquence précise en acides aminés qui est repliée en une de ces conformations détermine la séquence particulière d ADN reconnue. L'hétérodimérisation augmente la variété des séquences d ADN qui peuvent être reconnues par les protéines régulatrices. Il existe des techniques puissantes qui utilisent la spécificité des protéines régulatrices pour les séquences d ADN pour identifier et isoler ces protéines, les gènes qui codent pour elles, les séquences d ADN qu'elles reconnaissent et les gènes qu'elles régulent. D / Transcription et structure du matériel génétique 1 / Remodelage de la chromatine On pourra s intéresser plus précisément aux états de la chromatine et notamment aux effets de la méthylation des histones (sur les lysines et les arginines). La méthylation des histones joue un rôle dans la mise en place de l hétérochromatine. On rappellera que des modifications de la structure chromatinienne et des nucléosomes peuvent être nécessaires avant l initiation de la transcription : ces modifications portent le nom de remodelage de la chromatine, processus actif réalisé par des complexes de remodelage, comprenant de 2 à 12 protéines, dont une hélicase. De manière simplifiée, les complexes de remodelage peuvent agir selon deux modes : - dans le premier cas, le remodelage consiste en un déplacement des nucléosomes, ce qui libère l ADN sur une courte portion et permet la fixation des facteurs de transcription. - dans le second cas, les complexes de remodelage modifient la torsion de l ADN sur le nucléosome. La distorsion entraîne une modification de la structure de la double hélice, qui rend les bases de l ADN plus accessibles aux facteurs de transcription. 2 / Modifications de la structure primaire de l ADN
16 Certaines séquences CG de l ADN peuvent être méthylées, la méthylation ne portant que sur les cytosines. Un ensemble de résultats expérimentaux ont montré que la méthylation des cytosines situées dans la région en 5 non transcrite des gènes était associée à une diminution de leur activité transcriptionnelle. L ADN des cellules cancéreuses, dont la transcription est très active, est à l inverse largement hypométhylé. Le chromosome X inactivé est globalement hyperméthylé, ce qui peut être mis en relation avec l inactivation de la plupart de ses gènes. L effet inhibiteur de la méthylation sur la transcription résulte d au moins deux types de mécanismes possibles : - la méthylation modifie la forme et les caractères physicochimiques de la molécule d ADN, ce qui modifie la reconnaissance des séquences par les facteurs de transcription. - L autre mécanisme est lié à l intervention de protéines MBD (Methyl Binding Domain), protéines qui contiennent un domaine de fixation à l ADN méthylé. Leur fixation interdit l accès des facteurs de transcription à l ADN. Elles sont par ailleurs associées à d autres molécules comme les histone-désacétylases qui ont un effet inhibiteur sur la transcription. Bilan sur le contrôle et la régulation de la transcription : La transcription de chaque gène est activée et inactivée dans les cellules par des protéines régulatrices. Certaines de ces protéines se fixent sur des séquences d ADN spécifiques près du point de départ de l ARN polymérase. La souplesse de l'hélice d ADN, cependant, permet également à des protéines fixées sur des sites distants de modifier l activité de l ARN polymérase au niveau du promoteur. Cette action à distance, sur des séquences situées à des milliers de paires de nucléotides du promoteur, est extrêmement fréquente dans les cellules eucaryotes. Les activateurs et les répresseurs eucaryotes agissent selon un large éventail de mécanismes - qui provoquent généralement la modification locale de la structure de la chromatine, l'assemblage des facteurs généraux de transcription au niveau du promoteur et le recrutement de l ARN polymérase. La régulation de l expression des gènes eucaryotes supérieurs est ainsi beaucoup plus complexe que celle des gènes procaryotes, en relation avec la grande taille du génome et la grande variété des types cellulaires formés. La région de contrôle du gène eve de drosophile, par exemple, s'étend sur paires de nucléotides d ADN et présente des sites de liaison pour plus de 20 protéines régulatrices. Certaines de ces protéines sont des répresseurs de la transcription, d'autres des activateurs de la transcription. Ces protéines se fixent sur des séquences régulatrices organisées en une série de modules régulateurs répartis le long de l ADN et provoquent ensemble le bon patron temporel et spatial d'expression génique. Les gènes eucaryotes et leurs régions contrôles sont souvent entourés d'éléments isolateurs, séquences d ADN reconnues par des protéines qui empêchent la diaphonie entre les gènes régulés indépendamment. L assemblage moléculaire contrôlant la transcription apparaît ainsi constitué de quatre types de molécules. Les facteurs généraux de transcription sont essentiels à la transcription mais ne peuvent par euxmêmes augmenter ou abaisser le taux de transcription. Cet effet revient aux protéines régulatrices appelées activateurs ou répresseurs. Les activateurs et sans doute les répresseurs, communiquent avec les facteurs généraux par le biais de co-activateurs, des protéines complexées avec la protéine qui se fixe à la TATA box, le premier des facteurs élémentaires de transcription à se fixer au promoteur central.
. - excision des introns et épissage des exons.")
17 IV / La maturation des ARN pré-messagers La maturation des ARN correspond aux phénomènes suivants : - coiffage de l extrémité 5 par un résidu 7-méthylguanosine. - polyadénylation de l extrémité 5 (longueur de 150 à 200 résidus). - excision des introns et épissage des exons. La présence des extrémités particulières des ARNm est nécessaire à leur exportation hors du noyau. Certaines de ces modifications de l ARN qui affectent le transcrit initial d ARN (par exemple, celles impliquées dans l'épissage de l ARN) sont surtout effectuées par de petites molécules particulières d ARN (voir plus loin). A / Coiffage et polyadénylation de lʼarn 1 / Le coiffage de l extrémité 5 Le coiffage est la première modification du pré-arnm. Dès que l ARN polymérase a produit 25 nucléotides environ, la terminaison 5 de la nouvelle molécule est modifiée par addition de la coiffe. Cette modification fait intervenir trois enzymes (phosphatase, guanyl-transférase, méthyl-transférase). La coiffe permet de distinguer l ARN des autres types d ARN de la cellule (les ARN polymérases I et III produisent des ARN sans coiffe). Dans le noyau, la coiffe s unit à un complexe protéique appelé CBC (cap-binding complex) qui facilite la maturation correcte de l ARN puis son exportation. Cette coiffe joue aussi un rôle important dans la traduction cytosolique de l ARNm.
18 2 / La polyadénylation La polyadénylation s effectue lors de la terminaison de la transcription (voir plus haut).
19 CPSF : cleavage and polyadenylation specificity factor Cstf : cleavage stimulating factor B / Épissage des ARN prémessagers (pré-arnm) Les séquences d introns et d exons sont transcrites en ARN. Les séquences d introns sont retirées de l ARN néosynthétisé lors du processus d épissage. 1 / Les mécanismes moléculaires L épissage est assuré par un complexe moléculaire, le splicéosome. Celui-ci associe au moins 5 types d ARN de petite taille (moins de 200 nucléotides). Appelés ARNsn, chacun forme un complexe avec au moins 7 sous-unités protéiques pour former une RNPsn (small nuclear ribonucleoprotein). Ces RNPsn forment le coeur du splicéosome, gros assemblage de plus de 50 protéines qui effectue l épissage. Chaque épissage s effectue par deux réactions séquentielles de transfert d un phosphoryle qui réunissent deux exons tout en enlevant un intron, formant un lasso. L épissage est consommateur d ATP : le splicéosome utilise l hydrolyse de l ATP pour effectuer une série complexe de réarrangements ARN- ARN. L élimination de la séquence d intron fait intervenir trois positions sur l ARN : le site d épissage 5, le site d épissage 3 et le point d embranchement de la séquence d intron qui forme la base du lasso
20 excisé. Ces trois sites ont chacun une séquence consensus en nucléotides, similaire d un intron à l autre. Chaque séquence présente cependant suffisamment de variations pour qu il soit très difficile de repérer les signaux d épissage d un génome.

21 2 / L épissage alternatif Lorsqu il existe différentes possibilités d épissage dans diverses positions du transcrit, un seul gène peut produire de nombreuses protéines différentes. Épissage alternatif de constitution :
22 Dans certains cas, l épissage alternatif se produit lorsqu il existe une ambiguïté dans la séquence d introns : le mécanisme standard du splicéosome ne distingue pas nettement entre plusieurs appariements alternatifs des sites d épissage : les asociations se font au hasard. Dans ce cas, les différentes versions de la protéine codée par le gène sont fabriquées dans toutes les cellules où le gène s exprime. Épissage alternatif régulé : Dans les cas les plus simples, l épissage régulé sert à passer de la production d une protéine non fonctionnelle à la production d une protéine fonctionnelle. Cet épissage permet aussi de produire différentes versions d une protéine dans différents types cellulaires, selon les besoins de la cellule. La tropomyosine est par exemple produite sous des formes spécialisées dans différents types cellulaires. La régulation de l épissage de l ARN peut être soit négative par l intermédiaire d une molécule régulatrice qui empêche la machinerie d épissage d accéder à un site d épissage particulier, soit positive par l intermédiaire d une protéine régulatrice qui dirige la machinerie d épissage sur un site d épissage négligé jusqu à alors. L épissage d une molécule de pré-arnm peut ainsi apparaître comme un équilibre entre des sites d épissage compétitifs, équilibre qui peut être modifié par des protéines régulatrices. Exemple du gène morcelé de l ovalbumine : les différentes étapes de la maturation de l ARNm. 3 / La signification de l épissage Possibilité d épissage alternatif. L épissage est un phénomène précis : si des erreurs d épissage de l ARN étaient fréquentes, elles endommageraient sérieusement la cellule en entraînant des dysfonctionnements protéiques. En cas d erreur d épissage, la cellule possède d ailleurs un dispositif de sécurité qui élimine les ARN mal épissés. Il peut sembler que l élimination d un grand nombre d introns soit du gaspillage. L arrangement exon intron peut toutefois faciliter l émergence de protéines nouvelles. La présence de nombreux introns permet à la recombinaison génétique d associer plus facilement les exons de différents gènes. Cette hypothèse est confortée par le fait qu un grand nombre de protéines actuelles ressemblent à des patchworks composés à partir d un ensemble commun de domaines protéiques.

