CYCLE CELLULAIRE Phase G Phase S Phase G2 Phase M
|
|
|
- Marie-Claude Papineau
- il y a 10 ans
- Total affichages :
Transcription
1 NOYAU INTERPHASIQUE

2 CYCLE CELLULAIRE C'est l'intervalle de temps s'écoulant entre le moment où une cellule vient de se former (après une mitose) et le moment où elle donne deux cellules filles à la suite d'une nouvelle division. C'est donc une interphase et une mitose: c'est le temps d'une génération à l'échelle cellulaire. Le cycle cellulaire (voir planche 75 du polycopié de cytologie) est subdivisé en plusieurs phases : G1, S, G2, pour l'interphase et M pour la mitose. Phase G (de gap = espace, intervalle) Elle survient dès la fin de la télophase. En raison de la variabilité de sa durée, cette phase détermine la durée du cycle cellulaire. GI = 0 pour les cellules à divisions rapides (cellules jeunes d'embryons, cellules souches de certains tissus, cellules cancéreuses). G: = durée de la vie de la cellule (G1 = G0) pour les cellules ayant perdu leur capacité mitotique (cellules musculaires, globules rouges). G, à durée variable pour les autres catégories de cellules Cette phase se caractérise par un noyau contenant 2Q d'adn. Chaque molécule d'adn correspondant à un chromosome de mitose, n'est présentée qu'en un seul exemplaire et les chromosomes ne sont pas visibles. Cette quantité d'adn reste constante en G1. Pendant cette phase, il y a aussi réactivation dans le noyau de la transcription en ARNm et de la synthèse des protéines dans le cytosol, en particulier celles qui sont nécessaires au bon fonctionnement de la cellule interphasique. Phase S (S pour synthèse) dure 6 à 8h La durée de cette phase est constante pour un type de cellule donnée. Au cours de la phase S, il y a réplication de l'adn grâce à des DNA polymérases. La quantité d'adn est donc doublée. Il y a passage de 2Q d'adn à 4Q d'adn et à la fin de la phase S, chaque molécule d'adn est représentée en double exemplaire correspondant aux deux chromatides filles du chromosome mitotique. Il y a aussi synthèse de protéines dans le cytosol, en particulier des histones qui seront associées à l'adn. Phase G2 Sa durée est également constante mais cette fois-ci le noyau est à 4Q d'adn. Le cytoplasme se prépare à la mitose avec une grande activité de synthèse protéïque: en particulier les tubulines qui serviront à la formation du fuseau achromatique. La phase G2 peut être bloquée par l'action de la colchicine qui empêche la formation des microtubules labiles ou par les rayons X. Phase M C'est la mitose qui est présentée dans les planches 141 à145 du polycopié et dans le film mitose: les 2n chromosomes deviennent visibles et seront répartis dans les deux cellules filles qui auront toutes les 2n chromosomes et 2Q d'adn grâce à la séparation à l'anaphase des chromatides sœurs. De nombreuses protéines assurent la régulation du cycle cellulaire.
 Elle survient dès la fin de la télophase. En raison de la variabilité de sa durée, cette phase détermine la durée du cycle cellulaire.")
3 LE NOYAU INTERPHASIQUE

4 DEFINITION Organite spécifique aux cellules eucaryotes, il est délimité par l'enveloppe nucléaire qui sépare son contenu du reste du cytoplasme. Il renferme le nucleoplasme dans lequel baigne essentiellement la chromatine et un ou plusieurs nucléoles. Il est le centre vital de la cellule et il contrôle grâce a l ADN toutes les activités de la cellule. L'ADN constituant essentiel de la chromatine porte les gêner du patrimoine hériditaire ou génome. I. STRUCTURE ET ULTRASTRUCTURE 1. Au Microscope Photonique (MP) Au microscope photonique le noyau a l'interphase apparaît souvent de forme sphérique et de taille variable. On le rencontre généralement en un seul exemplaire par cellule, Il existe toutefois quelques cellules plurinucléées, contenant plusieurs noyaux (Ex. ostéoclastes ou cellules osseuses) et d'autres anucléées avant perdu leur noyau au cours de leur maturité (Ex. hématies ou globules rouges). Remarque Pour mettre en évidence la chromatine, on utilise certaines techniques telles le test de Feulgen, qui permet la coloration de l ADN en violet, ou le test de double coloration de Brachet utilisant le vert de méthyle et la pyronine qui colorent respectivement l ADN en vert et l'arn en rose. 2. Au Microscope Electronique L'observation, des coupes minces au MET et au MEB avec l utilisation de différentes techniques: augmentation de contraste, cryodecapage, coloration négative associées aux traitements des images en 3 dimensions par ordinateur etc. ont permis de montrer "organisation ultrastructurale de: 2.1. L'enveloppe nucléaire (polycopié p17) C'est une portion spécialisée du Réticulum Edoplasmique (RE). Elle est formée de 2 membranes de 6nm d'épaisseur chacune (à structure trilamellaire, asymétrique et mosaïque fluide), séparées par une cavité de 10 a 50 nm en continuité avec celle du RE : l espace périnueléaire. La membrane nucléaire externe porte sur sa face cytosoilque des ribosomes et la membrane nucléaire interne est associée sur sa face nucleoplasmique a une fine couche dense aux électrons correspondant à un réseau de filaments intermédiaires constitués de lamines (voir cytosquelette): la lamina densa. Elle permet un support structural rigide a l enveloppe nucléaire et sert à la fixation des fibres de chromatine à la périphérie du noyau. L'enveloppe nucléaire permet de maintenir la forme du noyau mais assure surtout la protection du matériel génétique. Comme le REG elle est impliquée dans la synthèse de certaines protéines résidentes de l'enveloppe nucléaire et comme le REL elle est un lieu de stockage du calcium. Les complexes des pores nucléaires (polycopié p17 et Figures 1, 2 et 3) L'enveloppe nucléaire est une barrière sélective au niveau de laquelle se trouvent des zones d'interruption: les pores nucléaires. Leur nombre est variable selon le type mais surtout selon l'activité physiologique des cellules, Ils contiennent une structure complexe, le complexe du pore nucléaire (CPN). Il est constitué de 2 grands anneaux de 120nm de diamètre (l'anneau cytosolique et l'anneau nucleoplasmique) délimitant un orifice central ou transporteur central de de 30nm de diamètre. Chacun des 2 anneaux est un assemblage de 8 bras radiaires qui font saillie dans l'orifice central, délimitant 8 canaux latéraux. Un troisième petit anneau est situe à l'intérieur du nucléoplasme. Il semble qu'il existe une interconnexion très stable entre les CPN et la lamina densa, suggérant que la lamina sert aussi d'ancrage pour les CPN
 Au microscope photonique le noyau a l'interphase apparaît souvent de forme sphérique et de taille variable.")
5 Le transport passif de molécules solubles s'effectue au niveau des canaux latéraux et le transport actif de molécules plus grosses se fait par le canal ou transporteur central. L'enveloppe nucléaire permet aussi l'importation et l'exportation de molécules diverses à travers les CPN et les doubles membranes (Figure 4). Elle assure ainsi le control et la regulation des échanges entre cytosol et le nucleoplasme.
.")
6
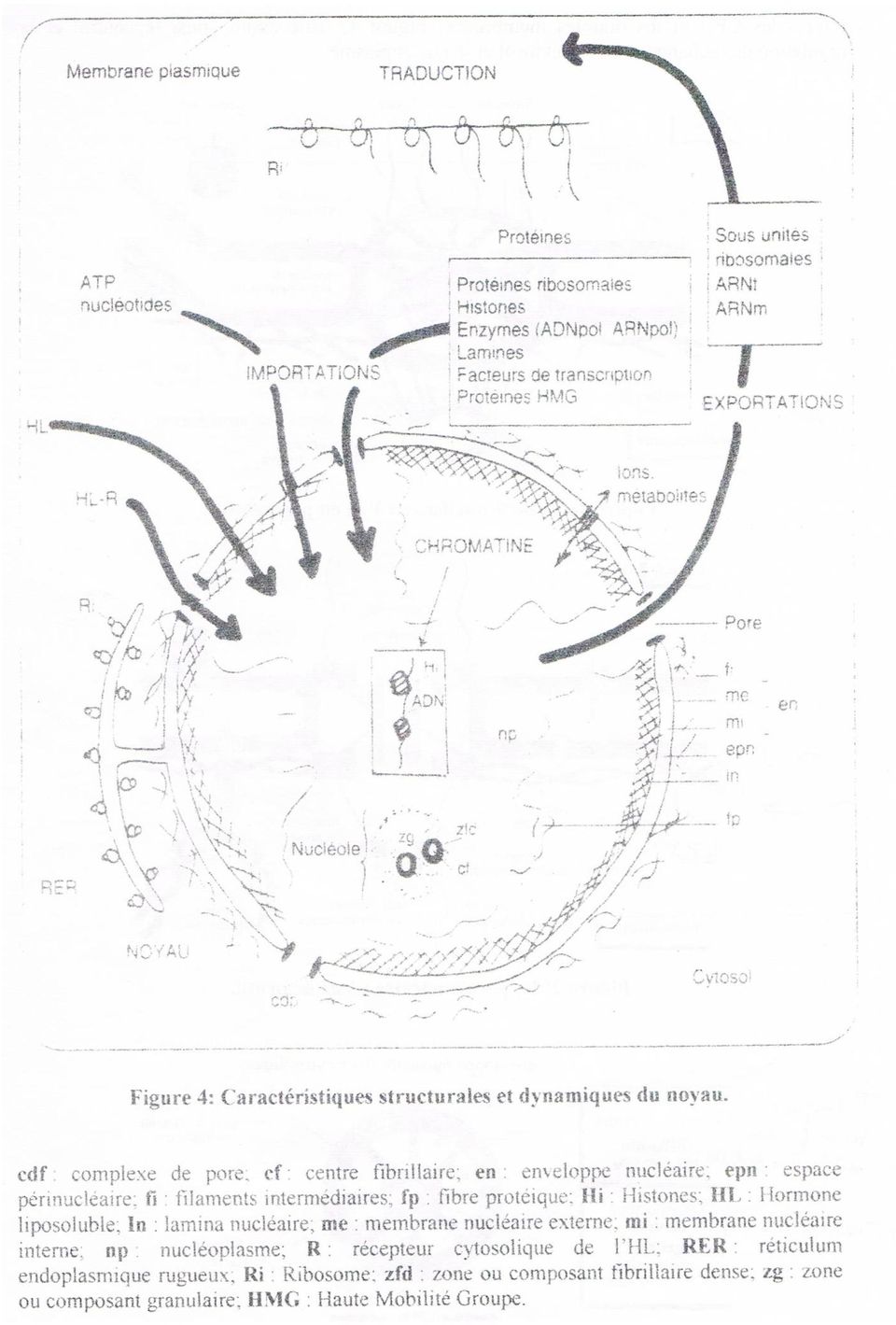
7 2.2. La chromatine (polycopié p77) Au MET la chromatine se présente sous 2 aspects: - Condensée, dense aux électrons: l'hétérochromatine située essentiellement en périphérie du noyau et un peu autour du nucléole. - Décondensée, claire: l'euchromatine, diffuse ou dispersée dans le reste du noyau. L'application de la technique de l'autoradiographie dont le principe consiste à utiliser des précurseurs radioactifs (voir TP) a montrée la synthèse d'arn au niveau de l'euchromatine et pas dans l'hétérochromatine. Ainsi l'euchromatine est la forme active de la chromatine et l'hétérochromatine la forme inactive. La chromatine intervient dans la division et la croissance cellulaire (voir cycle Cellulaire) II. ANALYSES DES DIFFERENTS CONTITUANTS L'UCD permet d'isoler la fraction noyau (PL 2 Techniques et polycopié p 35). La rupture de l'enveloppe nucléaire après action d'ultrasons ou de chocs osmotiques et un certain nombre d'ucd permettent d'isoler séparément les sous fractions du noyau: l'enveloppe nucléaire, le nucléoplasme, la chromatine et le nucléole. Les sous fractions isolées sont soumises â des analyses aussi bien utrastructurales (à fort grossissement) que biochimiques, celles-ci ont permis d'apporter un complément d'information â l'aspect ultrastructural, à la composition chimique et à l'architecture moléculaire des différents constituants. 1. Le nucléoplasme (Figure 5) C'est un gel visqueux équivalent au hyaloplasme, dans lequel se trouve une matrice nucléaire composée de minces fibrilles protéiques en réseau comportant des granules à l'intersection de ses mailles, une sorte de squelette nucléaire qui maintient la forme du noyau et agit comme un échafaudage sur lequel s'organise la chromatine. Cette matrice sert d'ancrage à l'adn et aux ARN dans les mécanismes de réplication, de transcription et de maturation Le nucléoplasme contient aussi diverses molécules: des protéines variées (enzymatiques au structurales), des ARNm et ARNt, différents types d'ions Ca ++, Na +, K + et Mg ++, des nucléotides, des sous unités ribasomales et un autre réseau protéique sous membranaire à organisation grillagée qui englobe le petit anneau nucléosomique des CPN (Figures I et 2). Figure 5: La matrice nucléaire au MET (d'après David G. Capco Katherine M.Wan et SheldonPenman). 2. La chromatine Après son isolement, son étalement et l'application de la technique de la coloration négative, on peut observer au MET à fort grossissement qu'elle est constituée de 2 types de fibres: - La fibre A de 10 à 11 nm de diamètre, organisée en collier de perle, constituant l euchromatine. - La fibre B de 25 à 30nm de diamètre, beaucoup plus condensée et plus épaisse, constituant l hétérochromatine.

8 2.1. Composition chimique Elle contient 30 à 35% d ADN ; 30 à 40% de petites protéines basiques comprenant 5 classes d'histones, H1, H2A, H2B, H3 et H4 ; 10 à 25% de protéines acides et 5 à 10% d ARN liés à l'adn au niveau de l'euchromatine. 2.2 Organisation moléculaire (Figures 6, 7a. 7h et 8) 221. La libre A ou euchromatine Dans l'organisation en collier de perles de la fibre A, les perles représentent les nucléosomes ainsi la fibre A correspond a la fibre nucléosomique (Figure 7a).Un nucléosome, disque de 10 à 11 nm de diamètre et de 6 nm de hauteur est un octamère d'histones qui comporte une paire de H2A. H2B, H3, et H4. L'ADN situé a la périphérie s'enroule autour de l'octamère d'histone. Une cinquième histone H1 intervient pour verrouiller l'adn en se liant à chaque nucléosome prés du site où l'hélice d'adn entre et sort de I'octamère d'histone. En sa présence l'adn constitue 2 tours complets. Les nucléosomes sont relies entre eux par l'adn internucléosomique. L'ADN entourant d'histones et l' internucléosomique sont constitués au total d'environ 200 paires de nucléotides. Figure 6: Le nucléosome, unité élémentaire de la chromatine La fibre B ou hétérochromatine L'histone H1 constituée par une partie globulaire et de bras correspondant aux extrémités amino et carboxy terminales intervient aussi dans l'empilement des nucleosomes en se liant grâce a sa portion globulaire à un site unique sur un nucléosome et il est supposé que ses bras se déploient pour entrer en contact avec d'autres sites sur l'octamère d'histones des nucléosomes adjacents. Les nucléosomes sont ainsi réunis en une rangée répétitive et régulière organisée en hélice, présentant ainsi une structure d'ordre supérieure en solénoïde, d'où le nom de fibre de solénoïde correspondant à la fibre B (Figure 7b). Figure 7: (a) Chromatine en collier de perles ou fibre A et (b) fibre en solénoïde ou fibre B (Barbara Hankalo et Victoria Foe).

9 2.3. Le chromosome (polycopie p85 et Figure 8) La chromatine et le chromosome sont deux états morphologiques différents d'un même matériel génétique. Au cours de la mitose la chromatine se condense de plus en plus et de façon plus complexe grâce à la participation de protéines acides qui constituent un squelette de base ( scaffold= échafaudage) autour duquel la fibre solénoïde constitue des boucles qui se condense de plus en plus pour atteindre le maximum au cours de la métaphase. A cc stade le chromosome est fois plus court que la molécule d ADN déroulée. 3. Le nucléole C'est une structure plus ou moins sphérique dans le noyau, non délimitée par une membrane (polycopié p77). En nombre défini pour chaque type cellulaire, généralement 1 ou 2 par noyau ou plusieurs (Ex, ovocytes en croissance) ou absent (Ex. spermatozoïdes), il disparaît à la prophase et se reforme à la télophase, pour persister durant toute l interphase. Au MET (Figure 9) le nucléole présente 3 parties relativement distinctes - Le centre fibrillaire CF généralement central (ou plusieurs CF) -Le composant fibrillaire dense CFD entourant le CF - Le composant granulaire CG situé en périphérie des 2 parties précédentes L agencement de ces 3 parties peut toutefois varier selon le type cellulaire (voir TP noyau) Composition chimique Le CF contient les séquences intercalaires non transcrites de L'ADN nucléolaire Le CFD contient les séquences transcrites de l'adn nucléolaire en activité, les transcrits ARNr 45S,

10 Les protéines diverses (protéines ribosomales L «Large» et S «Small» associées aux transcrits ARNr 45S, des histones, de nombreux enzymes etc. Le CG contient les ARNr en cours de maturation associés aux protéines ribosomales L et S, des enzymes et des catalyseurs qui interviennent dans la maturation comme l'arnase et les ribonucléoprotéines ou RNP (ARN + protéine), des petites et grosses sous unités ribosomales en fin de synthèse. Figure 9: Ultrastructure du nucléole Organisation du nucléole (polycopié p87 et Figure 10) Le nucléole contient de grandes boucles d'adn qui proviennent de plusieurs fibres de chromatine ou chromosomes, chacune d'elle contenant le gène d'arnr 45S amplifié: il est répété plusieurs fois en tandem (ou gène redondant) orienté dans un seul sens, séparé chaque fois par une séquence d'adn non transcrite. On appelle chaque boucle du nucléole l organisateur nucléolaire.
 Le nucléole contient de grandes boucles d'adn qui proviennent de plusieurs fibres de chromatine ou chromosomes, chacune d'elle contenant le gène")
11 3.3. Biogenèse du nucléole Au cours de la mitose ces organisateurs nucléolaires se situent au niveau des constrictions secondaires des chromosomes qui les portent (10 sur 46 dans les cellules humaines polycopié p83). La disparition du nucléole à la prophase est liée à la condensation de la chromatine pour devenir chromosomes et donc à l'arrêt de l'activité des organisateurs nucléolaires. Sa réformation à la télophase est liée à la décondensation des chromosomes qui se transforment en chromatine et à la reprise de leur activité Rôles du nucléole Biogenèse des sous unités ribosomiques La première fonction fondamentale du nucléole est la biogenèse des sous unités des ribosomes. C'est donc le lieu de formation de ces structures. Cette activité comporte 2 étapes: - Une étape transcriptionnelle: (Figure 11) Dans la zone fibrillaire dense, tous les gènes des ARNr 45S de la totalité des organisateurs nucléolaires du nucléole se mettent a transcrire des ARNr 45S dans le sen(3' ' 5)grâce à des ARN polymérases. A ces transcrits d'arnr 45S sont associées des protéines ribosomales L et S (polycopié p87), qui ont migrées dans le noyau après leur synthèse dans le cytoplasme. Les transcrits d'arnr 45S associés à leurs protéines L et S passent dans la zone granulaire tout en subissant une maturation (fragmentation) sous l'action des ARNases et des RNP, pour former différents ARNr matures: ARNr 28S, ARNr 18S et ARNr 5,8S. Remarque: Dans les préparations de la chromatine étalée (régions du nucléole) grâce à la disposition périodique des gènes des ARNr (en tandem) et leur transcription très rapide, ils peuvent être facilement observés. Les ARN polymérases et les transcrits qui leur sont associés sont groupes d'une manière si dense (environ 100/gène) que les transcrits s'écartent perpendiculairement à l'adn et donnent ainsi à chaque unité de transcription une allure caractéristique en «arbre de noël» ou plume d'oiseau. Comme pour toute transcription (quelque soit le type d'arn) le sommet de chaque arbre ou plume correspond sur l'adn au point de départ où débute la transcription et où les transcrits sont donc les plus courts, alors que l'autre extrémité de l'unité de transcription des ARNr est nettement délimitée par la disparition soudaine des molécules d'arn polymérase et de leur transcrits. Figure 11: Ultrastructure du gène amplifié d'arnr (D'après Ulrich Scheer). - Assemblage des sous unités des ribosomes: (Figure 12) L'assemblage des ARNr matures associés à leurs protéines L et S s'effectue dans la zone granulaire: l'arnr 18S et 30 à 33 protéines S constituent la petite sous unité ribosomale 40S. Les ARNr 28S et 5,8S et 40 à 50 protéines L auxquels vient s'associer un ARNr 55, synthétisé dans le noyau en dehors du nucléole (nucléoplasme), constituent la grosse sous unité ribosomale 60S. Les 2 sous unités ribosomales quittent séparément le noyau en passant par les pores nucléaires au cytoplasme, où elles s'associent grâce à l'arnm pour la synthèse des protéines Autres fonctions du nucléole Récemment, le nucléole a été impliqué dans d'autres fonctions cellulaires, comme l'assemblage ou la maturation de complexes impliquant des ARN différents des ARN.r tels le SRP (signal recognition particule = particule de reconnaissance du signal) qui intervient dans synthèse de protéines ou des ARN de transfert etc. Il intervient aussi dans le cycle cellulaire ou le vieillissement.

12 Figure 12 : Fonction du nucléole dans la synthèse des ribosomes Pour en savoir plus: 1- Alberts B., Bray D., Lewis.J., Raff M., Roberts K. et Watson J biologie moléculaire de la cellule. 2eme édition, Edit. Flammarion Medecine-Sciences. 2- Beaudouin J. et Daigle N La dynamique de l'enveloppe nucléaire. Médecine/ Science, N 1, Vol. 18, pp,4 I Cau P. et Seite R Cours de biologie cellulaire (les cours du PCEM), 3eme édition, Edit. Ellipses. 4- De Robertis E., Nowinski W.et Saez F Biologie Cellulaire, Traduction de la 5 édition de (Cell Biology, éd. Les pesses de l'université Laval. 5- Hernandez-verdun D. et Louvet E Le nucléole: Structure, fonction et maladies associées. Médecine/Science N 1, Vol.20, pp Karp G Biologie cellulaire et moléculaire (l et 2eme cycles LMD Sciences de la vie), 2eme édition, Edit. DeBoeck 7- Strachan T. et Read A.P , Génétique moléculaire humaine, Édit. Flammarion Médecine- Science. 8- Wehner R. et Gehring W Biologie et physiologie animale. 23eme édition, Edit. DeBoeck université Thieme Verlag.