23 L épissage a un avantage actuel : les transcrits de beaucoup de gènes (60 % chez l homme) sont épissés de diverses manières, ce qui permet de produire un ensemble correspondant de protéines à partir d un même gène. Ceci augmente le potentiel de codage du génome. Rmq : L auto-épissage de l ARN : il existe de nombreux exemples de molécules d ARN capables de catalyser l épissage de leurs introns sans l aide de protéines (mis en évidence dans des ARNr chez le cilié Tetrahymena). Ce fut la première démonstration d une propriété catalytique spécifique pour une molécule d ARN : les ARN pourvus d une activité catalytique sont appelés des ribozymes. Erreurs très dommageables Émergences de protéines nouvelles? La présence de nombreux introns permet à la recombinaison d associer plus facilement les exons de différents gènes (beaucoup de protéines ressemblent à des patchworks composés à partir d un ensemble commun de domaines). 4 / Épissage alternatif et définition du gène Évolution de la notion de gène : - Approches de génétique classique : Région du génome qui se sépare en une seule unité lors de la méiose et qui donne naissance à un caractère phénotypique défini : Beadle et Tatum (Neurospora) un gène une enzyme : Yanofsky : la colinéarité - Années 60, un segment d ADN transcrit en ARN et qui code pour une seule chaîne polypeptidique ou transcrit en ARN structural (t ou r...). - Notion de 1 gène-1 ARN remise en cause lors de la découverte de l épissage alternatif Gène : toute séquence d ADN transcrite en un ARN structural ou sous forme d une seule unité ARN codant pour un groupe de chaînes polypeptidiques très apparentées (protéines isoformes) Nouvelles techniques d approches : Sondes, blotting, analyse des transcriptomes par hybridation sur puces à ADN on conduit à la définition actuelle : Un gène est un ensemble de séquences génomiques qui sont potentiellement recouvrantes, qui codent des molécules fonctionnelles (protéines ou ARN) et qui contiennent les signaux nécessaires à son expression. Schéma d un gène eucaryote codant une protéine : Structure d un ARNm d Eucaryote :
 est généralement prédominant.")
 l'atténuation des transcrits d ARN par leur terminaison prématurée, (2) la sélection de sites d'épissage alternatif, (3) le contrôle de la formation de l'extrémité 3' par")
24 Les différentes étapes de l expression du génome chez les Eucaryotes : Les cellules eucaryotes peuvent contrôler l'expression génique par différents processus. La plupart des gènes peuvent ainsi être régulés à de multiples niveaux, même si le contrôle de l'initiation de la transcription (contrôle transcriptionnel) est généralement prédominant. Certains gènes sont cependant transcrits à vitesse constante et activés ou inactivés uniquement par des processus régulateurs posttranscriptionnels. Ces processus incluent : (1) l'atténuation des transcrits d ARN par leur terminaison prématurée, (2) la sélection de sites d'épissage alternatif, (3) le contrôle de la formation de l'extrémité 3' par le clivage et l'addition du poly-a, (4) l'édition des ARN (modification de la séquence nucléotidique d un transcrit d ARNm une fois transcrit), (5) le contrôle du transport du noyau au cytosol, (6) la localisation de l ARNm dans des sites cellulaires particuliers, (7) le contrôle de l'initiation de la traduction, (8) la dégradation régulée des ARNm. La plupart de ces processus de contrôle nécessitent la reconnaissance de séquences ou de structures spécifiques situées sur la molécule d ARN à réguler. Cette reconnaissance s'effectue soit par des protéines régulatrices soit par des molécules d ARN régulatrices. V/ La traduction des ARN messagers (le protéome) Le résultat final de l expression du génome est le protéome, ensemble des protéines fonctionnelles de la cellule. A / Le code génétique Le code génétique est le système de correspondance entre les séquences de nucléotides et les séquences d acides aminés. Ce code a été déchiffré au début des années 1960.
25 Chaque groupe de trois nucléotides consécutifs de l ARN messager constitue un codon. Il spécifie soit un acide aminé soit un signal d arrêt de la traduction (trois codon stop, UAA, UAG et UGA). Le code génétique est redondant : certains acides aminés sont spécifiés par plusieurs triplets. Le code génétique est utilisé par tous les organismes actuels : il est ainsi considéré comme universel. Toutefois, des différences ont été observées : - exemple : Candida albicans : CUG : sérine alors qu habituellement leucine. - dans l expression des génomes mitochondriaux (mitochondries de mammifères : AUA signifie méthionine alors qu habituellement ce codon est traduit en isoleucine. B / Le rôle des ARNt et des aminoacyl-arnt synthétases 1 / Structure et propriétés des ARNt Les molécules d ARNt comprennent 80 nucléotides ; elles présentent une structure tridimensionnelle précise, en forme de trèfle. Cette structure est stabilisée par quatre courts segments d ARN appariés. Cette structure en trèfle subit d autres repliements qui conduisent à une structure spatiale en L. Deux régions non appariées des ARNt ont une grande importance fonctionnelle : - L anticodon est un groupe de trois nucléotides consécutifs qui s apparient au codon complémentaire de la molécule d ARNm. - Une courte séquence au niveau de l extrémité 3 est le lieu de fixation de l acide aminé. Structure secondaire d un ARNt La redondance du code génétique peut résulter du fait : - qu il existe plusieurs ARNt pour chaque acide aminé. - que certaines molécules d ARNt peuvent former des appariements de bases avec plusieurs codons. Certains ARNt ne forment ainsi qu un appariement précis pour les deux premières positions du codon. La troisième position du codon, dite position flottante, peut s apparier avec différentes bases de l anticodon. Cet appariement de bases flottant permet d adapter les 20 acides aminés sur les 61 codons avec seulement 31 types de molécules d ARNt. 2 / Le rôle des aminoacyl-arnt synthétases La reconnaissance et la fixation d un acide aminé sur l ARNt correspondant sont le fait des aminoacyl- ARNt synthétases. Dans la plupart des cellules, il y a une synthétase différente pour chaque acide aminé. Formation des complexes aminoacyl-arnt synthétases.

26 Les synthétases présentent une double spécificité essentielle, sur laquelle repose la correspondance acide aminé déterminé anticodon. Ce rôle majeur a été démontré expérimentalement : si on modifie expérimentalement l acide aminé après sa fixation sur l ARNt correspondant, cet acide aminé modifié est partout inséré en place de l acide aminé initial. La traduction met donc en jeu deux types d adaptateurs agissant de manière séquentielle, et de même importance. La fixation de l acide aminé sur l ARNt requiert l hydrolyse d ATP (formation préalable d un acide aminé activé). L énergie de liaison entre l acide aminé et l ARNt sera ensuite utilisée lors de la synthèse protéique pour enchaîner de manière covalente l acide aminé sur la chaîne polypeptidique en croissance. 3 / Le rôle des ribosomes La synthèse des protéines est réalisée au niveau des ribosomes, machineries catalytiques associant plusieurs molécules d ARNr à plus de 50 protéines. Un ribosome est constitué de deux sous-unités : - la petite sous-unité assure la bonne correspondance entre le codon de l ARNm et l ARNt correspondant. - la grande sous-unité catalyse la formation de la liaison peptidique. Les ARNr apparaissent déterminer la structure globale du ribosome, sa capacité à positionner l ARNt sur l ARNm et sa capacité catalytique pour la formation des liaisons peptidiques. (ce qui conduit à considérer certains ARNr comme des ribozymes). Le rôle principal des protéines semble être de stabiliser le coeur d ARNr, tout en permettant les variations de conformation des ARNr nécessaires à leur action de catalyse. Petite sous-unité des ribosomes eucaryotes : ARNr 18 S (1900 nucléotides) et 33 protéines Grande sous-unité des ribosomes eucaryotes : ARNr 5S (120 nucléotides), 28S (4700 nucléotides), 5,8S (160 nucléotides) et 49 protéines. Un ribosome contient 4 sites de liaisons aux molécules d ARN : - un site pour les molécules d ARNm trois sites pour les molécules d ARNt : sites A (aminoacyl-arnt), P (peptidyl-arnt) et E (exit). Au cours du processus de synthèse, il semble qu il n y ait pas plus de deux sites occupés par une molécule d ARNt à un instant donné C / Les étapes de la traduction Le site où débute la synthèse est de grande importance, toute erreur à ce niveau pouvant conduire à un décalage du cadre de lecture.
27 La traduction d un ARNm commence par le codon AUG, avec un ARNt spécifique qui initie la traduction. Cet ARNt initiateur transporte toujours l acide aminé méthionine. Toutes les protéines néosynthétisées possèdent la méthionine comme premier acide aminé au niveau de leur extrémité N- terminale. La méthionine est généralement retirée ultérieurement par une protéase spécifique. L ARNt initiateur possède une séquence nucléotidique différente de celles des autres ARNt qui transportent normalement la méthionine. - L ARNt initiateur est placé dans la petite sous-unité ribosomique avec d autres protéines appelées facteurs d initiation des eucaryotes ou eif. Parmi tous les aminoacyl-arnt de la cellule, seul l ARNt initiateur chargé de méthionine peut se fixer sur la petite sous-unité du ribosome alors que celui-ci n est pas complet. - La petite sous-unité ribosomique se fixe à l extrémité 5 d une molécule d ARNm qui est reconnue grâce à sa coiffe 5 et à deux facteurs d initiation qui s y sont fixés. - La petite sous-unité ribosomique se déplace alors dans le sens 5-3 jusqu au premier codon AUG. Ce mouvement est facilité par d autres facteurs d initiation qui agissent comme des hélicases actionnées par l ATP. - La traduction débute à ce premier codon AUG : les facteurs d initiation se dissocient de la petite sous-unité pour laisser la place à la grande sous-unité. L ARNt initiateur est alors lié au site P et laisse le site A vacant.