13 LES RIBOSOMES I- Définition: Les ribosomes sont de petites particules compactes présentes dans toutes les cellules, en très grand nombre. Ce sont des complexes ribonucléoprotéïques majeurs de la cellule aussi bien procaryote qu'eucaryote, qui catalysent l'assemblage des AA dans un ordre prédéterminé et donc l'allongement des polypeptides = synthèse des protéines. II- Ultrastructure: Au MET ils apparaissent comme des particules globulaires distinctes denses aux e-, de 14 à 23 nm de diamètre. Ils existent dans les cellules: * soit libres dans te hyaloplasme: sous forme de deux sous unité séparées lorsque inactifs regroupés en chapelets sur l'arnm constituant des polyribosomes (ou polysomes) lorsque actifs. * Soit attachés en polyribosomes sur la face externe (face cytosolique) de la membrane des citernes du REG et de l'enveloppe nucléaire. On les rencontre aussi dans les mitochondries et les chloroplastes. III- Mise en évidence: La technique de coloration négative qui consiste à augmenter le contraste au MET par utilisation d'une substance dense aux e - tel que l'acide phosphotungstique (polycop.p.31), a permis de révéler que les ribosomes sont des édifices compacts et relativement denses, constitués de 2 sous unités de forme et de taille différentes, qui s'adaptent l'une à l'autre grâce à la présence d'une molécule d'arnm pendant leur activité qui est ta traduction. IV - Composition chimique I- Isolement (voir planche des différentes méthodes d'isolement): Broyage cellulaire homogénat 3 UCD de surnageant microsomes (fragmentation du REG en petites vésicules avec ribosomes) dans le dernier (31ème) culot -----addition de détergent au 3ième culot détachement des ribosomes des membranes des microsomes 4éme UCD à vitesse très élevée ribosomes libres des microsomes dans culot 4, UCD dans un gradient de concentration de saccharose. Les ribosomes sont ensuite caractérisés selon leur vitesse de sédimentation dans le gradient de saccharose par leur coefficient de sédimentation exprimé en unité de Svedberg ou S. 2- Résultats de l'analyse: Les ribosomes de toutes les cellules comprennent donc une grosse sous unité (ou gros élément) et une petite sous-unité (ou petit élément). Celles-ci renferment des molécules d'arn ribosomiaux (ARNr) de diverses longueurs pour 2/3 (env.65%) et un certain nombre de protéines différentes pour 1/3 (env.35%). Les protéines ainsi que les ARNr sont différents d'une sous unité à l'autre. La grosse sous unité comprend une grande molécule d'arnr principale, la petite sous unité, une petite molécule d'arnr. On désigne la taille des sous unités et leur molécule d'arnr en unité de Svedberg(S), une mesure de la vitesse de sédimentation des particules en suspension après leur ultracentrifugation dans un gradient de saccharose (Unité ni additive ni rigoureusement proportionnelle à la taille des molécules ou des

14 particules). La grosse sous unité comprend des protéines différentes entre elles représentées par la lettre L (pour large en anglais= gros), la petite sous unité, d'autres protéines, différentes aussi entre elles représentées par la lettre S (pour small=petit). Les ribosomes des cellules procaryotes et des cellules eucaryotes ont une structure et une fonction semblables. Cependant la longueur des molécules principales d'arnr,.la teneur en protéines de chaque sous unité et; par suite, la taille des éléments diffèrent entre procaryotes et eucaryotes. Un trait remarquable du ribosome est qu'on peut le reconstituer in-vitro à partir de ces constituants ribonucléoprotéiques. En effet en mélangeant dans des conditions choisies les ARNr et les protéines ribosomiales purifiées, on obtient un ribosome fonctionnel. Grosse S/U (Gros élément) Petite S/U (Petit élément) Ribosome assemblé (actif) PROCARYOTE 50S ARNr 23S+5S entre31 à 34 protéines L 30S ARNr16S 21 protéines S 70S taille réduite moins nombreux EUCARYOTES 60S ARNr28S+5.8S±5S entre 45 à 50 protéines L 40S ARNr18S entre30 à 33 protéines S 80S taille plus grande plus nombreux REMARQUE : On trouve aussi des ribosomes dans les chloroplastes et les mitochondries; les ribosomes des chloroplastes sont semblables aux ribosomes des procaryotes (70S) ; les ribosomes des mitochondries possèdent de plus petits ARNr et moins de protéines que les ribosomes des procaryotes. V -Organisation moléculaire ou structure du ribosome La microscopie électronique à haute résolution couplée à des analyses immunologiques (marquage in situ par des anticorps spécifiques) a révélé que la structure du ribosome est très volumineuse et complexe. Sa structure tridimensionnelle complète (de sa grosse S/U unité et de sa petite S/U) n'a été déterminée que récemment Elle a permis de montrer que ce sont les ARNr à configuration tridimensionnelle très précise et non les protéines ribosomiales qui sont responsables de la structure globale du ribosome, de sa capacité à positionner l'arnt sur l'arnm et de son activité catalytique pour la formation de liaisons peptidiques. Ainsi la forme générale du ribosome est déterminée par le repliement des ARNr. Les protéines ribosomiales sont généralement localisées à sa surface et remplissent les fentes et les crevasses formées par les ARNr repliés, liées par des liens non covalents aux ARNr et entre elles. Le rôle principal des protéines ribosomiales est essentiellement structural puisqu'elles semblent jouer un rôle de stabilisatrices des ARNr tout en permettant les variations de conformation des ARNr

15 nécessaires à leur catalyse efficace de la synthèse des protéines. Il a été ainsi identifié sur les ARNr les sites où viennent se fixer l'arnm et l'arnt tous en jeu dans le mécanisme de synthèse protéique. Ainsi le ribosome possède 4 sites de liaisons pour les autres molécules d'arn, situés exclusivement sur les ARNr: 1site de liaison de l'arnm situé sur ARNr du petit élément (petite sous unité) 3sites de liaison des ARNt situés en grande partie sur l'arnr du gros élément(gsu) le site de liaison de l'aminoacyl-arnt= site A, qui fixe la molécule d'arnt entrante, portant un nouveau acide aminé. Le site de liaison du peptidyl-arnt =site P, qui fixe la molécule d'arnt portant le polypeptide en croissance, c'est le site où se forme une nouvelle liaison entre 2 AA traduits. Ce site catalytique de la formation de la liaison peptidique est clairement formé par l'arnr 23S. La peptidyl transférase n'est donc pas une enzyme protéique ribosomiale mais un ARNr. Et enfin le site de liaison de l'arn t vide sortant=site E (Exit). Ainsi ce sont les molécules d'arnr qui ont un rôle catalytique comme des enzymes et non les Protéines; pour cette raison on considère les ribosomes comme étant des ribozymes V1 Fonction des ribosomes et molécules impliquées 1- Fonction : -la fonction des ribosomes est la traduction ou synthèse des protéines, dont les différentes étapes résumées sont les suivantes: La petite S/U (le petit élément) fait correspondre les ARNt aux codons de l'arnm La grosse S/U (le gros élément) catalyse la formation des liaisons peptidiques qui lient en semble les AA en une chaine polypeptidique. Les 2 éléments se rassemblent sur une molécule d'arnm à son extrémité 5' pour débuter la traduction. Le ribosome se déplace le long de l'arnm en traduisant codon par codon la séquence nucléotidique dans le sens 5 3' en une séquence d'aa, utilisant les ARNt comme adaptateurs pour ajouter l'ordre correcte de chaque AA à l'extrémité de la chaine polypeptidique en croissance. Les 2 éléments se séparent quand la synthèse de la protéine ou du polypeptide est achevée. 2. Molécules impliquées dans la traduction : les molécules impliquées dans les différentes étapes de la synthèse protéique sont succinctement: * Des molécules d'arnr constituant le ribosome et dont certaines jouent le rôle d'enzyme Une molécule d'arnm venue du noyau portant l'information génétique sous forme de codons ou triplets de bases. Des molécules d'arnt venues aussi du noyau avec l'anticodon et portant les AA activés, provenant du cytosol Les AA présents dans le cytosol Plusieurs types de facteurs cytosoliques protéines enzymatiques: + Facteurs responsables de l'activation des AA et de leur accrochage aux ARNt (différents entre procaryotes et eucaryotes). + Facteurs intervenant à plusieurs étapes de la traduction : facteurs d'initiation, facteurs d'élongation et facteurs de terminaison. - L'ATP (Adénosine triphosphate) e-gtp (guanosine triphosphate) REFEREN CES: -La cellule par Alberts Bray, Lewis, Raff, Roberts et Watson. Second édit, 1990; -L'essentiel de la biologie cellulaire: Introduction à la biologie moléculaire de la cellule par Alberts, Bray, Lewis, Raif et Walter. VA (1998), VF (1999), -Biology par Raven et Johnson. 5éme édit
 le site de liaison de l'aminoacyl-arnt= site A, qui fixe la molécule d'arnt entrante, portant un nouveau acide aminé.")
16 Biologie moléculaire de la cellule par Lodish, Baltimore, Berk, Zipursky,Matsudaira et Darnell. Deboeck université. édit Chapitre 4 p La cellule par Cooper G.MDeboeck université édit chapitre 7 p , -Biologie par Campbell et Reece. Deboeck université édit 2004.chapitre 7 p , chapitre 17 p Biologie Moléculaire de la cellule par Alberts, Johynson, Lewis, Raif, Roberts et Walter.4eme édit Médecine- Sciences Flammarion.chapitre 6 p Bologie cellulaire: des molécules aux organismes (cours, questions de révisions et QROC) par Callen J.C. (collabo. Perasso R.).2005, 2 édit. Dunod.chapitre 4, p
 par Callen J.C. (collabo. Perasso R.).2005, 2 édit.")
17 LE SYSTEME ENDOMEMBRANAIRE DEFINITION Le système endomembranaire, présent uniquement dans les cellules eucaryotes, est l'ensemble des cavités cytoplasmiques limitées par des membranes intercommunicantes entre elles par l'intermédiaire de vésicules ou canalicules. Composants du système endomembranaire Les différents compartiments du système endomembranaire sont: le Réticuhnn Endoplasmique (RE), l'appareil de Golgi, les lysosomes, les phagosomes-endosomes et la vacuole végétale. A - RETICULUM ENDOPLASMIQUE (R.E.) Définition C'est un ensemble de membranes délimitant des cavités sous forme de citernes ou des tubules. II peut être dépourvu de ribosomes, c'est le réticulum endoplasmique lisse (REL) ou porteur de ribosomes, c'est le réticulum endoplasmique granulaire (REG) qui est en relation avec l'enveloppe nucléaire (polycope p103). I- Ultrastructure de la membrane du R.E Au MET la membrane est tripartite, dépaisseur plus petite que la membrane plasmique (6nm). Au MEB, il y a présence de particules globulaires intramembranaires. II-Composition chimique 1- Technique d'isolement: Le RE, est isolé à partir du 3éme culot de l'ultracentrifugation différentielle (polycopié p37, planche technique 2). Dans ce 3ème culot le RE, forme de petites vésicules= microsomes (rugueux, lisse). Après centrifugation dans un gradient de densité de saccharose, il y a séparation entre microsomes rugueux et microsomes lisses. Microsomes rugueux + détergents, après centrifugation, permet de séparer les ribosomes des membranes. Microsomes lisses (origine REL, appareil de Golgi et membrane plasmique) après centrifugation dans un gradient de concentration de saccharose, les microsomes lisses se séparent en fonction de leur densité. 2- Résultats: a- La membrane Elle est constituée: de protéines (70%) surtout des protéines enzymatiques (glycosyl transférase, cytocbmme P450, glucose 6 phosphatase etc.), de lipides (30%), les phospholipides forment une bicouche avec un pourcentage important d'acides gras insaturés: (fluidité importante) et du cholestérol présent en liés faible quantité, de glucides. Les chaînes glucidiques attachées aux protéines et aux lipides se trouvent sur le côté luminal. Architecture moléculaire de la membrane: mosaïque fluide asymétrique (polycope p107). b- Le contenu de la cavité: Il est différent d'une cellule à l'autre, Exemples: la cavité du REG de la cellule du pancréas exocrine contient des protéines enzymatiques, celle du REL de la cellule lutéale contient des hormones stéroïdes, la cavité du réticulum sarcoplasmique de la cellule contient du Ca ++.
 Définition C'est un ensemble de membranes délimitant des cavités sous forme de citernes ou des tubules.")
18

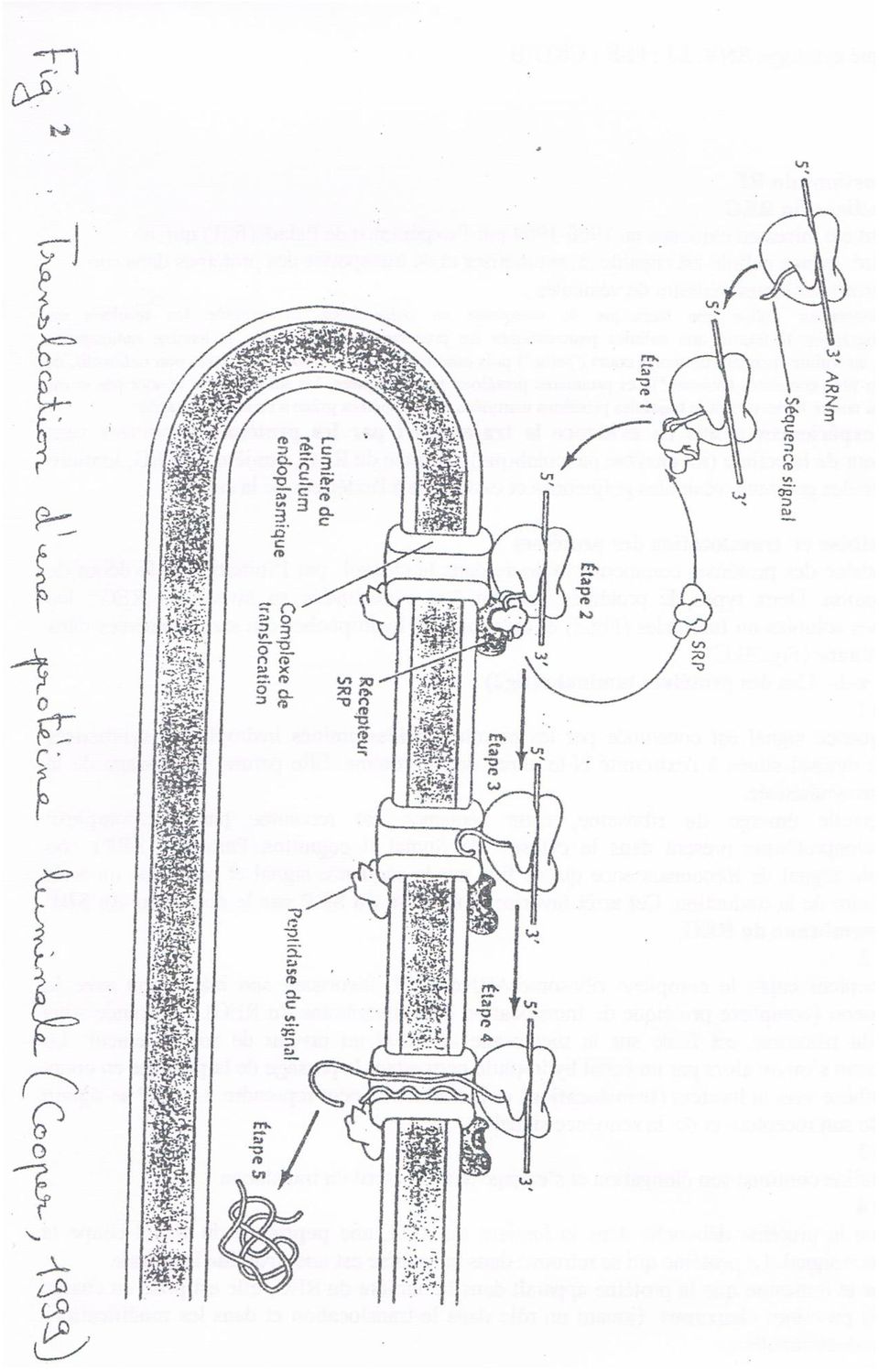
19 III- Fonctions du RE 1- Fonctions du REG Elles ont été mises en évidence en par l'expérience de Palade (fig.1) qui a démontré qu ne cellule est capable de synthétiser et de transporter des protéines dans son cytoplasme par l intermédiaire de vésicules. L expérimentateur utilise une technique de marquage en pulse-chasse et exploite les résultats en autoradiographie Il fournit aux cellules pancréatiques un précurseur de protéines: la leucine radioactive, marquée au tritium pendant un temps court (" pulse") puis ensuite fournit le même précurseur, non radioactif, et beaucoup plus concentré ("chasse"). Les premières protéines sont marquées, les suivantes ne le sont pas et on peut ainsi suivre le devenir et le trajet des protéines marquées et les repérées grâce à l autoradiographie. Cette expériences a mis en évidence le trajet suivi par les protéines exportées vers l'extérieur de la cellule (ici enzyme pancréatique): surface du REG, lumière du REG, lumière des saccules golgiens, vésicules golgiennes et exocytose â l extérieur de la cellule. a - Synthèse et translocation des protéines La synthèse des protéines commence toujours dans le cytosol par l'initiation et le début de l'élongation. Deux types de protéines peuvent être synthétisées au niveau du REG: les protéines solubles ou luminales (Fig.2) et les protéines hydrophobes qui seront insérées dans la membrane (Fig.3B.C). a-1- Cas des protéines luminales (fig.2) Etape l La séquence signal est constituée par les premiers acides aminés hydrophobes synthétisés dans le cytosol situés à l extrémité N-terminal de la protéine. Elle permet l'adressage de la protéine synthétisée. Dès quelle émerge du ribosome, cette séquence est reconnue par un complexe ribonucléoprotéique présent dans le cytosol: la Signal Recognition Particule (SRP) ou Particule Signal de Reconnaissance qui se fixe sur la séquence signal et provoque un arrêt temporaire de la traduction. Cet arrêt favorise la fixation du SRF sur le récepteur du SRP de la membrane du REG. Etape 2 Ce récepteur capte le complexe ribosome-arnm-srp, favorisant son interaction avec le translocon (complexe protéique de translocation de la membrane du REG). La grande sous unité du ribosome est fixée sur la membrane du REG au niveau de son récepteur. Le translocon s'ouvre alors par un canal hydrophile permettant le passage de la protéine en cours de synthèse vers la lumière (translocation) et la traduction peut reprendre. La SRP se sépare alors de son récepteur et de la séquence signal. Etape 3 La protéine continue son élongation et s'engage dans le canal du translocon. Etape 4 Lorsque la protéine débouche dans la lumière du REG, une peptidase du signal coupe la séquence signal. La protéine qui se retrouve dans la lumière est une protéine luminale Au fur et à mesure que la protéine apparaît dans la lumière du REG, elle est prise en charge par des protéines chaperons (jouant un rôle dans le translocation et dans les modifications posttraductionnelles). a-2- Cas des protéines membranaires (fig3 B.C) Les protéines destinées aux membranes subissent aussi un adressage par la séquence signal. Mais la translocation des protéines destinées à la membrane est bloquée par une séquence très hydrophobe qui déstabilise le translocon et la protéine reste insérée dans la membrane du REG: c'est une protéine membranaire.