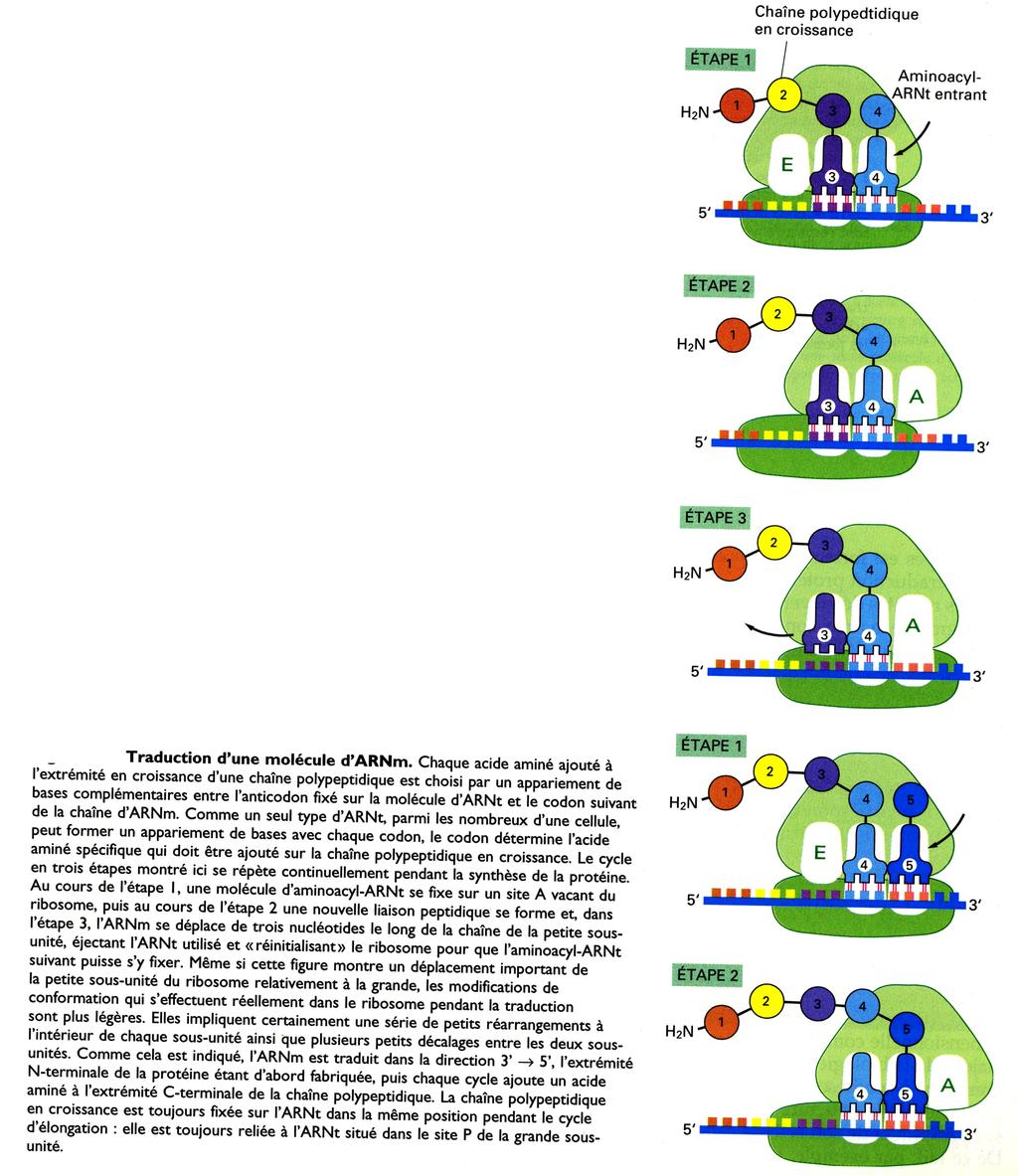
28 1 / L initiation

29 2 / L élongation La chaîne protéique est synthétisée par étapes, de son extrémité N-terminale à son extrémité C-terminale. L élongation fait intervenir un ensemble de processus cycliques : - fixation d une nouvelle molécule d aminoacyl-arnt sur le site A. - libération de l extrémité carboxyle de la chaîne polypeptidique jusqu alors liée à l ARNt fixé au site P. - Liaison de cette extrémité carboxyle avec le groupement amine libre de l acide aminé lié à l ARNt situé au site P. Cette étape correspond à la formation d une liaison peptidique : elle est catalysée par l activité catalytique de la peptidyl-transférase contenue dans la grande sous-unité ribosomique. - Cette étape s accompagne de plusieurs modifications de conformation du ribosome qui déplacent les deux ARNt dans les sites E et P de la grande sous-unité.
.")

30 Détail de la formation de la liaison peptidique Rmq : la synthèse protéique est un exemple de polymérisation par l avant : chaque monomère porte une liaison riche en énergie (monomère activé) qui sera utilisée pour ajouter le monomère suivant (autre exemple : acides gras). Les acides nucléiques, les polysaccharides sont le siège d une polymérisation par l arrière, chaque monomère portant une liaison riche en énergie (monomère activé) utilisée à sa propre incorporation. La traduction est un compromis entre deux contraintes opposées : l exactitude et la vitesse. Eucaryotes : vitesse d élongation de l ordre de 2 acides aminés ajoutés par seconde (Procaryotes, 20 acides aminés par seconde) avec 1 erreur/ acides aminés reliés. L élongation fait intervenir deux facteurs d élongation, chacun hydrolysant un GTP en GDP. L aminoacyl-arnt entrant dans le site est dans un premier temps lié avec un facteur d élongation (EF-Tu) porteur d un GTP. Cet aminoacyl-arnt s apparie transitoirement au niveau du site A : fixation sur un site de liaison initial appelé état hybride A/T. L appariement codon-anticodon déclenche l hydrolyse du GTP par l EF-Tu, ce qui provoque la dissociation de ce dernier du complexe aminoacyl-arnt : celui-ci entre pleinement dans le site A (site A/A). Ce facteur d élongation introduit donc deux courts délais entre l appariement des bases codonanticodon et l élongation de la chaîne polypeptidique : ces retards permettent sélectivement aux ARNt incorrects liés de ressortir du ribosome avant que l étape irréversible d élongation de la chaîne ne se produise. Le premier délai est le temps nécessaire à l hydrolyse du GTP. La vitesse d hydrolyse est plus rapide si la paire codon-anticodon est correcte ce qui donne l opportunité plus longue de dissociation pour une paire incorrecte. Le deuxième retard se produit entre la dissociation du facteur d élongation et la mise en place complète de l ARNt dans le site A.
31 Ces mécanismes augmentent considérablement la précision de la traduction. Un second facteur d élongation EF-G sous sa forme liée au GTP entre ensuite dans le ribosome et se fixe à l intérieur ou près du site A, sur la grande sous-unité, ce qui accélère le déplacement des deux ARNt fixés qui passent dans les états hybrides A/P et P/E. Le contact avec le ribosome stimule l activité GTPasique de l EF-G, dont la conformation est alors modifiée : cette modification déplace l ARNt lié dans son état hybride A/P vers le site P et avance le site de traduction d un codon. À chaque cycle, on considère ainsi qu une molécule d ARNt occupe 6 sites différents : site de liaison initial (état hybride A/T), site A/A, état hybride A/P, site P/P, état hybride P/E et le site E. On pense que chaque ARNt se visse par un système de cliquet dans ces positions, subissant des rotations le long de son grand axe à chaque changement de localisation. 3 / La terminaison La terminaison est signalée par la présence d un codon Stop (UAA, UAG et UGA). Ils ne sont pas reconnus par un ARNt et ne spécifient aucun acide aminé. Une protéine, appelée facteur de libération se fixe alors sur le site A, ce qui conduit la peptidyltransférase à catalyser l addition d une molécule d eau sur le peptidyl-arnt. Cette réaction libère l extrémité carboxyle de la chaîne polypeptidique en croissance de son attache sur la molécule d ARNt. Les facteurs de libération fournissent un exemple de mimétisme moléculaire, dans lequel un type de macromolécule a une forme qui ressemble à celle d une molécule chimiquement non apparentée. Dans ce cas, la structure tridimensionnelle des facteurs de libération, protéiques, présente une ressemblance surprenante avec la forme d une molécule d ARNt. Pendant la traduction, le polypeptide naissant se déplace à travers un grand tunnel rempli d eau (10 nm x 1,5 nm) de la grande sous-unité du ribosome. Cette structure associe des surfaces hydrophobes encastrées sur des surfaces hydrophiles et constitue une sorte de «revêtement de téflon» sur lequel la chaîne polypeptidique glisse aisément. La structure tridimensionnelle de la protéine est acquise ultérieurement. Des pistes de conclusion On pourra notamment penser : - à envisager les principales directions de recherche visant à améliorer notre connaissance du génome et de son expression. Ceci conduit notamment à développer quelques aspects relatifs au génie génétique. - à présenter des applications de la connaissance du génome dans les domaines de la médecine. à envisager la transmission du génome, avec des particularités relatives à l activité des gènes ( ex : transmission épigénétique
CHAPITRE 3 LA SYNTHESE DES PROTEINES
 CHAITRE 3 LA SYNTHESE DES ROTEINES On sait qu un gène détient dans sa séquence nucléotidique, l information permettant la synthèse d un polypeptide. Ce dernier caractérisé par sa séquence d acides aminés
CHAITRE 3 LA SYNTHESE DES ROTEINES On sait qu un gène détient dans sa séquence nucléotidique, l information permettant la synthèse d un polypeptide. Ce dernier caractérisé par sa séquence d acides aminés
VI- Expression du génome
 VI- Expression du génome VI-1.- EXPRESSION DU GÉNOME- PRINCIPES GÉNÉRAUX DOGME CENTRAL Les gènes et l information génétique sont conservés sous forme d acides nucléiques La perpétuation à l identique de
VI- Expression du génome VI-1.- EXPRESSION DU GÉNOME- PRINCIPES GÉNÉRAUX DOGME CENTRAL Les gènes et l information génétique sont conservés sous forme d acides nucléiques La perpétuation à l identique de
TD de Biochimie 4 : Coloration.