 puis ensuite fournit le même précurseur, non radioactif, et beaucoup plus concentré (\"chasse\").")
20
21
22 b-modifications post traductionnelles des protéines b-1- N- glycosylation Au cours de leur synthèse, les protéines peuvent être glycosylées : un "arbre oligosaccharidique" très riche en mannose est transféré en bloc sur la protéine en cours de traduction par une oligosaccharidyl transférase sur le groupement amine de l asparagine. b-2 Modifications des sucres Cet arbre oligosaccharidique est aussitôt modifié par les enzymes du REG. b-3- Etablissentent de ponts disulfures entre acides aminés soufrés. b-4- Association entre protéines et lipides membranaires (cas des protéines intégrées ancrées dans l'une ou l'autre des bicouches lipidiques (voir Fig.1 de membrane plasmique). e- Sortie du REG par un transport vésiculaire Ce sont des vésicules qui bourgeonnent du REG et qui vont fusionner avec la face cis de l'appareil de Golgi. Ces vésicules ont une membrane issue du REG et peuvent donc transporter les protéines luminales et les protéines membranaires intégrées. Le compartiment intermédiaire entre REG et appareil de Golgi est appelé Endoplasmic Reticulum-Golgi intermediate Compartment ou ERGIC (Fig.4). En fonction de certaines séquences, les protéines transmembranaires ou luminales dites "résidentes du REG (spécifiques du REG) ne peuvent sortir du REG. Si elles ont été transportées par erreur dans I'ERGIC par erreur, leur retour vers le REG est assuré. 2-Fonctions du REL: a- Synthèse des phospholipides Les phospholipides sont synthétisées grâce aux enzymes transmembranaires du REL dont le site actif est orienté vers le cytosol. Les lipides sont ensuite répartis dans les bicouches par des flipases, intégrées dans la membrane du REL, permettant ainsi leur basculement de I'hémi couche cytosolique vers l'hémicouche luminale. Le REL est le principal fournisseur de phospholipides pour les membranes cellulaires. b-synthèse des hormones stéroïdes à partir de la prégnénolone synthétisée dans la mitochondrie grâce au cytochrome P450. c- Détoxification Grâce au cytochrome P450, il y a hydroxylation des produits toxiques. Devenus hydrosolubles, ces produits seront facilement transportés et éliminés. d-accumulation et libération de Ca++ B- APPAREIL DE GOLGI (A.G.) Définition C'est un organite situé au voisinage du noyau, proche du matériel péricentrosomal. II est constitué par des empilements de saccules aplatis, associés à de nombreuses vésicules (poly 103).
23 1-Structure et Ultrastructure a- Structure: Au microscope photonique, après imprégnation au nitrate d'argent, Golgi a mis en évidence de petites écailles à proximité du noyau. b- Ultrastructure: b-1- organisation: Au M.E.T, il y a empilement de saccules (comme une pile d'assiettes) et des petites vésicules. Chaque empilement de 4 à 8 saccules est un dictyosome. En moyenne on compte environ 20 dictyosomes par cellule. Chaque dictyosome comporte des saccules cis (proches du RE), face d'entrée alimentée par le RE, des saccules médians et des saccules trans (face de sortie) en continuité avec un réseau de canalicules appelé le réseau trans- golgien ou T.G.N. (Trans Golgi Network) (Fig.4). b-2-- Caractéristiques des membranes golgiennes: Au MET membrane tripartite, au MEB présence de particules globulaires intégrées. L'épaisseur de ces membranes est variable et intermédiaire entre celle du REG et celle de la membrane plasmique (saccule cis 6 nm et saccule trans 7,5 nm). II- Composition chimique 1- Technique d'isolement (polycop. p37 et 103, Planche technique 2) Broyage brutal d'un fragment d'organe + U.CD. -> le 3ème culot contient des microsomes (voir R.E.), broyage aménagé (doux) + U.C.D. - le 3èm culot contient des saccules empilés. 2- Résultats de l'analyse chimique a- Membranes Lipides: taux intermédiaires entre celui de la membrane de RE et celui de la membrane plasmique. La longueur des chaînes d'acide gras, leur degré de saturation et par conséquent leur fluidité sont également intermédiaires entre membrane du RE, et membrane plasmique. Protéines: pourcentage intermédiaire entre le % des protéines de la membrane du RE et le % des protéines de la membrane plasmique. On distingue les protéines communes aux membranes du RE et de l'a.g. (glycosyl- transférases, cytochrome b5 réductase) et des protéines particulières aux membranes de 1 A-G. (les sulfo-transférases, les phosphatases). Glucides : la quantité est négligeable et situés sur la face luminale. Architecture moléculaire des membranes: Ce sont des mosaïques fluides asymétriques. b- Le contenu des cavités: Mêmes produits que ceux des cavités de RE, à des concentrations différentes. III- Fonctions (fig. 4) 1- Transport L ERGIC délivre les protéines et les glycoprotéines, les lipides et les glycolipides au cis-golgi qui sont ensuite transportés à travers l'appareil de Golgi, où ils passent du cis-golgi au Golgi médian puis vers le trans-golgi pour finalement atteindre le trans-golgi network (TGN). Ce transport est effectué grâce à des vésicules qui bourgeonnent des différents compartiments.
24 Mannose = ose Glc NAc = N acetylglucosamine Gal= galactose NANA= Acide N Acétyl Neuraminique
25 2-Gycosylation définitive des protéines a- Modifications des sucres Au cours du transport du REG vers le saccule cis et le saccule médian, les glycoprotéines subissent des modifications de leurs arbres oligosaccharidiques N liés. b- 0-Glycosylation Il y a construction d'oligosasaccharides O-liés dans les saccules médians et trans: ces oligosaccharides sont bâtis, oses par oses sur la protéine au niveau du groupement hydroxyle d un résidu sérine ou thréonine, grâce aux glycosyltransférases. 3-Phosphorylation du Mannose 6P dans les citernes cis. Cette étape ne s'effectue que pour les glycoprotéines (enzymes) destinées aux lysosomes. 2-4-Sulfatation dans les citernes trans. Il y a addition d'un groupement SO 4 sulfotransférases. grâce aux 5-Modification des lipides Certains lipides assemblés au niveau du REL sont modifiés pour former les glycolipides. 6- Stockage et libération du calcium 7-Formation des vacuoles de macroautophagie (voir 1ysosomes) La citerne membranaire encerclant l'organite à détruire peut provenir de citerne issue du trans, soit du REL. 8- Tri et maturation (Fig. 5 et 6) Ces deux phénomènes s'effectuent en même temps. Le tri se déroule au niveau du réseau transgolgien (TGN). Les vésicules en formation sont pourvues d un manteau cytoplasmique formé par des protéines et sont appelées vésicules recouvertes. Ce manteau reconnaît les signaux portés par la substance à transporter et permet sa concentration dans une zone restreinte (tri). De plus il permet le bourgeonnement de la membrane en vésicules. Les vésicules peuvent suivre deux voies: a- Voies de sécrétion (Fig.5et 6) La voie de sécrétion constitutive permet de déverser continuellement vers l'extérieur (par exemple sécrétion permanente des protéines de la matrice extra cellulaire). Ces vésicules sont recouvertes par un manteau de coatomères. - La voie de sécrétion régulée: le matériel de sécrétion soit hormonal (ex. insuline) ou de type exocrine (ex. enzymes du pancréas), subit une maturation au cours de son transport dans des vésicules recouvertes d'un manteau de clathrine et de protéines d'adaptation. Les vésicules augmentent de taille et forment des grains de sécrétion mâtures. Ce type de sécrétion après un signal selon les besoins. b- Voie endosomale, lysosomale Elle concerne les protéines pourvues d'un mannose 6P (M6P), mannose qui a été phosphorylé au niveau du saccule cis. Ces protéines luminales M6P reconnaissent un récepteur membranaire et l'ensemble est adressé aux lysosomes en passant par un compartiment intermédiaire endosomal caractérisé par un pli relativement acide 5 à 6. Les vésicules, déstabilisées par le ph acide perdent leur récepteur mannose 6P qui est recyclé vers le TGN. On obtient une vésicule, dépourvue de récepteur M6P, contenant des hydrolases acides et dont la lumière est à ph acide. Quelle que soit la voie suivie par les vésicules, elles perdent leur manteau avant d'atteindre leur destination.
26
27 Les vésicules dont la membrane est riche en cholestérol, en phospholipides en glycoprotéines liées à un GPI sont recouvertes de cavéoline, conservent ce manteau de cavéolin et par exocytose, elles insèrent dans la membrane plasmique des microdomaines riches en lipides (Fig.5). c- L'adressage des vésicules C'est un phénomène complexe moléculaire en voie de découverte faisant intervenir de nombreuses protéines. V1- Conclusion: Biogenèse membrane plasmique (Polycop. P107)
28 C- Compartiment endosomal Le compartiment endosomal (polycopié p.61) étudié en partie dans les échanges avec déformation de la membrane plasmique, fait partie du système endomembranaire. Les premiers endosomes formés juste après l'endocytose, proches de la membrane plasmique, sont appelés endosomes précoces (Fig.8). Leur fonction consiste à trier les molécules ingérées et dans le cas de l'endocytose par récepteurs ils jouent aussi un rôle dans le recyclage des récepteurs vers la membrane plasmique (CURL). Les endosomes qui contiennent la molécule triée, rapprochés du noyau, sont appelés endosomes tardifs (Fig.8). Ils évoluent en lysosomes. D- Compartiment lysosonial Définition: Découverts pour la première fois par De Duve grâce à leur fonction, ce sont des compartiments du système endomembranaire de forme variable, Ils renferment des enzymes bydrolytiques (hydrolases acides) permettant la digestion de particules et de molécules extracellulaires ingérées ou intracellulaires de petites tailles solubles et d'organites cellulaires vieillis ou inutiles. I- Ultrastructure au MET Ils apparaissent comme des vésicules de formes et de tailles variables de 0.05 à 0.5µm de diamètre selon le matériel ingéré. Ils peuvent être identifiés par des techniques cytochimiques grâce à une enzyme présente dans leur membrane la pbosphatase acide qui forme un précipité de phosphate de plomb dense aux e- (voir TP N 7). II- Composition chimique 1- technique d'isolement (Planche technique 2) Membrane lysosomale 2 UCD Choc osmotique Homogénat cellulaire Lysosomes 2 sous fractions Culot 2 UCD Contenu cavité lysosomale 2- Résultats a- la membrane C'est une membrane particulière car la plupart de ces protéines sont inhabituellement hautement glycosylées du coté luminale pour la protéger des hydrolases acides contenues dans la cavité. Les protéines membranaires sont de différents types (Fig.7): - des protéines enzymatiques (ex pbosphatase acide) et non enzymatiques ainsi que des perméases, transporteurs des matériaux à dégrader de petites tailles du cytosol et des produits terminaux de la digestion de macromolécules (ex. A.A., nucléotides, sucres simples etc...). - Des ATPases à protons (H + ) qui pompent les H + dans la cavité du lysosome, maintenant son PH très acide. b- Contenu de la cavité Elle contient environ 40 types d'enzymes digestives hydrolytiques (hydrolases acides), qui dégradent les protéines, les acides nucléiques, oligosaccharides et les phospholipides. Leur activité est optimale à ph5.
29 Figure 7 : les constituants d'un lysosome III- Origine des matériaux à dégrader et biogenèse «possible*» des lysosomes (Fig.8 et polycopié p 119) Les matériaux à dégrader suivent en fonction de leur origine des voies différentes vers le stade lysosome. ils peuvent avoir 2 origines: 1- ExtraceIluIaire : voie hétérophagie Dans ce cas, ils sont ingérés par phagocytose ou par endocytose. Ils sont alors contenus dans des phagosomes ou dans des endosomes précoces à ph neutre (7.4).Cependant leur ph diminue progressivement à cause des ATPases à H + apportées par la fusion de vésicules golgiennes (appelées anciennement lysosomes primaires), provenant du TGN et renfermant dans leur cavité quelques hydrolases acides. Les endosomes précoces se transforment alors en endosomes tardifs à ph 6.5. De nombreuses vésicules golgiennes affluant du TGN continuent à fusionner avec les endosomes tardifs, augmentant ainsi le nombre des ATPases à H + et des hydrolases acides, permettant alors la conversion des endosomes tardifs en lysosomes à ph5. Des vésicules golgiennes à membrane riche en ATPases à H + et en hydrolases acides dans leur cavité fusionnent avec le phagosome qui se convertit aussi mais directement en lysosome à ph5. *Remarque : La transformation des phagosomes et des endosomes tardifs en lysosomcs est hypothétique. 2-Intracellulaire: voies autophagiques a- Micro autophagie Entrée directe dans le lysosome de petites molécules solubles du cytosol (ex peptides) via des perméases. b- Macro autophagie Création d'autophagosome (ou vacuole autophagique) formé par l'englobement (séquestration) d'organite(s) et d'un peu de cytosol par une citerne membranaire provenant de citerne soit du REL ou de l'appareil de Golgi (saccule Trans ou du TGN). Quelque soit son origine, cette citerne membranaire contient dans sa cavité quelques hydrolases acides qui deviennent actives après changement du ph intra cavitaire, grâce semble-t-il à la présence d'atpases à H + dans la membrane de la citerne. La digestion débute donc dans l'auto phagosome et s'achève dans le lysosome avec lequel il a fusionné ou en quoi il s'est converti. e- Crinophagie C'est une forme d'autophagie qui concerne l'élimination des grains de sécrétion (sécrétion régulée) qui ne sert plus ex. prolactine.
30
31 IV- Devenir des lysosomes (Polycopié p. l19) Les lysosomes âgés, dépourvus d'hydrolases fonctionnelles, constituent des corps résiduels. Le plus souvent ces corps résiduels libèrent leur contenu à l'extérieur de la cellule par exocytose: c'est la défécation cellulaire. Mais ils peuvent aussi persister dans la cellule toute sa vie. V- Fonction des lysosomes (Fig.9) Toutes les familles de molécules biologiques sont dégradées en métabolites élémentaires qui repartent vers le cytosol grâce à des protéines transporteurs de la membrane lysosomale. Ainsi les lysosomes jouent un rôle fondamentale dans la digestion cellulaire, la destruction des corps étranger et donc dans la nutrition cellulaires entre autres fonctions. Pour en savoir plus Ouvrages BASSAGLIA Y., Biologie cellulaire, Editions Maloine, Paris: CAU P. et SEITE R., Cours de biologie cellulaire. Editions Ellipses, Paris: Campbell J. et Reece M., biologie. Second édition, De Boeck université. Karp G biologie cellulaire et moléculaire. Second édition, De Boeck université MAILLET M., Biologie cellulaire, Editions Masson Paris. Articles ANTONY B., Contrôle de l'assemblage des manteaux protéiques COP par les petites protéines G Arf et Sar. MIS, 18: BENICHOU S; BENMERAH A., 2003-Protéines Nef du VIII et dk3/k5 du virus associé au sarcome de Kaposi: des "parasites" de la voie d'endocytose. M/S, 19: COURVALIN J.C., RABOUILLE C., Réorganisation des compartiments intracellulaires membranaires pendant la mitose. M/S, 18: GALLI T., MARTINEZ-ARCA S., PAUMET F., Mécanisme de la fusion membranaire. M/S, 18: TIXIER-VIDAL A., Les compartiments membranaires de la cellule eucaryote. M/S,
32 E- LA VACUOLE I- Définition La plupart des cellules végétales contiennent une ou plusieurs vésicules très grosses remplies de liquide appelées vacuoles. L'ensemble des vacuoles d'une cellule constitue l'appareil vacuolaire ou vacuome qui peut occuper selon le type cellulaire 30 à 90 % du volume cellulaire. Il fait parti du système endomembranaire. II- Structure et ultrastructure 1- au MP: La vacuole est limitée par une membrane continue appelée tonoplaste; celui-ci délimite une solution interne dite suc vacuolaire (colorée en rose ou autres couleurs selon le ph ou incolore). 2- au MET: Le tonoplaste est une membrane biologique, à structure trilamellaire, asymétrique (le feuillet dense cytosolique est plus mince que le feuillet dense luminale: présence d'un revêtement fibreux). 3- au MEB: Présence de particules globulaires après cryodécapage. III- Composition chimique 1- Isolement : tissu végétal plasmolysé protoplastes vacuoles digestion des parois choc osmotique + centrifugation ou thermique UCD.. vacuoles éclatées 2 sous fractions : tonoplaste et suc vacuolaire, Centrifugation 2- Résultats de l'analyse chimique: 2-1- le tonoplaste: 3 types de molécules (lipides, protéines et glucides) (Fig. 1). - lipides: phospholipides et stérols (stigmastérol et sitostérols). - protéines : grande diversité (structurales, enzymatiques et transporteurs). - glucides: revêtement fibreux (chaînes polysaccharidiques), non considéré comme le cell coat (g1ycocalyx) le suc vacuolaire: contient une grande diversité de composés formant avec l'eau des solutions aqueuses dont la concentration varie en fonction de l'espèce végétale, de la fonction et de l'état physiologique (fruit, graine, fleur...) de la cellule, exemple: le grain d'aleurone présent dans la graine, riche en protéines cristallisées ( Polycopié p. 121). 1V- Architecture moléculaire (Fig. 1) Fig. 1: Constituants et architecture moléculaire du tonoplaste. AP, ATPase; EV, groupe enzymatique vectoriel; P, pore protéique-, T1 et T2, transporteurs spécifiques ; PP, protéines périphériques ; RC, relargage et absorption cytosolique; RV, relargage et absorption vacuolaires.
33 V- Biogenèse de la vacuole Les provacuoles dérivent du réticulum endoplasmique lisse. Celui-ci peut entourer une portion de cytosol qu'il va détruire grâce â des enzymes hydrolytiques (hydrolases acides). Seules persistera la membrane externe qui deviendra la membrane de la vacuole. Ce mode de formation des vacuoles par autolyse cellulaire serait assez répandu et diffère d'un modèle proposé anciennement où l'on considérait les vacuoles comme des dilatations du réticulum. Les provacuoles fusionnent pour donner une ou plusieurs vacuoles. VI- Rôles physiologiques (Fonctions) 1- Transport, fonctions communes à tontes les membranes biologiques: le tonoplaste intervient dans les échanges avec le cytosol (Fig. 2) par: 1-1- Transport passif: -la diffusion simple à travers la bicouche lipidique. -Transport par diffusion facilitée de l'eau (plasmolyse et turescence) à l'aide d'aquaporines (très nombreuses) et d'ions par les canaux ioniques Transport actif: -Pompe à proton (ATPase-H +, symports et antiports à protons. -le transport spécifique du saccharose qui couple le transport de ce composé à sa synthèse à partir de ses deux précurseurs à l'aide d'un complexe enzymatique transmembranaire à sens unique vectoriel (cytosol suc vacuolaire) Transport avec déformation: ex pinocytose intravacuolaire. Fig. 2 : Mécanismes de transport à travers le tonoplaste 2- Fonctions caractéristiques de la vacuole: 2-1-Stockage: La vacuole est un lieu de stockage d'une grande variété de substances aussi bien minérales qu'organiques. Certaines sont réutilisables par la cellule (ions, acides aminés, sucre,...) d'autres toxiques pour la cellule sont piégées. Par exemple, contrairement à la cellule animale qui excrète ses déchets toxiques, la cellule végétale utilise la vacuole pour isoler ses déchets du cytoplasme Les substances réutilisables sont transportées par diffusion (Fig. 2 et 3) Piégeage: Les substances toxiques ayant franchi le tonoplaste sont soumises à différents types de modifications qui les empêchent de ressortir (Fig.3);
34 - l'ionisation: concernant les molécules lipophiles (ex. alcaloïdes). - le changement de conformation moléculaire et la glycosylation (hétérosides). - la cristallisation (ex. en oxalate de calcium, car le calcium et l'acide oxalique son toxiques pour la plante). - la formation de liaisons avec d'autres composés et accumulation (polyphosphate Mg2 +, mucilages, tanins, polyphénols). Fig. 3 : Mécanismes de stockage et de piégeage intravacuolaires Support mécanique: Grâce â la turgescence la vacuole fournit un support mécanique aux tissus mous (support dressé des plantes herbacées) Croissance cellulaire: la croissance cellulaire en largeur et en longueur se fait grâce à la turgescence de la vacuole. 3- Fonctions communes au lysosome: La vacuole (phytolysosome) est assimilée au lysosome (Polycopié p. 121), - autophagie: les hydrolases du suc vacuolaire semblables â celles des lysosomes sont capables de dégrader tous les substrats, y compris tout le contenu cytoplasmique (ex. cellules mortes du suber, bois,... voir cours histologie végétale). - hétérophagie: est un moyen de- défense en cas d'attaque parasitaire (bactéries, champignons,...). Pour en savoir plus: 1- ROBERT D. & ROLAND J.C., Organisation cellulaire, Tome I, Ed. Dom. 2- COULOMB P. & COULOMB C., Le phytolysosome, Annales de Sciences Naturelles, Botanique, série 13, Vol. 5, pp CALLEN J.C. & PERASSO R, Biologie cellulaire, des molécules aux organismes, 2ô Ed. Dunod. 4- KARP G., Biologie cellulaire et moléculaire, 2è Ed. De Boeck. 5- ALBERTS B., JOHNSON A., LEWIS J., RAFF M., ROBERTS K. & WALTER P., Biologie moléculaire de la cellule, Ed. Flammarion Médecine-Sciences. 5-GOURRET J.P., Cellules végétales, Documents de microscopie électronique, tome 2, Publication de l INRAP, Dijon
35 ORGANITES SEMI AUTONOMES
36 LES ORGANITES SEMI AUTONOMES MITOCHONDRIE CHLOROPLASTE Généralités: Les mitochondries sont des organites présents dans toutes les cellules eucaryotes aérobies Les plastes, en particulier les chloroplastes sont présents dans les cellules végétales. Les mitochondries e les chloroplastes sont dits organites semi-autonomes c.à.d, en partie indépendants de l ADN nucléaire, car ils contiennent leur propre ADN et par conséquent, ces organites sont capables de synthétiser une partie de leurs protéines. MITOCHONDRIE DEFINIT1ON La mitochondrie est un organite cellulaire possédant une double membrane. Elle est responsable de la production sous forme d'atp de la majeure partie de l énergie necessaire à la cellule respiration). Elle possède son propre génome (ADN mit), elle est le siège des étapes indispensable au métabolisme des différentes molécules biologiques. I- STRUCTURE ET ULTRASTRUCTURE: (voir TP, et polycopié Pl. 125). L ensemble des mitochondries dans une cellule constitue le chondriome coloré au Vert Janus en MP (1700 mitochondries dans un hépatocyte). Les membranes mitochondriales ont une épaisseur moyenne de 6nm, elles sont trilamellaires et asymétrique (asymétrie plus importante au niveau de la membrane interne). II COMPOSITION CHIMIQUE II-1- Isolement des fractions et sous fractions *Fractions mitochondries 2 centrifugations centrifugation en gradient saccharose Homogénat cellulaire Culot (mitochondries, lysosomes et peroxysomes) Mit entières *Sous fractions mitochondriales II- 2- Résultats de l analyse chimique II Membranes mitochondriales (voir Fig. I) Les principaux constituants chimiques des membranes de la mitochondrie sont les lipides et les protéines.