 TD de Biochimie 4 : Coloration. Synthèse de l expérience 2 Les questions posées durant l expérience 2 Exposé sur les méthodes de coloration des molécules : Générique Spécifique Autres Questions Pourquoi
TD de Biochimie 4 : Coloration. Synthèse de l expérience 2 Les questions posées durant l expérience 2 Exposé sur les méthodes de coloration des molécules : Générique Spécifique Autres Questions Pourquoi
Contrôle de l'expression génétique : Les régulations post-transcriptionnelles
 Contrôle de l'expression génétique : Les régulations post-transcriptionnelles http://perso.univ-rennes1.fr/serge.hardy/ utilisateur : biochimie mot de passe : 2007 L'ARNm, simple intermédiaire entre le
Contrôle de l'expression génétique : Les régulations post-transcriptionnelles http://perso.univ-rennes1.fr/serge.hardy/ utilisateur : biochimie mot de passe : 2007 L'ARNm, simple intermédiaire entre le
Cellules procaryotes Service histologie Pr.k.mebarek
 Cellules procaryotes Service histologie Pr.k.mebarek I) Les cellules procaryotes II) Les cellules eucaryotes o 1) Caractéristiques générales des cellules eucaryotes o 2) Organisation des cellules eucaryotes
Cellules procaryotes Service histologie Pr.k.mebarek I) Les cellules procaryotes II) Les cellules eucaryotes o 1) Caractéristiques générales des cellules eucaryotes o 2) Organisation des cellules eucaryotes
ULBI 101 Biologie Cellulaire L1. Le Système Membranaire Interne
 ULBI 101 Biologie Cellulaire L1 Le Système Membranaire Interne De la nécessité d un SMI Le volume augmente comme le cube de la dimension linéaire, alors que la surface n'est augmentée que du carré Une
ULBI 101 Biologie Cellulaire L1 Le Système Membranaire Interne De la nécessité d un SMI Le volume augmente comme le cube de la dimension linéaire, alors que la surface n'est augmentée que du carré Une
Chapitre 7 : Structure de la cellule Le noyau cellulaire
 UE2 : Structure générale de la cellule Chapitre 7 : Structure de la cellule Le noyau cellulaire Professeur Michel SEVE Année universitaire 2010/2011 Université Joseph Fourier de Grenoble - Tous droits
UE2 : Structure générale de la cellule Chapitre 7 : Structure de la cellule Le noyau cellulaire Professeur Michel SEVE Année universitaire 2010/2011 Université Joseph Fourier de Grenoble - Tous droits
Module 5 La maturation de l ARN et le contrôle post-transcriptionnel chez les eucaryotes
 Module 5 La maturation de l ARN et le contrôle post-transcriptionnel chez les eucaryotes Où trouver l'information complémentaire? MCB -11, GVII-5, 22, 23. La maturation des ARNm chez les eucaryotes Les
Module 5 La maturation de l ARN et le contrôle post-transcriptionnel chez les eucaryotes Où trouver l'information complémentaire? MCB -11, GVII-5, 22, 23. La maturation des ARNm chez les eucaryotes Les
Dr E. CHEVRET UE2.1 2013-2014. Aperçu général sur l architecture et les fonctions cellulaires
 Aperçu général sur l architecture et les fonctions cellulaires I. Introduction II. Les microscopes 1. Le microscope optique 2. Le microscope à fluorescence 3. Le microscope confocal 4. Le microscope électronique
Aperçu général sur l architecture et les fonctions cellulaires I. Introduction II. Les microscopes 1. Le microscope optique 2. Le microscope à fluorescence 3. Le microscope confocal 4. Le microscope électronique
Contrôle de l'expression génétique :
 Contrôle de l'expression génétique : Les régulations post-transcriptionnelles L'ARNm, simple intermédiaire entre le génome et les protéines? gène protéine L'ARNm, simple intermédiaire entre le génome et
Contrôle de l'expression génétique : Les régulations post-transcriptionnelles L'ARNm, simple intermédiaire entre le génome et les protéines? gène protéine L'ARNm, simple intermédiaire entre le génome et
Travaux dirigés de Microbiologie Master I Sciences des Génomes et des Organismes Janvier 2015
 Andrew Tolonen [email protected] Travaux dirigés de Microbiologie Master I Sciences des Génomes et des Organismes Janvier 2015 A- Généralités I- La vie sur terre telle que nous la connaissons ne
Andrew Tolonen [email protected] Travaux dirigés de Microbiologie Master I Sciences des Génomes et des Organismes Janvier 2015 A- Généralités I- La vie sur terre telle que nous la connaissons ne
Séquence 2. L expression du patrimoine génétique. Sommaire
 Séquence 2 L expression du patrimoine génétique Sommaire 1. La synthèse des protéines 2. Phénotypes, génotypes et environnement Synthèse de la séquence 2 Exercices de la séquence 2 Glossaire des séquences
Séquence 2 L expression du patrimoine génétique Sommaire 1. La synthèse des protéines 2. Phénotypes, génotypes et environnement Synthèse de la séquence 2 Exercices de la séquence 2 Glossaire des séquences
Les OGM. 5 décembre 2008. Nicole Mounier
 Les OGM 5 décembre 2008 Nicole Mounier Université Claude Bernard Lyon 1 CGMC, bâtiment Gregor Mendel 43, boulevard du 11 Novembre 1918 69622 Villeurbanne Cedex OGM Organismes Génétiquement Modifiés Transfert
Les OGM 5 décembre 2008 Nicole Mounier Université Claude Bernard Lyon 1 CGMC, bâtiment Gregor Mendel 43, boulevard du 11 Novembre 1918 69622 Villeurbanne Cedex OGM Organismes Génétiquement Modifiés Transfert
Compétence 3-1 S EXPRIMER A L ECRIT Fiche professeur
 Compétence 3-1 S EXPRIMER A L ECRIT Fiche professeur Nature de l activité : Réaliser 3 types de productions écrites (réécriture de notes, production d une synthèse de documents, production d une argumentation)
Compétence 3-1 S EXPRIMER A L ECRIT Fiche professeur Nature de l activité : Réaliser 3 types de productions écrites (réécriture de notes, production d une synthèse de documents, production d une argumentation)
Séquence 1. Reproduction conforme de la cellule et réplication de l ADN Variabilité génétique et mutation de l ADN
 Séquence 1 Reproduction conforme de la cellule et réplication de l ADN Variabilité génétique et mutation de l ADN Sommaire 1. Reproduction conforme de la cellule et réplication de l ADN 2. Variabilité
Séquence 1 Reproduction conforme de la cellule et réplication de l ADN Variabilité génétique et mutation de l ADN Sommaire 1. Reproduction conforme de la cellule et réplication de l ADN 2. Variabilité
IMMUNOLOGIE. La spécificité des immunoglobulines et des récepteurs T. Informations scientifiques
 IMMUNOLOGIE La spécificité des immunoglobulines et des récepteurs T Informations scientifiques L infection par le VIH entraîne des réactions immunitaires de l organisme qui se traduisent par la production
IMMUNOLOGIE La spécificité des immunoglobulines et des récepteurs T Informations scientifiques L infection par le VIH entraîne des réactions immunitaires de l organisme qui se traduisent par la production
Bases moléculaires des mutations Marc Jeanpierre
 Bases moléculaires des mutations Marc Jeanpierre Chaque enfant qui naît hérite de 10 à 30 nouvelles mutations ponctuelles. L essentiel des ces mutations sont heureusement des variations neutres de séquence
Bases moléculaires des mutations Marc Jeanpierre Chaque enfant qui naît hérite de 10 à 30 nouvelles mutations ponctuelles. L essentiel des ces mutations sont heureusement des variations neutres de séquence
I. La levure Saccharomyces cerevisiae: mode de vie
 LES LEVURES UE «levures» -5 avril: généralités (MN Simon) -6 avril: analyse génétique (MN Simon) -6 avril: Cycle cellulaire I: la réplication (E. bailly) -7 avril: Cycle cellulaire II: la mitose (E. Bailly)
LES LEVURES UE «levures» -5 avril: généralités (MN Simon) -6 avril: analyse génétique (MN Simon) -6 avril: Cycle cellulaire I: la réplication (E. bailly) -7 avril: Cycle cellulaire II: la mitose (E. Bailly)
LA MITOSE CUEEP - USTL DÉPARTEMENT SCIENCES BAHIJA DELATTRE
 Biologie LA MITOSE CUEEP - USTL DÉPARTEMENT SCIENCES BAHIJA DELATTRE Février 2006 I. L'INTRODUCTION Chaque cellule d'un organisme supérieur provient de la multiplication d'une cellule préexistante (cellule
Biologie LA MITOSE CUEEP - USTL DÉPARTEMENT SCIENCES BAHIJA DELATTRE Février 2006 I. L'INTRODUCTION Chaque cellule d'un organisme supérieur provient de la multiplication d'une cellule préexistante (cellule
Hépatite chronique B Moyens thérapeutiques
 Hépatite chronique B Moyens thérapeutiques Dr Olfa BAHRI Laboratoire de Virologie Clinique Institut Pasteur de Tunis INTRODUCTION Plus de 300. 10 6 porteurs chroniques de VHB dans le monde Hépatite chronique
Hépatite chronique B Moyens thérapeutiques Dr Olfa BAHRI Laboratoire de Virologie Clinique Institut Pasteur de Tunis INTRODUCTION Plus de 300. 10 6 porteurs chroniques de VHB dans le monde Hépatite chronique
Univers Vivant Révision. Notions STE
 Univers Vivant Révision Notions STE Chap. 13) L Écologie 1) a) Qu est-ce que l empreinte écologique? L empreinte écologique correspond à la surface terrestre et aquatique totale nécessaire à un individu,
Univers Vivant Révision Notions STE Chap. 13) L Écologie 1) a) Qu est-ce que l empreinte écologique? L empreinte écologique correspond à la surface terrestre et aquatique totale nécessaire à un individu,
EXERCICES : MECANISMES DE L IMMUNITE : pages 406 407 408 409 410
 EXERCICES : MECANISMES DE L IMMUNITE : pages 406 407 408 409 410 EXERCICE 1 PAGE 406 : EXPERIENCES A INTERPRETER Question : rôles respectifs du thymus et de la moelle osseuse dans la production des lymphocytes.
EXERCICES : MECANISMES DE L IMMUNITE : pages 406 407 408 409 410 EXERCICE 1 PAGE 406 : EXPERIENCES A INTERPRETER Question : rôles respectifs du thymus et de la moelle osseuse dans la production des lymphocytes.
Mise en place d une solution automatique de stockage et de visualisation de données de capture des interactions chromatiniennes à l échelle génomique
 Rapport de stage de deuxième année de DUT Génie Biologique option Bioinformatique Mise en place d une solution automatique de stockage et de visualisation de données de capture des interactions chromatiniennes
Rapport de stage de deuxième année de DUT Génie Biologique option Bioinformatique Mise en place d une solution automatique de stockage et de visualisation de données de capture des interactions chromatiniennes
Génomique Comparative et intégrative
 Génomique Comparative et intégrative Introduction : Le big data : on peut traiter des données massives à présent, l'objectif à présent est d'éviter les transferts de données trop longs. On a tout à portée
Génomique Comparative et intégrative Introduction : Le big data : on peut traiter des données massives à présent, l'objectif à présent est d'éviter les transferts de données trop longs. On a tout à portée
Les outils de génétique moléculaire Les techniques liées aux acides nucléiques
 Les outils de génétique moléculaire Les techniques liées aux acides nucléiques Sommaire Preparation des acides nucléiques Extraction / purification Les enzymes agissant sur les acides nucléiques Les enzymes
Les outils de génétique moléculaire Les techniques liées aux acides nucléiques Sommaire Preparation des acides nucléiques Extraction / purification Les enzymes agissant sur les acides nucléiques Les enzymes
Chapitre II La régulation de la glycémie
 Chapitre II La régulation de la glycémie Glycémie : concentration de glucose dans le sang valeur proche de 1g/L Hypoglycémie : perte de connaissance, troubles de la vue, voire coma. Hyperglycémie chronique
Chapitre II La régulation de la glycémie Glycémie : concentration de glucose dans le sang valeur proche de 1g/L Hypoglycémie : perte de connaissance, troubles de la vue, voire coma. Hyperglycémie chronique
Partie Observer : Ondes et matière CHAP 04-ACT/DOC Analyse spectrale : Spectroscopies IR et RMN
 Partie Observer : Ondes et matière CHAP 04-ACT/DOC Analyse spectrale : Spectroscopies IR et RMN Objectifs : Exploiter un spectre infrarouge pour déterminer des groupes caractéristiques Relier un spectre
Partie Observer : Ondes et matière CHAP 04-ACT/DOC Analyse spectrale : Spectroscopies IR et RMN Objectifs : Exploiter un spectre infrarouge pour déterminer des groupes caractéristiques Relier un spectre
Annales de Biologie Cellulaire QCM (niveau SVT 1 er année)
") Annales de Biologie Cellulaire QCM (niveau SVT 1 er année) Equipe pédagogique Université Bordeaux-1 Didier Morin, Michel Moenner, Sophie North, Gérard Tramu et IJsbrand Kramer Contact : [email protected]
Annales de Biologie Cellulaire QCM (niveau SVT 1 er année) Equipe pédagogique Université Bordeaux-1 Didier Morin, Michel Moenner, Sophie North, Gérard Tramu et IJsbrand Kramer Contact : [email protected]
CATALOGUE DES PRESTATIONS DE LA
 1/23 La plate-forme Biopuces et Séquençage de Strasbourg est équipée des technologies Affymetrix et Agilent pour l étude du transcriptome et du génome sur puces à ADN. SOMMAIRE ANALYSE TRANSCRIPTIONNELLE...
1/23 La plate-forme Biopuces et Séquençage de Strasbourg est équipée des technologies Affymetrix et Agilent pour l étude du transcriptome et du génome sur puces à ADN. SOMMAIRE ANALYSE TRANSCRIPTIONNELLE...
------- SESSION 2013 ÉPREUVE À OPTION. (durée : 4 heures coefficient : 6 note éliminatoire 4 sur 20) CHIMIE
 CHIMIE") CNCURS SUR ÉPREUVES UVERT AUX CANDIDATS TITULAIRES D UN DIPLÔME U TITRE CNFÉRANT LE GRADE DE MASTER U D'UN DIPLÔME U TITRE HMLGUÉ U ENREGISTRÉ AU RÉPERTIRE NATINAL DES CERTIFICATINS PRFESSINNELLES AU NIVEAU
CNCURS SUR ÉPREUVES UVERT AUX CANDIDATS TITULAIRES D UN DIPLÔME U TITRE CNFÉRANT LE GRADE DE MASTER U D'UN DIPLÔME U TITRE HMLGUÉ U ENREGISTRÉ AU RÉPERTIRE NATINAL DES CERTIFICATINS PRFESSINNELLES AU NIVEAU
De la physico-chimie à la radiobiologie: nouveaux acquis (I)
") De la physico-chimie à la radiobiologie: nouveaux acquis (I) Collaboration: - Laboratoire de Radiotoxicologie et Oncologie (L. Sabatier) CEA, DSV - Laboratoire de Génotoxicité et Modulation de l Expression
De la physico-chimie à la radiobiologie: nouveaux acquis (I) Collaboration: - Laboratoire de Radiotoxicologie et Oncologie (L. Sabatier) CEA, DSV - Laboratoire de Génotoxicité et Modulation de l Expression
Lycée Galilée Gennevilliers. chap. 6. JALLU Laurent. I. Introduction... 2 La source d énergie nucléaire... 2
 Lycée Galilée Gennevilliers L'énergie nucléaire : fusion et fission chap. 6 JALLU Laurent I. Introduction... 2 La source d énergie nucléaire... 2 II. Équivalence masse-énergie... 3 Bilan de masse de la
Lycée Galilée Gennevilliers L'énergie nucléaire : fusion et fission chap. 6 JALLU Laurent I. Introduction... 2 La source d énergie nucléaire... 2 II. Équivalence masse-énergie... 3 Bilan de masse de la
Conférence technique internationale de la FAO
 Décembre 2009 ABDC-10/7.2 F Conférence technique internationale de la FAO Biotechnologies agricoles dans les pays en développement: choix et perspectives pour les cultures, les forêts, l élevage, les pêches
Décembre 2009 ABDC-10/7.2 F Conférence technique internationale de la FAO Biotechnologies agricoles dans les pays en développement: choix et perspectives pour les cultures, les forêts, l élevage, les pêches
Tableau 1. Liste (non exhaustive) des protéines se localisant dans les P-Bodies
 des protéines se localisant dans les P-Bodies") NOM FONCTION EFFET DE L ABSENCE OU DE REFERENCES LA SUREXPRESSION SUR LES P- BODIES XRN1 exonucléase 5 3 Absence : augmente la taille et le Bashkirov et al. 1997 Shet et Parker 2003 Cougot nombre des P-bodies
NOM FONCTION EFFET DE L ABSENCE OU DE REFERENCES LA SUREXPRESSION SUR LES P- BODIES XRN1 exonucléase 5 3 Absence : augmente la taille et le Bashkirov et al. 1997 Shet et Parker 2003 Cougot nombre des P-bodies
2 C est quoi la chimie?
 PARTIE 1 AVANT LA CHIMIE VERTE... 2 C est quoi la chimie? L inconnu étant source d angoisse, nous allons essayer de définir les grands domaines de la chimie pour mieux la connaître, l appréhender et donc
PARTIE 1 AVANT LA CHIMIE VERTE... 2 C est quoi la chimie? L inconnu étant source d angoisse, nous allons essayer de définir les grands domaines de la chimie pour mieux la connaître, l appréhender et donc
Biomarqueurs en Cancérologie
 Biomarqueurs en Cancérologie Définition, détermination, usage Biomarqueurs et Cancer: définition Anomalie(s) quantitative(s) ou qualitative(s) Indicative(s) ou caractéristique(s) d un cancer ou de certaines
Biomarqueurs en Cancérologie Définition, détermination, usage Biomarqueurs et Cancer: définition Anomalie(s) quantitative(s) ou qualitative(s) Indicative(s) ou caractéristique(s) d un cancer ou de certaines
3: Clonage d un gène dans un plasmide
 3: Clonage d un gène dans un plasmide Le clonage moléculaire est une des bases du génie génétique. Il consiste à insérer un fragment d'adn (dénommé insert) dans un vecteur approprié comme un plasmide par
3: Clonage d un gène dans un plasmide Le clonage moléculaire est une des bases du génie génétique. Il consiste à insérer un fragment d'adn (dénommé insert) dans un vecteur approprié comme un plasmide par
UE : GENE 2208. Responsable : Enseignant : ECUE 1. Enseignant : ECUE 2. Dr COULIBALY Foungotin Hamidou
 UE : GENE 2208 Responsable : Enseignant : ECUE 1 Enseignant : ECUE 2 Dr COULIBALY Foungotin Hamidou Spécialités : Génétique Humaine Biologie de la Procréation Cytogénétique Spermiologie Essais Cliniques
UE : GENE 2208 Responsable : Enseignant : ECUE 1 Enseignant : ECUE 2 Dr COULIBALY Foungotin Hamidou Spécialités : Génétique Humaine Biologie de la Procréation Cytogénétique Spermiologie Essais Cliniques
Les débuts de la génétique
 HPITRE 9 DES DÉBTS DE L ÉNÉTIQE X ENJEX TELS DES BIOTEHNOLOIES 1 Les débuts de la génétique est avec les travaux de regor Mendel vers la fin du XIX e siècle que furent posées les bases de la génétique.
HPITRE 9 DES DÉBTS DE L ÉNÉTIQE X ENJEX TELS DES BIOTEHNOLOIES 1 Les débuts de la génétique est avec les travaux de regor Mendel vers la fin du XIX e siècle que furent posées les bases de la génétique.
Université d Evry-Val d Essonne Ecole Doctorale des Génomes Aux Organismes. Thèse
 Université d Evry-Val d Essonne Ecole Doctorale des Génomes Aux Organismes Thèse Présentée pour obtenir le grade de Docteur en sciences de l université d Evry-Val d Essonne Spécialité Bioinformatique par
Université d Evry-Val d Essonne Ecole Doctorale des Génomes Aux Organismes Thèse Présentée pour obtenir le grade de Docteur en sciences de l université d Evry-Val d Essonne Spécialité Bioinformatique par
La physique nucléaire et ses applications
 La physique nucléaire et ses applications I. Rappels et compléments sur les noyaux. Sa constitution La représentation symbolique d'un noyau est, dans laquelle : o X est le symbole du noyau et par extension
La physique nucléaire et ses applications I. Rappels et compléments sur les noyaux. Sa constitution La représentation symbolique d'un noyau est, dans laquelle : o X est le symbole du noyau et par extension
Transformations nucléaires
 Transformations nucléaires Stabilité et instabilité des noyaux : Le noyau d un atome associé à un élément est représenté par le symbole A : nombre de masse = nombre de nucléons (protons + neutrons) Z :
Transformations nucléaires Stabilité et instabilité des noyaux : Le noyau d un atome associé à un élément est représenté par le symbole A : nombre de masse = nombre de nucléons (protons + neutrons) Z :
La gestion de données dans le cadre d une application de recherche d alignement de séquence : BLAST.