37 Constituants membrane mitochondriale externe membrane mitochondrial interne -Taux 40% -Taux : 20% Lipides -Phospholipides -Phospholipides -Peu de cholestérol -Pas de cholestérol Protéines caractéristiques -Taux : 60% *Perméase passives : Porines (molécules PM < 10000) * Enzymes du métabolisme des lipides (ex: acétyl-coa synthétase) *Complexe transporteur (molécules PM> 10000) *Complexe récepteur.(ex récepteurs ADN et ARN polymérase) * Complexe d'entrée du cholestérol Très perméable -Taux: 80% * 4 Complexes enzymatiques de la chaine respiratoire complexe I: NADH déshydrogénase * Complexe II :succinate déshydrogénase Complexe III: complexe cyt. b-cl ' Complexe IV: complexe cyt oxydase * Perméases spécifiques actives (transporteurs ATP, ADP, pyruvate..). *Complexe enzymatique ATP synthétase * Cyt. P450 (synthèse hormones stéroïdes) Perméabilité Organisation de ATP synthétase: Complexe F0-F1 Les ATP synthétases (ATPases) au niveau de la membrane interne ont été mises en évidence par la technique de coloration négative (voir TP). L'ATPase est un complexe enzymatique formé d'une quinzaine de polypeptides, on distingue 2 parties: Un pédoncule F0 (facteur F0), hydrophobe, intégré dans la bicouche de la membrane Interne Une sphère ou partie globulaire F1, hydrophile, fait saillie dans la matrice, c'est la partie catalytique du complexe appelé aussi facteur de couplage (asymétrie de la membrane interne). Architecture moléculaire de la mb Interne (voir polycopié pl 125) II-2-2. Contenu de l'espace intermembranaire: Comme la membrane mitochondriale externe est très perméable, la composition de l'espace intermembranaire est voisine du hyaloplasme. C'est un lieu de transit pour toutes les molécules PM 10000, il contient des protons H+, ADP, ATP, enzymes II Matrice mitochondriale Granules denses aux e-: cations Ca 2+, Mg 2+, mitoribosomes, ADN en plusieurs copies (circulaire, double hélice, absence d'histones), ARNm et ARNt, nombreuses enzymes (β oxydation des acides gras, cycle de Krebs, duplication, transcription, et traduction) et de nombreux métabolites. III - Fonctions de la mitochondrie (voir poly. PI. 127, 129 et 137) III-1. Synthèse d ATP La fonction principale de la mitochondrie est la formation d'atp par un mécanisme de phosphorylation oxydative. Ce mécanisme utilise 1'O 2 dissout et s'accompagne de la formation du CO 2 permettant ainsi de définir la respiration cellulaire. La synthèse d'atp est décomposée en plusieurs étapes:
38 Figure 1. les principaux constituants chimiques des membranes de mitochondries (Cau et Seite, 1996) La membrane interne ne permet qu un passage actif de ces diverses molécules, ce qui la différencie de la membrane externe. Figure 2. Organisation de la membrane mitochondriale interne (Callen, 2005) Les trois gros complexes d'oxydoréduction protéiques notés I, III et IV, chassent les protons vers l'espace intermembranaire et participent à la création du gradient de protons qui est exploité par I'ATP synthétase pour fabriquer de 1'ATP Deux transporteurs mobiles sont impliqués le coenzyme Q (qui diffuse dans la bicouche) et le cytochrome c, situé dans l espace intermembranaire. La succinate-déshydrogénase est également représentée ( complexe II).
39 I ère étape: dans le cytosol Les produits de la digestion (acides amines, sucres simples et acides gras) pénètrent dans le cytosol ou se fera leur catabolisme (acides aminés et glucose sont dégradés en pyruvate). 2 ème étape: dans la matrice, formation de l acétyl-coenzyme A Le pyruvate et les acides gras sont convertis en acétyl-coenzyrne A ce dernier entre dans le cycle de Krebs Ce cycle produit du NADH du FADH 2, du GTP, du CO 2. 3 ème étape: dans la membrane mitochondriale interne, la chaîne respiratoire ( Fg. 2) Les e - transportés par NADH et FADH 2 sont transférés à O 2 par l'intermédiaire des complexes enzymatiques de la chaine respiratoire. Le complexe I (CI) porte d entrée des e- portés par le NADH présent dans la matrice. Le CII, porte d entrée des e- portés par le FADH 2. Les e- sont transportés entre les complexes par 2 molécules de petite taille. Ubiquinone (ou coenzyme Q) entre CI (ou CII et CIII, le Cytochrome c entre CIII et C IV. Les complexes I, III et 1V transportent les e- à l'intérieur de la membrane mitochondriale interne et les H + de la matrice vers l'espace intermembranaire, le CII transporte uniquement des e-. Le C IV (cytochrome oxydase) accepteur final de la chaine respiratoire, transfère les e- à l O2 au niveau de la matrice et production H2O. Phosphorylation de I ADP en ATP : Le transport de H+ dans l espace intermembranaire provoque I 'apparition d'un gradient de H+ entre les 2 faces de la membrane interne. L ATPase utilise le gradient de H+ pour produire de l ATP. Les H+ passent de l'espace intermembranaire vers la matrice à travers l ATPase (canal à protons) La phosphorylation oxydative ne peut s'observer que si la membrane interne délimite un espace clos L AT P formé est stocké dans la matrice ou transporté dans le cytosol vers les sites de son utilisation, le CO 2 et H 2 O produits quittent la mitochondrie de manière passive à travers les 2 membranes. III-2. Synthèse protéique : (pi. 137) La présence d ADN et de mitoribosomes permet à la mitochondrie dé synthétiser une partie de ses protéines. Ex cytochrome b codé par le génome mitochondrial. ÀTPase (une partie codé par ADN mitochondrial l'autre partie codé par ADN nucléaire III.3. Synthèse de nombreux précurseur (pl 127) La mitochondrie assure la synthèse des précurseurs de la biogenèse des acides gras et des aminées, des glucides, de l urée.. Dans le cas des cellules élaborant les hormones stéroïdes, la membrane mitochondrial interne participe grâce au cytochrorne P450 (voir REL). Pour en savoir plus ROBERT D., et VlAN B Eléments de biologie cellulaire. MULLER Y., et CLOS J Organisation fonctionnelle de la cellule (tome I). CAU P., et SEITE R., Biologie cellulaire. Editions ELLIPSE: g. CALLLN J.C.2005.Biologie cellulaire: des molécules aux organismes 2 ème édition Danod..
ULBI 101 Biologie Cellulaire L1. Le Système Membranaire Interne
 ULBI 101 Biologie Cellulaire L1 Le Système Membranaire Interne De la nécessité d un SMI Le volume augmente comme le cube de la dimension linéaire, alors que la surface n'est augmentée que du carré Une
ULBI 101 Biologie Cellulaire L1 Le Système Membranaire Interne De la nécessité d un SMI Le volume augmente comme le cube de la dimension linéaire, alors que la surface n'est augmentée que du carré Une
Chapitre 7 : Structure de la cellule Le noyau cellulaire
 UE2 : Structure générale de la cellule Chapitre 7 : Structure de la cellule Le noyau cellulaire Professeur Michel SEVE Année universitaire 2010/2011 Université Joseph Fourier de Grenoble - Tous droits
UE2 : Structure générale de la cellule Chapitre 7 : Structure de la cellule Le noyau cellulaire Professeur Michel SEVE Année universitaire 2010/2011 Université Joseph Fourier de Grenoble - Tous droits
Cellules procaryotes Service histologie Pr.k.mebarek
 Cellules procaryotes Service histologie Pr.k.mebarek I) Les cellules procaryotes II) Les cellules eucaryotes o 1) Caractéristiques générales des cellules eucaryotes o 2) Organisation des cellules eucaryotes
Cellules procaryotes Service histologie Pr.k.mebarek I) Les cellules procaryotes II) Les cellules eucaryotes o 1) Caractéristiques générales des cellules eucaryotes o 2) Organisation des cellules eucaryotes
LA MITOSE CUEEP - USTL DÉPARTEMENT SCIENCES BAHIJA DELATTRE
 Biologie LA MITOSE CUEEP - USTL DÉPARTEMENT SCIENCES BAHIJA DELATTRE Février 2006 I. L'INTRODUCTION Chaque cellule d'un organisme supérieur provient de la multiplication d'une cellule préexistante (cellule
Biologie LA MITOSE CUEEP - USTL DÉPARTEMENT SCIENCES BAHIJA DELATTRE Février 2006 I. L'INTRODUCTION Chaque cellule d'un organisme supérieur provient de la multiplication d'une cellule préexistante (cellule
CHAPITRE 3 LA SYNTHESE DES PROTEINES
 CHAITRE 3 LA SYNTHESE DES ROTEINES On sait qu un gène détient dans sa séquence nucléotidique, l information permettant la synthèse d un polypeptide. Ce dernier caractérisé par sa séquence d acides aminés
CHAITRE 3 LA SYNTHESE DES ROTEINES On sait qu un gène détient dans sa séquence nucléotidique, l information permettant la synthèse d un polypeptide. Ce dernier caractérisé par sa séquence d acides aminés
Dr E. CHEVRET UE2.1 2013-2014. Aperçu général sur l architecture et les fonctions cellulaires
 Aperçu général sur l architecture et les fonctions cellulaires I. Introduction II. Les microscopes 1. Le microscope optique 2. Le microscope à fluorescence 3. Le microscope confocal 4. Le microscope électronique
Aperçu général sur l architecture et les fonctions cellulaires I. Introduction II. Les microscopes 1. Le microscope optique 2. Le microscope à fluorescence 3. Le microscope confocal 4. Le microscope électronique
Annales de Biologie Cellulaire QCM (niveau SVT 1 er année)
") Annales de Biologie Cellulaire QCM (niveau SVT 1 er année) Equipe pédagogique Université Bordeaux-1 Didier Morin, Michel Moenner, Sophie North, Gérard Tramu et IJsbrand Kramer Contact : [email protected]
Annales de Biologie Cellulaire QCM (niveau SVT 1 er année) Equipe pédagogique Université Bordeaux-1 Didier Morin, Michel Moenner, Sophie North, Gérard Tramu et IJsbrand Kramer Contact : [email protected]
Le rôle de l endocytose dans les processus pathologiques
 UE7 Cours n 9 C. LAMAZE 24.11.11 Elise GODEAU (partie1) Guillaume MERGENTHALER (partie2) Le rôle de l endocytose dans les processus pathologiques SOMMAIRE : I. L endocytose à récepteurs : la voie des clathrines
UE7 Cours n 9 C. LAMAZE 24.11.11 Elise GODEAU (partie1) Guillaume MERGENTHALER (partie2) Le rôle de l endocytose dans les processus pathologiques SOMMAIRE : I. L endocytose à récepteurs : la voie des clathrines
TD de Biochimie 4 : Coloration.