 La gestion de données dans le cadre d une application de recherche d alignement de séquence : BLAST. Gaël Le Mahec - p. 1/12 L algorithme BLAST. Basic Local Alignment Search Tool est un algorithme de recherche
La gestion de données dans le cadre d une application de recherche d alignement de séquence : BLAST. Gaël Le Mahec - p. 1/12 L algorithme BLAST. Basic Local Alignment Search Tool est un algorithme de recherche
MAB Solut. vos projets. MABLife Génopole Campus 1 5 rue Henri Desbruères 91030 Evry Cedex. www.mabsolut.com. intervient à chaque étape de
 Mabsolut-DEF-HI:Mise en page 1 17/11/11 17:45 Page1 le département prestataire de services de MABLife de la conception à la validation MAB Solut intervient à chaque étape de vos projets Création d anticorps
Mabsolut-DEF-HI:Mise en page 1 17/11/11 17:45 Page1 le département prestataire de services de MABLife de la conception à la validation MAB Solut intervient à chaque étape de vos projets Création d anticorps
COUSIN Fabien KERGOURLAY Gilles. 19 octobre 2007. de l hôte par les. Master 2 MFA Responsable : UE Incidence des paramètres environnementaux
 COUSIN Fabien KERGOURLAY Gilles 19 octobre 2007 Inhibition des défenses de l hôte par les bactéries pathogènes Master 2 MFA Responsable : UE Incidence des paramètres environnementaux Gwennola ERMEL I Détection
COUSIN Fabien KERGOURLAY Gilles 19 octobre 2007 Inhibition des défenses de l hôte par les bactéries pathogènes Master 2 MFA Responsable : UE Incidence des paramètres environnementaux Gwennola ERMEL I Détection
4 : MÉTHODES D ANALYSE UTILISÉES EN ÉCOLOGIE MICROBIENNE
 4 : MÉTHODES D ANALYSE UTILISÉES EN ÉCOLOGIE MICROBIENNE L écologie microbienne (ou étude des micro-organismes de l environnement) étudie : les relations entre les différentes populations de micro-organismes
4 : MÉTHODES D ANALYSE UTILISÉES EN ÉCOLOGIE MICROBIENNE L écologie microbienne (ou étude des micro-organismes de l environnement) étudie : les relations entre les différentes populations de micro-organismes
Comprendre l Univers grâce aux messages de la lumière
 Seconde / P4 Comprendre l Univers grâce aux messages de la lumière 1/ EXPLORATION DE L UNIVERS Dans notre environnement quotidien, les dimensions, les distances sont à l échelle humaine : quelques mètres,
Seconde / P4 Comprendre l Univers grâce aux messages de la lumière 1/ EXPLORATION DE L UNIVERS Dans notre environnement quotidien, les dimensions, les distances sont à l échelle humaine : quelques mètres,
Chapitre 6. Réactions nucléaires. 6.1 Généralités. 6.1.1 Définitions. 6.1.2 Lois de conservation
 Chapitre 6 Réactions nucléaires 6.1 Généralités 6.1.1 Définitions Un atome est constitué d électrons et d un noyau, lui-même constitué de nucléons (protons et neutrons). Le nombre de masse, noté, est le
Chapitre 6 Réactions nucléaires 6.1 Généralités 6.1.1 Définitions Un atome est constitué d électrons et d un noyau, lui-même constitué de nucléons (protons et neutrons). Le nombre de masse, noté, est le
MABioVis. Bio-informatique et la
 MABioVis Modèles et Algorithmes pour la Bio-informatique et la Visualisation Visite ENS Cachan 5 janvier 2011 MABioVis G GUY MELANÇON (PR UFR Maths Info / EPI GRAVITE) (là, maintenant) - MABioVis DAVID
MABioVis Modèles et Algorithmes pour la Bio-informatique et la Visualisation Visite ENS Cachan 5 janvier 2011 MABioVis G GUY MELANÇON (PR UFR Maths Info / EPI GRAVITE) (là, maintenant) - MABioVis DAVID
Chapitre n 6 MASSE ET ÉNERGIE DES NOYAUX
 Chapitre n 6 MASSE ET ÉNERGIE DES NOYAUX T ale S Introduction : Une réaction nucléaire est Une réaction nucléaire provoquée est L'unité de masse atomique est une unité permettant de manipuler aisément
Chapitre n 6 MASSE ET ÉNERGIE DES NOYAUX T ale S Introduction : Une réaction nucléaire est Une réaction nucléaire provoquée est L'unité de masse atomique est une unité permettant de manipuler aisément
Chapitre 02. La lumière des étoiles. Exercices :
 Chapitre 02 La lumière des étoiles. I- Lumière monochromatique et lumière polychromatique. )- Expérience de Newton (642 727). 2)- Expérience avec la lumière émise par un Laser. 3)- Radiation et longueur
Chapitre 02 La lumière des étoiles. I- Lumière monochromatique et lumière polychromatique. )- Expérience de Newton (642 727). 2)- Expérience avec la lumière émise par un Laser. 3)- Radiation et longueur
INTRODUCTION À L'ENZYMOLOGIE
 INTRODUCTION À L'ENZYMOLOGIE Les enzymes sont des macromolécules spécialisées qui - catalysent les réactions biologiques - transforment différentes formes d'énergie. Les enzymes diffèrent des catalyseurs
INTRODUCTION À L'ENZYMOLOGIE Les enzymes sont des macromolécules spécialisées qui - catalysent les réactions biologiques - transforment différentes formes d'énergie. Les enzymes diffèrent des catalyseurs
Réseaux grande distance
 Chapitre 5 Réseaux grande distance 5.1 Définition Les réseaux à grande distance (WAN) reposent sur une infrastructure très étendue, nécessitant des investissements très lourds. Contrairement aux réseaux
Chapitre 5 Réseaux grande distance 5.1 Définition Les réseaux à grande distance (WAN) reposent sur une infrastructure très étendue, nécessitant des investissements très lourds. Contrairement aux réseaux
ACIDES BASES. Chap.5 SPIESS
 ACIDES BASES «Je ne crois pas que l on me conteste que l acide n ait des pointes Il ne faut que le goûter pour tomber dans ce sentiment car il fait des picotements sur la langue.» Notion d activité et
ACIDES BASES «Je ne crois pas que l on me conteste que l acide n ait des pointes Il ne faut que le goûter pour tomber dans ce sentiment car il fait des picotements sur la langue.» Notion d activité et
Organisme de recherche et d information sur la logistique et le transport LES TECHNIQUES DE SUIVI DES ARTICLES ET DES STOCKS
 LES TECHNIQUES DE SUIVI DES ARTICLES ET DES STOCKS La gestion et la maîtrise des flux entrants et sortants du magasin nécessite la mise en œuvre d un dispositif à trois composantes : la classification
LES TECHNIQUES DE SUIVI DES ARTICLES ET DES STOCKS La gestion et la maîtrise des flux entrants et sortants du magasin nécessite la mise en œuvre d un dispositif à trois composantes : la classification
a. Fusion et énergie de liaison des noyaux b. La barrière Coulombienne c. Effet tunnel & pic de Gamov
 V. Les réactions r thermonucléaires 1. Principes a. Fusion et énergie de liaison des noyaux b. La barrière Coulombienne c. Effet tunnel & pic de Gamov 2. Taux de réactions r thermonucléaires a. Les sections
V. Les réactions r thermonucléaires 1. Principes a. Fusion et énergie de liaison des noyaux b. La barrière Coulombienne c. Effet tunnel & pic de Gamov 2. Taux de réactions r thermonucléaires a. Les sections
ÉCOLES NORMALES SUPÉRIEURES ÉCOLE NATIONALE DES PONTS ET CHAUSSÉES CONCOURS D ADMISSION SESSION 2013 FILIÈRE BCPST COMPOSITION DE BIOLOGIE
 ÉCOLES NORMALES SUPÉRIEURES ÉCOLE NATIONALE DES PONTS ET CHAUSSÉES CONCOURS D ADMISSION SESSION 2013 FILIÈRE BCPST COMPOSITION DE BIOLOGIE Épreuve commune aux ENS de Cachan, Lyon, Paris et de l ENPC Durée
ÉCOLES NORMALES SUPÉRIEURES ÉCOLE NATIONALE DES PONTS ET CHAUSSÉES CONCOURS D ADMISSION SESSION 2013 FILIÈRE BCPST COMPOSITION DE BIOLOGIE Épreuve commune aux ENS de Cachan, Lyon, Paris et de l ENPC Durée
MYRIAD. l ADN isolé n est à présent plus brevetable!
 MYRIAD La Cour Suprême des Etats-Unis revient sur plus de 30 ans de pratique : l ADN isolé n est à présent plus brevetable! Mauvaise passe pour les inventions en biotechnologies sur le territoire américain.
MYRIAD La Cour Suprême des Etats-Unis revient sur plus de 30 ans de pratique : l ADN isolé n est à présent plus brevetable! Mauvaise passe pour les inventions en biotechnologies sur le territoire américain.
Energie Nucléaire. Principes, Applications & Enjeux. 6 ème - 2014/2015
 Energie Nucléaire Principes, Applications & Enjeux 6 ème - 2014/2015 Quelques constats Le belge consomme 3 fois plus d énergie que le terrien moyen; (0,56% de la consommation mondiale pour 0,17% de la
Energie Nucléaire Principes, Applications & Enjeux 6 ème - 2014/2015 Quelques constats Le belge consomme 3 fois plus d énergie que le terrien moyen; (0,56% de la consommation mondiale pour 0,17% de la
Insulinothérapie et diabète de type 1
 Insulinothérapie et diabète de type 1 Introduction: la molécule d insuline L instauration de l insulinothérapie Dispositif d administration de l insuline Les propriétés de l insuline Insuline et schémas
Insulinothérapie et diabète de type 1 Introduction: la molécule d insuline L instauration de l insulinothérapie Dispositif d administration de l insuline Les propriétés de l insuline Insuline et schémas
Production d une protéine recombinante
 99 Production d une protéine recombinante Lic. B. PIRSON Lic. J-M. SERONT ISICHt - Mons Production de la protéine recombinante GFP (Green Fluorescent Protein d Aequoria victoria) par une bactérie ( E.
99 Production d une protéine recombinante Lic. B. PIRSON Lic. J-M. SERONT ISICHt - Mons Production de la protéine recombinante GFP (Green Fluorescent Protein d Aequoria victoria) par une bactérie ( E.
Université de Montréal. Développement d outils pour l analyse de données de ChIP-seq et l identification des facteurs de transcription
 Université de Montréal Développement d outils pour l analyse de données de ChIP-seq et l identification des facteurs de transcription par Eloi Mercier Département de bioinformatique Faculté de médecine
Université de Montréal Développement d outils pour l analyse de données de ChIP-seq et l identification des facteurs de transcription par Eloi Mercier Département de bioinformatique Faculté de médecine
Plateforme Transgenèse/Zootechnie/Exploration Fonctionnelle IBiSA. «Anexplo» Service Transgenèse. Catalogue des prestations
 Plateforme Transgenèse/Zootechnie/Exploration Fonctionnelle IBiSA «Anexplo» Service Transgenèse Catalogue des prestations 04/01/12 - Page 1 sur 8 Présentation du service de Transgenèse Le service de Transgenèse
Plateforme Transgenèse/Zootechnie/Exploration Fonctionnelle IBiSA «Anexplo» Service Transgenèse Catalogue des prestations 04/01/12 - Page 1 sur 8 Présentation du service de Transgenèse Le service de Transgenèse
Cet article s attache tout d abord
 Méthodes internationales pour comparer l éducation et l équité Comparaison entre pays des coûts de l éducation : des sources de financement aux dépenses Luc Brière Marguerite Rudolf Bureau du compte de
Méthodes internationales pour comparer l éducation et l équité Comparaison entre pays des coûts de l éducation : des sources de financement aux dépenses Luc Brière Marguerite Rudolf Bureau du compte de
Eléments de spécification des systèmes temps réel Pierre-Yves Duval (cppm)
") Eléments de spécification des systèmes temps réel Pierre-Yves Duval (cppm) Ecole d informatique temps réel - La Londes les Maures 7-11 Octobre 2002 - Evénements et architectures - Spécifications de performances
Eléments de spécification des systèmes temps réel Pierre-Yves Duval (cppm) Ecole d informatique temps réel - La Londes les Maures 7-11 Octobre 2002 - Evénements et architectures - Spécifications de performances
Nombres, mesures et incertitudes en sciences physiques et chimiques. Groupe des Sciences physiques et chimiques de l IGEN
 Nombres, mesures et incertitudes en sciences physiques et chimiques. Groupe des Sciences physiques et chimiques de l IGEN Table des matières. Introduction....3 Mesures et incertitudes en sciences physiques
Nombres, mesures et incertitudes en sciences physiques et chimiques. Groupe des Sciences physiques et chimiques de l IGEN Table des matières. Introduction....3 Mesures et incertitudes en sciences physiques
Formavie 2010. 2 Différentes versions du format PDB...3. 3 Les champs dans les fichiers PDB...4. 4 Le champ «ATOM»...5. 6 Limites du format PDB...
 Formavie 2010 Les fichiers PDB Les fichiers PDB contiennent les informations qui vont permettre à des logiciels de visualisation moléculaire (ex : RasTop ou Jmol) d afficher les molécules. Un fichier au
Formavie 2010 Les fichiers PDB Les fichiers PDB contiennent les informations qui vont permettre à des logiciels de visualisation moléculaire (ex : RasTop ou Jmol) d afficher les molécules. Un fichier au
L universalité et la variabilité de l ADN
 L universalité et la variabilité de l DN Unité 4 3 μm hromosomes observés en microscopie électronique à balayage. Les chromosomes, présents dans le noyau, sont constitués d acide désoxyribonucléique (DN).
L universalité et la variabilité de l DN Unité 4 3 μm hromosomes observés en microscopie électronique à balayage. Les chromosomes, présents dans le noyau, sont constitués d acide désoxyribonucléique (DN).
Big data et sciences du Vivant L'exemple du séquençage haut débit
 Big data et sciences du Vivant L'exemple du séquençage haut débit C. Gaspin, C. Hoede, C. Klopp, D. Laborie, J. Mariette, C. Noirot, MS. Trotard [email protected] INRA - MIAT - Plate-forme
Big data et sciences du Vivant L'exemple du séquençage haut débit C. Gaspin, C. Hoede, C. Klopp, D. Laborie, J. Mariette, C. Noirot, MS. Trotard [email protected] INRA - MIAT - Plate-forme
Compléments - Chapitre 5 Spectroscopie
 ompléments - hapitre 5 Spectroscopie Spectroscopie par résonance magnétique nucléaire (RMN 13 ) Tandis que la spectroscopie RMN 1 H fournit des données sur la disposition des atomes d'hydrogène dans une
ompléments - hapitre 5 Spectroscopie Spectroscopie par résonance magnétique nucléaire (RMN 13 ) Tandis que la spectroscopie RMN 1 H fournit des données sur la disposition des atomes d'hydrogène dans une
1.2 Coordinence. Notion de liaison de coordinence : Cas de NH 3. et NH 4+ , 3 liaisons covalentes + 1 liaison de coordinence.
 Règle de l octet : tendance qu on les atomes à s entourer de 8 électrons dans l édifice moléculaire. Ce n est pas une règle générale. Composés respectant la règle de l octet Composés ne respectant pas
Règle de l octet : tendance qu on les atomes à s entourer de 8 électrons dans l édifice moléculaire. Ce n est pas une règle générale. Composés respectant la règle de l octet Composés ne respectant pas
Détection des duplications en tandem au niveau nucléique à l'aide de la théorie des flots
 Université Toulouse 3 Paul Sabatier(UT3 Paul Sabatier) Informatique Spécialité Bioinformatique Eric AUDEMARD lundi 28 novembre 2011 Détection des duplications en tandem au niveau nucléique à l'aide de
Université Toulouse 3 Paul Sabatier(UT3 Paul Sabatier) Informatique Spécialité Bioinformatique Eric AUDEMARD lundi 28 novembre 2011 Détection des duplications en tandem au niveau nucléique à l'aide de
Energie nucléaire. Quelques éléments de physique
 Energie nucléaire Quelques éléments de physique Comment produire 1 GW électrique Nucléaire (rendement 33%) Thermique (38%) Hydraulique (85%) Solaire (10%) Vent : 27t d uranium par an : 170 t de fuel par
Energie nucléaire Quelques éléments de physique Comment produire 1 GW électrique Nucléaire (rendement 33%) Thermique (38%) Hydraulique (85%) Solaire (10%) Vent : 27t d uranium par an : 170 t de fuel par
Luca : à la recherche du plus proche ancêtre commun universel Patrick Forterre, Simonetta Gribaldo, Céline Brochier
 MEDECINE/SCIENCES 2005 ; 21 :???-??? > L application des méthodes de la biologie moléculaire à l étude des premières étapes de l évolution a montré que le monde vivant pouvait être divisé en trois domaines
MEDECINE/SCIENCES 2005 ; 21 :???-??? > L application des méthodes de la biologie moléculaire à l étude des premières étapes de l évolution a montré que le monde vivant pouvait être divisé en trois domaines
POLY-PREPAS Centre de Préparation aux Concours Paramédicaux. - Section Orthoptiste / stage i-prépa intensif -
 POLY-PREPAS Centre de Préparation aux Concours Paramédicaux - Section Orthoptiste / stage i-prépa intensif - 1 Suite énoncé des exos du Chapitre 14 : Noyaux-masse-énergie I. Fission nucléaire induite (provoquée)
POLY-PREPAS Centre de Préparation aux Concours Paramédicaux - Section Orthoptiste / stage i-prépa intensif - 1 Suite énoncé des exos du Chapitre 14 : Noyaux-masse-énergie I. Fission nucléaire induite (provoquée)
Chapitre 10 : Radioactivité et réactions nucléaires (chapitre 11 du livre)
") Chapitre 10 : Radioactivité et réactions nucléaires (chapitre 11 du livre) 1. A la découverte de la radioactivité. Un noyau père radioactif est un noyau INSTABLE. Il se transforme en un noyau fils STABLE
Chapitre 10 : Radioactivité et réactions nucléaires (chapitre 11 du livre) 1. A la découverte de la radioactivité. Un noyau père radioactif est un noyau INSTABLE. Il se transforme en un noyau fils STABLE
Équivalence masse-énergie
 CHPITRE 5 NOYUX, MSSE ET ÉNERGIE Équivalence masse-énergie. Équivalence masse-énergie Einstein a montré que la masse constitue une forme d énergie appelée énergie de masse. La relation entre la masse (en
CHPITRE 5 NOYUX, MSSE ET ÉNERGIE Équivalence masse-énergie. Équivalence masse-énergie Einstein a montré que la masse constitue une forme d énergie appelée énergie de masse. La relation entre la masse (en
Chapitre V : La gestion de la mémoire. Hiérarchie de mémoires Objectifs Méthodes d'allocation Simulation de mémoire virtuelle Le mapping
 Chapitre V : La gestion de la mémoire Hiérarchie de mémoires Objectifs Méthodes d'allocation Simulation de mémoire virtuelle Le mapping Introduction Plusieurs dizaines de processus doivent se partager
Chapitre V : La gestion de la mémoire Hiérarchie de mémoires Objectifs Méthodes d'allocation Simulation de mémoire virtuelle Le mapping Introduction Plusieurs dizaines de processus doivent se partager
Structure quantique cohérente et incohérente de l eau liquide
 Structure quantique cohérente et incohérente de l eau liquide Prof. Marc HENRY Chimie Moléculaire du Solide Institut Le Bel, 4, Rue Blaise Pascal 67070 Strasbourg Cedex, France Tél: 03.68.85.15.00 e-mail:
Structure quantique cohérente et incohérente de l eau liquide Prof. Marc HENRY Chimie Moléculaire du Solide Institut Le Bel, 4, Rue Blaise Pascal 67070 Strasbourg Cedex, France Tél: 03.68.85.15.00 e-mail:
Biologie Appliquée. Dosages Immunologiques TD9 Mai 2015. Stéphanie Sigaut INSERM U1141 [email protected]
 Biologie Appliquée Dosages Immunologiques TD9 Mai 2015 Stéphanie Sigaut INSERM U1141 [email protected] 1 ELISA 2 3 4 [Ac] 5 6 7 8 9 Correction : Faire la moyenne D0-1 et D0-2 pour toute les valeurs
Biologie Appliquée Dosages Immunologiques TD9 Mai 2015 Stéphanie Sigaut INSERM U1141 [email protected] 1 ELISA 2 3 4 [Ac] 5 6 7 8 9 Correction : Faire la moyenne D0-1 et D0-2 pour toute les valeurs
L ÉNERGIE C EST QUOI?
 L ÉNERGIE C EST QUOI? L énergie c est la vie! Pourquoi à chaque fois qu on fait quelque chose on dit qu on a besoin d énergie? Parce que l énergie est à l origine de tout! Rien ne peut se faire sans elle.