 TD de Biochimie 4 : Coloration. Synthèse de l expérience 2 Les questions posées durant l expérience 2 Exposé sur les méthodes de coloration des molécules : Générique Spécifique Autres Questions Pourquoi
TD de Biochimie 4 : Coloration. Synthèse de l expérience 2 Les questions posées durant l expérience 2 Exposé sur les méthodes de coloration des molécules : Générique Spécifique Autres Questions Pourquoi
IMMUNOLOGIE. La spécificité des immunoglobulines et des récepteurs T. Informations scientifiques
 IMMUNOLOGIE La spécificité des immunoglobulines et des récepteurs T Informations scientifiques L infection par le VIH entraîne des réactions immunitaires de l organisme qui se traduisent par la production
IMMUNOLOGIE La spécificité des immunoglobulines et des récepteurs T Informations scientifiques L infection par le VIH entraîne des réactions immunitaires de l organisme qui se traduisent par la production
Travaux dirigés de Microbiologie Master I Sciences des Génomes et des Organismes Janvier 2015
 Andrew Tolonen [email protected] Travaux dirigés de Microbiologie Master I Sciences des Génomes et des Organismes Janvier 2015 A- Généralités I- La vie sur terre telle que nous la connaissons ne
Andrew Tolonen [email protected] Travaux dirigés de Microbiologie Master I Sciences des Génomes et des Organismes Janvier 2015 A- Généralités I- La vie sur terre telle que nous la connaissons ne
ATELIER SANTE PREVENTION N 2 : L ALIMENTATION
 ATELIER SANTE PREVENTION N 2 : L ALIMENTATION Mardi 24 janvier 2012 au Centre de Formation Multimétiers de REIGNAC L objectif de cet atelier sur la santé est de guider chacun vers une alimentation plus
ATELIER SANTE PREVENTION N 2 : L ALIMENTATION Mardi 24 janvier 2012 au Centre de Formation Multimétiers de REIGNAC L objectif de cet atelier sur la santé est de guider chacun vers une alimentation plus
TP N 3 La composition chimique du vivant
 Thème 1 : La Terre dans l'univers, la vie et l'évolution du vivant : une planète habitée Chapitre II : La nature du vivant TP N 3 La composition chimique du vivant Les conditions qui règnent sur terre
Thème 1 : La Terre dans l'univers, la vie et l'évolution du vivant : une planète habitée Chapitre II : La nature du vivant TP N 3 La composition chimique du vivant Les conditions qui règnent sur terre
VI- Expression du génome
 VI- Expression du génome VI-1.- EXPRESSION DU GÉNOME- PRINCIPES GÉNÉRAUX DOGME CENTRAL Les gènes et l information génétique sont conservés sous forme d acides nucléiques La perpétuation à l identique de
VI- Expression du génome VI-1.- EXPRESSION DU GÉNOME- PRINCIPES GÉNÉRAUX DOGME CENTRAL Les gènes et l information génétique sont conservés sous forme d acides nucléiques La perpétuation à l identique de
Séquence 2. L expression du patrimoine génétique. Sommaire
 Séquence 2 L expression du patrimoine génétique Sommaire 1. La synthèse des protéines 2. Phénotypes, génotypes et environnement Synthèse de la séquence 2 Exercices de la séquence 2 Glossaire des séquences
Séquence 2 L expression du patrimoine génétique Sommaire 1. La synthèse des protéines 2. Phénotypes, génotypes et environnement Synthèse de la séquence 2 Exercices de la séquence 2 Glossaire des séquences
INTRODUCTION À L'ENZYMOLOGIE
 INTRODUCTION À L'ENZYMOLOGIE Les enzymes sont des macromolécules spécialisées qui - catalysent les réactions biologiques - transforment différentes formes d'énergie. Les enzymes diffèrent des catalyseurs
INTRODUCTION À L'ENZYMOLOGIE Les enzymes sont des macromolécules spécialisées qui - catalysent les réactions biologiques - transforment différentes formes d'énergie. Les enzymes diffèrent des catalyseurs
Chapitre II La régulation de la glycémie
 Chapitre II La régulation de la glycémie Glycémie : concentration de glucose dans le sang valeur proche de 1g/L Hypoglycémie : perte de connaissance, troubles de la vue, voire coma. Hyperglycémie chronique
Chapitre II La régulation de la glycémie Glycémie : concentration de glucose dans le sang valeur proche de 1g/L Hypoglycémie : perte de connaissance, troubles de la vue, voire coma. Hyperglycémie chronique
EXERCICES : MECANISMES DE L IMMUNITE : pages 406 407 408 409 410
 EXERCICES : MECANISMES DE L IMMUNITE : pages 406 407 408 409 410 EXERCICE 1 PAGE 406 : EXPERIENCES A INTERPRETER Question : rôles respectifs du thymus et de la moelle osseuse dans la production des lymphocytes.
EXERCICES : MECANISMES DE L IMMUNITE : pages 406 407 408 409 410 EXERCICE 1 PAGE 406 : EXPERIENCES A INTERPRETER Question : rôles respectifs du thymus et de la moelle osseuse dans la production des lymphocytes.
Utilisation des substrats énergétiques
 Utilisation des substrats énergétiques Collège des Enseignants de Nutrition Date de création du document 2010-2011 Table des matières I Les organes et les substrats... 3 I.1 Les substrats énergétiques...
Utilisation des substrats énergétiques Collège des Enseignants de Nutrition Date de création du document 2010-2011 Table des matières I Les organes et les substrats... 3 I.1 Les substrats énergétiques...
- pellicule de fruits qui a un rôle de prévention contre l'évaporation, le développement de moisissures et l'infection par des parasites
 LES LIPIDES Quelles Sont les Idées Clés? Les lipides sont les huiles et les graisses de la vie courante. Ils sont insolubles dans l eau. Pour les synthétiser, une réaction : l Estérification. Pour les
LES LIPIDES Quelles Sont les Idées Clés? Les lipides sont les huiles et les graisses de la vie courante. Ils sont insolubles dans l eau. Pour les synthétiser, une réaction : l Estérification. Pour les
Déshydratation, Inclusions, Coupes, et microscopie électronique à transmission
 Déshydratation, Inclusions, Coupes, et microscopie électronique à transmission Valérie Messent Lobservation de tissus vivants en microscopie électronique à transmission impose des techniques de préparations
Déshydratation, Inclusions, Coupes, et microscopie électronique à transmission Valérie Messent Lobservation de tissus vivants en microscopie électronique à transmission impose des techniques de préparations
Information génétique
 chapitre 3 Information génétique et division cellulaire L étude de la division cellulaire est abordée pour découvrir comment est transmise et conservée l information génétique portée par les chromosomes.
chapitre 3 Information génétique et division cellulaire L étude de la division cellulaire est abordée pour découvrir comment est transmise et conservée l information génétique portée par les chromosomes.
Le trajet des aliments dans l appareil digestif.
 La digestion. La digestion, c est la transformation des aliments en nutriments assimilables par l organisme. Dans le tube digestif, les aliments subissent une série de dégradations mécaniques et chimiques
La digestion. La digestion, c est la transformation des aliments en nutriments assimilables par l organisme. Dans le tube digestif, les aliments subissent une série de dégradations mécaniques et chimiques
Formavie 2010. 2 Différentes versions du format PDB...3. 3 Les champs dans les fichiers PDB...4. 4 Le champ «ATOM»...5. 6 Limites du format PDB...
 Formavie 2010 Les fichiers PDB Les fichiers PDB contiennent les informations qui vont permettre à des logiciels de visualisation moléculaire (ex : RasTop ou Jmol) d afficher les molécules. Un fichier au
Formavie 2010 Les fichiers PDB Les fichiers PDB contiennent les informations qui vont permettre à des logiciels de visualisation moléculaire (ex : RasTop ou Jmol) d afficher les molécules. Un fichier au
Interactions des rayonnements avec la matière
 UE3-1 : Biophysique Chapitre 2 : Interactions des rayonnements avec la matière Professeur Jean-Philippe VUILLEZ Année universitaire 2011/2012 Université Joseph Fourier de Grenoble - Tous droits réservés.
UE3-1 : Biophysique Chapitre 2 : Interactions des rayonnements avec la matière Professeur Jean-Philippe VUILLEZ Année universitaire 2011/2012 Université Joseph Fourier de Grenoble - Tous droits réservés.
www.gbo.com/bioscience 1 Culture Cellulaire Microplaques 2 HTS- 3 Immunologie/ HLA 4 Microbiologie/ Bactériologie Containers 5 Tubes/ 6 Pipetage
 2 HTS 3 Immunologie / Immunologie Informations Techniques 3 I 2 ELISA 96 Puits 3 I 4 ELISA 96 Puits en Barrettes 3 I 6 en Barrettes de 8 Puits 3 I 7 en Barrettes de 12 Puits 3 I 8 en Barrettes de 16 Puits
2 HTS 3 Immunologie / Immunologie Informations Techniques 3 I 2 ELISA 96 Puits 3 I 4 ELISA 96 Puits en Barrettes 3 I 6 en Barrettes de 8 Puits 3 I 7 en Barrettes de 12 Puits 3 I 8 en Barrettes de 16 Puits
Table des matières Introduction Chapitre*1*:*De*la*matière*sérieuse Chapitre*2*:*Clair*comme*de*l eau*de*roche
 Table des matières Introduction! *! Chapitre*1*:*De*la*matière*sérieuse! 1.1!Les!matériaux! 1.2!Solides,!liquides!et!gaz 1.3!Gaspiller!de!l énergie! 1.4!Le!carburant!du!corps! Chapitre*2*:*Clair*comme*de*l
Table des matières Introduction! *! Chapitre*1*:*De*la*matière*sérieuse! 1.1!Les!matériaux! 1.2!Solides,!liquides!et!gaz 1.3!Gaspiller!de!l énergie! 1.4!Le!carburant!du!corps! Chapitre*2*:*Clair*comme*de*l
Transport des gaz dans le sang
 UE3-2 - Physiologie Physiologie Respiratoire Chapitre 9 : Transport des gaz dans le sang Docteur Sandrine LAUNOIS-ROLLINAT Année universitaire 2011/2012 Université Joseph Fourier de Grenoble - Tous droits
UE3-2 - Physiologie Physiologie Respiratoire Chapitre 9 : Transport des gaz dans le sang Docteur Sandrine LAUNOIS-ROLLINAT Année universitaire 2011/2012 Université Joseph Fourier de Grenoble - Tous droits
Séquence 1. Reproduction conforme de la cellule et réplication de l ADN Variabilité génétique et mutation de l ADN
 Séquence 1 Reproduction conforme de la cellule et réplication de l ADN Variabilité génétique et mutation de l ADN Sommaire 1. Reproduction conforme de la cellule et réplication de l ADN 2. Variabilité
Séquence 1 Reproduction conforme de la cellule et réplication de l ADN Variabilité génétique et mutation de l ADN Sommaire 1. Reproduction conforme de la cellule et réplication de l ADN 2. Variabilité
AGRÉGATION DE SCIENCES DE LA VIE - SCIENCES DE LA TERRE ET DE L UNIVERS
 AGRÉGATION DE SCIENCES DE LA VIE - SCIENCES DE LA TERRE ET DE L UNIVERS CONCOURS EXTERNE ÉPREUVES D ADMISSION session 2010 TRAVAUX PRATIQUES DE CONTRE-OPTION DU SECTEUR A CANDIDATS DES SECTEURS B ET C
AGRÉGATION DE SCIENCES DE LA VIE - SCIENCES DE LA TERRE ET DE L UNIVERS CONCOURS EXTERNE ÉPREUVES D ADMISSION session 2010 TRAVAUX PRATIQUES DE CONTRE-OPTION DU SECTEUR A CANDIDATS DES SECTEURS B ET C
ÉCOLES NORMALES SUPÉRIEURES ÉCOLE NATIONALE DES PONTS ET CHAUSSÉES CONCOURS D ADMISSION SESSION 2013 FILIÈRE BCPST COMPOSITION DE BIOLOGIE
 ÉCOLES NORMALES SUPÉRIEURES ÉCOLE NATIONALE DES PONTS ET CHAUSSÉES CONCOURS D ADMISSION SESSION 2013 FILIÈRE BCPST COMPOSITION DE BIOLOGIE Épreuve commune aux ENS de Cachan, Lyon, Paris et de l ENPC Durée
ÉCOLES NORMALES SUPÉRIEURES ÉCOLE NATIONALE DES PONTS ET CHAUSSÉES CONCOURS D ADMISSION SESSION 2013 FILIÈRE BCPST COMPOSITION DE BIOLOGIE Épreuve commune aux ENS de Cachan, Lyon, Paris et de l ENPC Durée
Commentaires sur les épreuves de Sciences de la Vie et de la Terre
 Commentaires sur les épreuves de Sciences de la Vie et de la Terre Épreuve ÉCRITE de BIOLOGIE A...2 Épreuve ÉCRITE de BIOLOGIE B...9 Épreuve ORALE de BIOLOGIE...13 ANNEXE 1 : LISTE DES SUJETS D ORAL PROPOSÉS
Commentaires sur les épreuves de Sciences de la Vie et de la Terre Épreuve ÉCRITE de BIOLOGIE A...2 Épreuve ÉCRITE de BIOLOGIE B...9 Épreuve ORALE de BIOLOGIE...13 ANNEXE 1 : LISTE DES SUJETS D ORAL PROPOSÉS
5.5.5 Exemple d un essai immunologique
 5.5.5 Exemple d un essai immunologique Test de grossesse Test en forme de bâtonnet destiné à mettre en évidence l'hormone spécifique de la grossesse, la gonadotrophine chorionique humaine (hcg), une glycoprotéine.
5.5.5 Exemple d un essai immunologique Test de grossesse Test en forme de bâtonnet destiné à mettre en évidence l'hormone spécifique de la grossesse, la gonadotrophine chorionique humaine (hcg), une glycoprotéine.
Module 5 La maturation de l ARN et le contrôle post-transcriptionnel chez les eucaryotes
 Module 5 La maturation de l ARN et le contrôle post-transcriptionnel chez les eucaryotes Où trouver l'information complémentaire? MCB -11, GVII-5, 22, 23. La maturation des ARNm chez les eucaryotes Les
Module 5 La maturation de l ARN et le contrôle post-transcriptionnel chez les eucaryotes Où trouver l'information complémentaire? MCB -11, GVII-5, 22, 23. La maturation des ARNm chez les eucaryotes Les
NEUROPHYSIOLOGIE (2)
") NEUROPHYSIOLOGIE (2) B - LE SYSTEME NERVEUX (SUITE) - NEURONES ET GLIOCYTES IV - LES NEURONES ET LES GLIOCYTES 1) OBJET DE LEUR ETUDE La NEUROPHYSIOLOGIE est l étude du fonctionnement du système nerveux.