L ÉNERGIE C EST QUOI? L énergie c est la vie! Pourquoi à chaque fois qu on fait quelque chose on dit qu on a besoin d énergie? Parce que l énergie est à l origine de tout! Rien ne peut se faire sans elle.
Suivi d une réaction lente par chromatographie
 TS Activité Chapitre 8 Cinétique chimique Suivi d une réaction lente par chromatographie Objectifs : Analyser un protocole expérimental de synthèse chimique Analyser un chromatogramme pour mettre en évidence
TS Activité Chapitre 8 Cinétique chimique Suivi d une réaction lente par chromatographie Objectifs : Analyser un protocole expérimental de synthèse chimique Analyser un chromatogramme pour mettre en évidence
Séquence 4. La nature du vivant. Sommaire. 1. L unité structurale et chimique du vivant. 2. L ADN, support de l information génétique
 Séquence 4 La nature du vivant Sommaire 1. L unité structurale et chimique du vivant 2. L ADN, support de l information génétique 3. Synthèse de la séquence 4. Exercices Devoir autocorrectif n 2 Séquence
Séquence 4 La nature du vivant Sommaire 1. L unité structurale et chimique du vivant 2. L ADN, support de l information génétique 3. Synthèse de la séquence 4. Exercices Devoir autocorrectif n 2 Séquence
Séquence 6. Mais ces espèces pour autant ne sont pas identiques et parfois d ailleurs ne se ressemblent pas vraiment.
 Sommaire Séquence 6 Nous avons vu dans les séances précédentes qu au cours des temps géologiques des espèces différentes se sont succédé, leur apparition et leur disparition étant le résultat de modifications
Sommaire Séquence 6 Nous avons vu dans les séances précédentes qu au cours des temps géologiques des espèces différentes se sont succédé, leur apparition et leur disparition étant le résultat de modifications
ATELIER SANTE PREVENTION N 2 : L ALIMENTATION
 ATELIER SANTE PREVENTION N 2 : L ALIMENTATION Mardi 24 janvier 2012 au Centre de Formation Multimétiers de REIGNAC L objectif de cet atelier sur la santé est de guider chacun vers une alimentation plus
ATELIER SANTE PREVENTION N 2 : L ALIMENTATION Mardi 24 janvier 2012 au Centre de Formation Multimétiers de REIGNAC L objectif de cet atelier sur la santé est de guider chacun vers une alimentation plus
8/10/10. Les réactions nucléaires
 Les réactions nucléaires En 1900, à Montréal, Rutherford observa un effet curieux, lors de mesures de l'intensité du rayonnement d'une source de thorium [...]. L'intensité n'était pas la même selon que
Les réactions nucléaires En 1900, à Montréal, Rutherford observa un effet curieux, lors de mesures de l'intensité du rayonnement d'une source de thorium [...]. L'intensité n'était pas la même selon que
grande simple microscope microscope inventé années 1825. biologie = cellule) et (logos de plus en Anglais. Utilise un La microscopie, 1665,
 et (logos de plus en Anglais. Utilise un La microscopie, 1665,") Cours de Biologie Cellulaire Présentés par Mr CHELLI A. FSNV 2012/ /2013 CHAPITRE I : INTRODUCTION A LA BIOLOGIE CELLULAIRE A- Introduction et définitionn de la biologie cellulaire : Il était difficile
Cours de Biologie Cellulaire Présentés par Mr CHELLI A. FSNV 2012/ /2013 CHAPITRE I : INTRODUCTION A LA BIOLOGIE CELLULAIRE A- Introduction et définitionn de la biologie cellulaire : Il était difficile
ATELIER IMAGEJ. Différentes applications vous sont proposées pour apprendre à utiliser quelques fonctions d ImageJ :
 Différentes applications vous sont proposées pour apprendre à utiliser quelques fonctions d ImageJ : 1. ANALYSE QUANTITATIVE D UN GEL D ELECTROPHORESE... 2 2. NUMERATION DE COLONIES BACTERIENNES SUR UNE
Différentes applications vous sont proposées pour apprendre à utiliser quelques fonctions d ImageJ : 1. ANALYSE QUANTITATIVE D UN GEL D ELECTROPHORESE... 2 2. NUMERATION DE COLONIES BACTERIENNES SUR UNE
Introduction à la Génomique Fonctionnelle
 Introduction à la Génomique Fonctionnelle Cours aux étudiants de BSc Biologie 3ème année Philippe Reymond, MER PLAN DU COURS - Séquençage des génomes - Fabrication de DNA microarrays - Autres méthodes
Introduction à la Génomique Fonctionnelle Cours aux étudiants de BSc Biologie 3ème année Philippe Reymond, MER PLAN DU COURS - Séquençage des génomes - Fabrication de DNA microarrays - Autres méthodes
Vue d ensemble de la vie microbienne
 Vue d ensemble de la vie microbienne C HAPITRE D EUX I Structure cellulaire et évolution 22 2.1 Les structures cellulaires et virales 22 2.2 L organisation de l ADN dans les cellules microbiennes 24 2.3
Vue d ensemble de la vie microbienne C HAPITRE D EUX I Structure cellulaire et évolution 22 2.1 Les structures cellulaires et virales 22 2.2 L organisation de l ADN dans les cellules microbiennes 24 2.3
Chapitre 2 - Complexité des relations entre génotype et phénotype
 Chapitre 2 - Complexité des relations entre génotype et phénotype Chaque chromosome est en double exemplaire Donc chaque gène (situé sur son locus) est en double exemplaires : et peut être sous différente
Chapitre 2 - Complexité des relations entre génotype et phénotype Chaque chromosome est en double exemplaire Donc chaque gène (situé sur son locus) est en double exemplaires : et peut être sous différente
ANALYSE SPECTRALE. monochromateur
 ht ANALYSE SPECTRALE Une espèce chimique est susceptible d interagir avec un rayonnement électromagnétique. L étude de l intensité du rayonnement (absorbé ou réémis) en fonction des longueurs d ode s appelle
ht ANALYSE SPECTRALE Une espèce chimique est susceptible d interagir avec un rayonnement électromagnétique. L étude de l intensité du rayonnement (absorbé ou réémis) en fonction des longueurs d ode s appelle
Utilisation des substrats énergétiques
 Utilisation des substrats énergétiques Collège des Enseignants de Nutrition Date de création du document 2010-2011 Table des matières I Les organes et les substrats... 3 I.1 Les substrats énergétiques...
Utilisation des substrats énergétiques Collège des Enseignants de Nutrition Date de création du document 2010-2011 Table des matières I Les organes et les substrats... 3 I.1 Les substrats énergétiques...
Chapitre 11: Réactions nucléaires, radioactivité et fission
 1re B et C 11 Réactions nucléaires, radioactivité et fission 129 Chapitre 11: Réactions nucléaires, radioactivité et fission 1. Définitions a) Nucléides (= noyaux atomiques) Les nucléides renferment les
1re B et C 11 Réactions nucléaires, radioactivité et fission 129 Chapitre 11: Réactions nucléaires, radioactivité et fission 1. Définitions a) Nucléides (= noyaux atomiques) Les nucléides renferment les
Physique Chimie. Utiliser les langages scientifiques à l écrit et à l oral pour interpréter les formules chimiques
 C est Niveau la représentation 4 ème 2. Document du professeur 1/6 Physique Chimie LES ATOMES POUR COMPRENDRE LA TRANSFORMATION CHIMIQUE Programme Cette séance expérimentale illustre la partie de programme
C est Niveau la représentation 4 ème 2. Document du professeur 1/6 Physique Chimie LES ATOMES POUR COMPRENDRE LA TRANSFORMATION CHIMIQUE Programme Cette séance expérimentale illustre la partie de programme
3 Charges électriques
 3 Charges électriques 3.1 Electrisation par frottement Expérience : Frottons un bâton d ébonite avec un morceau de peau de chat. Approchonsle de petits bouts de papier. On observe que les bouts de papier
3 Charges électriques 3.1 Electrisation par frottement Expérience : Frottons un bâton d ébonite avec un morceau de peau de chat. Approchonsle de petits bouts de papier. On observe que les bouts de papier
Bulletin officiel n 29 du 19 juillet 2012 Sommaire
 Bulletin officiel n 29 du 9 juillet 22 Sommaire Organisation générale Administration centrale du MN et du MSR Attributions de fonctions : modification arrêté du 5-6-22 (NOR : MNA2252A) Administration centrale
Bulletin officiel n 29 du 9 juillet 22 Sommaire Organisation générale Administration centrale du MN et du MSR Attributions de fonctions : modification arrêté du 5-6-22 (NOR : MNA2252A) Administration centrale
5.5.5 Exemple d un essai immunologique
 5.5.5 Exemple d un essai immunologique Test de grossesse Test en forme de bâtonnet destiné à mettre en évidence l'hormone spécifique de la grossesse, la gonadotrophine chorionique humaine (hcg), une glycoprotéine.
5.5.5 Exemple d un essai immunologique Test de grossesse Test en forme de bâtonnet destiné à mettre en évidence l'hormone spécifique de la grossesse, la gonadotrophine chorionique humaine (hcg), une glycoprotéine.
La reconnaissance moléculaire: la base du design rationnel Modélisation moléculaire: Introduction Hiver 2006
 La reconnaissance moléculaire: la base du design rationnel En 1890 Emil Fisher a proposé le modèle "serrure et clé" pour expliquer la façon de fonctionner des systèmes biologiques. Un substrat rentre et
La reconnaissance moléculaire: la base du design rationnel En 1890 Emil Fisher a proposé le modèle "serrure et clé" pour expliquer la façon de fonctionner des systèmes biologiques. Un substrat rentre et
CHAPITRE 3 LES TASQ COMME PLATEFORME POUR LA CATALYSE PSEUDO-ENZYMATIQUE
 L. Stefan Chapitre 3 CHAPITRE 3 LES TASQ COMME PLATEFORME POUR LA CATALYSE PSEUDO-ENZYMATIQUE Nous avons démontré dans le chapitre précédent que les TASQ hydrosolubles développés au cours de cette thèse
L. Stefan Chapitre 3 CHAPITRE 3 LES TASQ COMME PLATEFORME POUR LA CATALYSE PSEUDO-ENZYMATIQUE Nous avons démontré dans le chapitre précédent que les TASQ hydrosolubles développés au cours de cette thèse