NEUROPHYSIOLOGIE (2) B - LE SYSTEME NERVEUX (SUITE) - NEURONES ET GLIOCYTES IV - LES NEURONES ET LES GLIOCYTES 1) OBJET DE LEUR ETUDE La NEUROPHYSIOLOGIE est l étude du fonctionnement du système nerveux.
grande simple microscope microscope inventé années 1825. biologie = cellule) et (logos de plus en Anglais. Utilise un La microscopie, 1665,
 et (logos de plus en Anglais. Utilise un La microscopie, 1665,") Cours de Biologie Cellulaire Présentés par Mr CHELLI A. FSNV 2012/ /2013 CHAPITRE I : INTRODUCTION A LA BIOLOGIE CELLULAIRE A- Introduction et définitionn de la biologie cellulaire : Il était difficile
Cours de Biologie Cellulaire Présentés par Mr CHELLI A. FSNV 2012/ /2013 CHAPITRE I : INTRODUCTION A LA BIOLOGIE CELLULAIRE A- Introduction et définitionn de la biologie cellulaire : Il était difficile
Transport des gaz dans le sang
 UE3-2 - Physiologie Physiologie Respiratoire Chapitre 9 : Transport des gaz dans le sang Docteur Sandrine LAUNOIS-ROLLINAT Année universitaire 2010/2011 Université Joseph Fourier de Grenoble - Tous droits
UE3-2 - Physiologie Physiologie Respiratoire Chapitre 9 : Transport des gaz dans le sang Docteur Sandrine LAUNOIS-ROLLINAT Année universitaire 2010/2011 Université Joseph Fourier de Grenoble - Tous droits
STRUCTURE ET FONCTION DES PLURICELLULAIRES
 Plan de cours STRUCTURE ET FONCTION DES PLURICELLULAIRES 101-FYA-PT Pondération 3-1-2 Gilles Bourbonnais (C360) [email protected] Sciences de la Nature / PASC@L http://ici.cegep-ste-foy.qc.ca/profs/gbourbonnais/
Plan de cours STRUCTURE ET FONCTION DES PLURICELLULAIRES 101-FYA-PT Pondération 3-1-2 Gilles Bourbonnais (C360) [email protected] Sciences de la Nature / PASC@L http://ici.cegep-ste-foy.qc.ca/profs/gbourbonnais/
Contrôle de l'expression génétique : Les régulations post-transcriptionnelles
 Contrôle de l'expression génétique : Les régulations post-transcriptionnelles http://perso.univ-rennes1.fr/serge.hardy/ utilisateur : biochimie mot de passe : 2007 L'ARNm, simple intermédiaire entre le
Contrôle de l'expression génétique : Les régulations post-transcriptionnelles http://perso.univ-rennes1.fr/serge.hardy/ utilisateur : biochimie mot de passe : 2007 L'ARNm, simple intermédiaire entre le
2 C est quoi la chimie?
 PARTIE 1 AVANT LA CHIMIE VERTE... 2 C est quoi la chimie? L inconnu étant source d angoisse, nous allons essayer de définir les grands domaines de la chimie pour mieux la connaître, l appréhender et donc
PARTIE 1 AVANT LA CHIMIE VERTE... 2 C est quoi la chimie? L inconnu étant source d angoisse, nous allons essayer de définir les grands domaines de la chimie pour mieux la connaître, l appréhender et donc
AGREGATION DE BIOCHIMIE GENIE BIOLOGIQUE
 AGREGATION DE BIOCHIMIE GENIE BIOLOGIQUE CONCOURS EXTERNE Session 2005 TRAVAUX PRATIQUES DE BIOCHIMIE PHYSIOLOGIE ALCOOL ET FOIE L éthanol, psychotrope puissant, est absorbé passivement dans l intestin
AGREGATION DE BIOCHIMIE GENIE BIOLOGIQUE CONCOURS EXTERNE Session 2005 TRAVAUX PRATIQUES DE BIOCHIMIE PHYSIOLOGIE ALCOOL ET FOIE L éthanol, psychotrope puissant, est absorbé passivement dans l intestin
Structure quantique cohérente et incohérente de l eau liquide
 Structure quantique cohérente et incohérente de l eau liquide Prof. Marc HENRY Chimie Moléculaire du Solide Institut Le Bel, 4, Rue Blaise Pascal 67070 Strasbourg Cedex, France Tél: 03.68.85.15.00 e-mail:
Structure quantique cohérente et incohérente de l eau liquide Prof. Marc HENRY Chimie Moléculaire du Solide Institut Le Bel, 4, Rue Blaise Pascal 67070 Strasbourg Cedex, France Tél: 03.68.85.15.00 e-mail:
RDP : Voir ou conduire
 1S Thème : Observer RDP : Voir ou conduire DESCRIPTIF DE SUJET DESTINE AU PROFESSEUR Objectif Compétences exigibles du B.O. Initier les élèves de première S à la démarche de résolution de problème telle
1S Thème : Observer RDP : Voir ou conduire DESCRIPTIF DE SUJET DESTINE AU PROFESSEUR Objectif Compétences exigibles du B.O. Initier les élèves de première S à la démarche de résolution de problème telle
A B C Eau Eau savonneuse Eau + détergent
 1L : Physique et chimie dans la cuisine Chapitre.3 : Chimie et lavage I. Les savons et les détergents synthétiques 1. Propriétés détergentes des savons Le savon est un détergent naturel, les détergents
1L : Physique et chimie dans la cuisine Chapitre.3 : Chimie et lavage I. Les savons et les détergents synthétiques 1. Propriétés détergentes des savons Le savon est un détergent naturel, les détergents
La reconnaissance moléculaire: la base du design rationnel Modélisation moléculaire: Introduction Hiver 2006
 La reconnaissance moléculaire: la base du design rationnel En 1890 Emil Fisher a proposé le modèle "serrure et clé" pour expliquer la façon de fonctionner des systèmes biologiques. Un substrat rentre et
La reconnaissance moléculaire: la base du design rationnel En 1890 Emil Fisher a proposé le modèle "serrure et clé" pour expliquer la façon de fonctionner des systèmes biologiques. Un substrat rentre et
PHYSIQUE-CHIMIE DANS LA CUISINE Chapitre 3 : Chimie et lavage
 PHYSIQUE-CHIMIE DANS LA CUISINE Chapitre 3 : Chimie et lavage I) Qu'est-ce qu'un savon et comment le fabrique-t-on? D'après épreuve BAC Liban 2005 Physique-Chimie dans la cuisine Chapitre 3 1/6 1- En vous
PHYSIQUE-CHIMIE DANS LA CUISINE Chapitre 3 : Chimie et lavage I) Qu'est-ce qu'un savon et comment le fabrique-t-on? D'après épreuve BAC Liban 2005 Physique-Chimie dans la cuisine Chapitre 3 1/6 1- En vous
Biochimie I. Extraction et quantification de l hexokinase dans Saccharomyces cerevisiae 1. Assistants : Tatjana Schwabe Marcy Taylor Gisèle Dewhurst
 Biochimie I Extraction et quantification de l hexokinase dans Saccharomyces cerevisiae 1 Daniel Abegg Sarah Bayat Alexandra Belfanti Assistants : Tatjana Schwabe Marcy Taylor Gisèle Dewhurst Laboratoire
Biochimie I Extraction et quantification de l hexokinase dans Saccharomyces cerevisiae 1 Daniel Abegg Sarah Bayat Alexandra Belfanti Assistants : Tatjana Schwabe Marcy Taylor Gisèle Dewhurst Laboratoire
Séquence 4. La nature du vivant. Sommaire. 1. L unité structurale et chimique du vivant. 2. L ADN, support de l information génétique
 Séquence 4 La nature du vivant Sommaire 1. L unité structurale et chimique du vivant 2. L ADN, support de l information génétique 3. Synthèse de la séquence 4. Exercices Devoir autocorrectif n 2 Séquence
Séquence 4 La nature du vivant Sommaire 1. L unité structurale et chimique du vivant 2. L ADN, support de l information génétique 3. Synthèse de la séquence 4. Exercices Devoir autocorrectif n 2 Séquence
Lycée Galilée Gennevilliers. chap. 6. JALLU Laurent. I. Introduction... 2 La source d énergie nucléaire... 2
 Lycée Galilée Gennevilliers L'énergie nucléaire : fusion et fission chap. 6 JALLU Laurent I. Introduction... 2 La source d énergie nucléaire... 2 II. Équivalence masse-énergie... 3 Bilan de masse de la
Lycée Galilée Gennevilliers L'énergie nucléaire : fusion et fission chap. 6 JALLU Laurent I. Introduction... 2 La source d énergie nucléaire... 2 II. Équivalence masse-énergie... 3 Bilan de masse de la
Compétence 3-1 S EXPRIMER A L ECRIT Fiche professeur
 Compétence 3-1 S EXPRIMER A L ECRIT Fiche professeur Nature de l activité : Réaliser 3 types de productions écrites (réécriture de notes, production d une synthèse de documents, production d une argumentation)
Compétence 3-1 S EXPRIMER A L ECRIT Fiche professeur Nature de l activité : Réaliser 3 types de productions écrites (réécriture de notes, production d une synthèse de documents, production d une argumentation)
I. La levure Saccharomyces cerevisiae: mode de vie
 LES LEVURES UE «levures» -5 avril: généralités (MN Simon) -6 avril: analyse génétique (MN Simon) -6 avril: Cycle cellulaire I: la réplication (E. bailly) -7 avril: Cycle cellulaire II: la mitose (E. Bailly)
LES LEVURES UE «levures» -5 avril: généralités (MN Simon) -6 avril: analyse génétique (MN Simon) -6 avril: Cycle cellulaire I: la réplication (E. bailly) -7 avril: Cycle cellulaire II: la mitose (E. Bailly)
Effets électroniques-acidité/basicité
 Université du Maine Faculté des Sciences Retour Révisions de cours Effets électroniquesacidité/basicité Il est très important dans un cours de himie organique de connaitre ces notions qui vont intervenir
Université du Maine Faculté des Sciences Retour Révisions de cours Effets électroniquesacidité/basicité Il est très important dans un cours de himie organique de connaitre ces notions qui vont intervenir
Chapitre 6. Réactions nucléaires. 6.1 Généralités. 6.1.1 Définitions. 6.1.2 Lois de conservation
 Chapitre 6 Réactions nucléaires 6.1 Généralités 6.1.1 Définitions Un atome est constitué d électrons et d un noyau, lui-même constitué de nucléons (protons et neutrons). Le nombre de masse, noté, est le
Chapitre 6 Réactions nucléaires 6.1 Généralités 6.1.1 Définitions Un atome est constitué d électrons et d un noyau, lui-même constitué de nucléons (protons et neutrons). Le nombre de masse, noté, est le
3: Clonage d un gène dans un plasmide
 3: Clonage d un gène dans un plasmide Le clonage moléculaire est une des bases du génie génétique. Il consiste à insérer un fragment d'adn (dénommé insert) dans un vecteur approprié comme un plasmide par
3: Clonage d un gène dans un plasmide Le clonage moléculaire est une des bases du génie génétique. Il consiste à insérer un fragment d'adn (dénommé insert) dans un vecteur approprié comme un plasmide par
Les cytokines et leurs récepteurs. Laurence Guglielmi [email protected]
 Les cytokines et leurs récepteurs Laurence Guglielmi [email protected] l i@ i 1 Les cytokines et leurs récepteurs 2 mécanismes principaux d interactions cellulaires : - contact membranaire
Les cytokines et leurs récepteurs Laurence Guglielmi [email protected] l i@ i 1 Les cytokines et leurs récepteurs 2 mécanismes principaux d interactions cellulaires : - contact membranaire
DIFFRACTion des ondes
 DIFFRACTion des ondes I DIFFRACTION DES ONDES PAR LA CUVE À ONDES Lorsqu'une onde plane traverse un trou, elle se transforme en onde circulaire. On dit que l'onde plane est diffractée par le trou. Ce phénomène
DIFFRACTion des ondes I DIFFRACTION DES ONDES PAR LA CUVE À ONDES Lorsqu'une onde plane traverse un trou, elle se transforme en onde circulaire. On dit que l'onde plane est diffractée par le trou. Ce phénomène
AMINES BIOGENIQUES. Dopamine/Noradrénaline/Adrénaline (CATECHOLAMINES) Sérotonine/Histamine/Dopamine
 Sérotonine/Histamine/Dopamine") AMINES BIOGENIQUES Dopamine/Noradrénaline/Adrénaline (CATECHOLAMINES) Sérotonine/Histamine/Dopamine Effets physiologiques des bioamines via le systeme nerveux autonome Cardiovasculaire: Augmentation du
AMINES BIOGENIQUES Dopamine/Noradrénaline/Adrénaline (CATECHOLAMINES) Sérotonine/Histamine/Dopamine Effets physiologiques des bioamines via le systeme nerveux autonome Cardiovasculaire: Augmentation du
COUSIN Fabien KERGOURLAY Gilles. 19 octobre 2007. de l hôte par les. Master 2 MFA Responsable : UE Incidence des paramètres environnementaux
 COUSIN Fabien KERGOURLAY Gilles 19 octobre 2007 Inhibition des défenses de l hôte par les bactéries pathogènes Master 2 MFA Responsable : UE Incidence des paramètres environnementaux Gwennola ERMEL I Détection
COUSIN Fabien KERGOURLAY Gilles 19 octobre 2007 Inhibition des défenses de l hôte par les bactéries pathogènes Master 2 MFA Responsable : UE Incidence des paramètres environnementaux Gwennola ERMEL I Détection
Epreuve de biologie... 2 Annexe : Liste des sujets de la session 2013... 9
 Epreuve de biologie... 2 Annexe : Liste des sujets de la session 2013... 9 Travaux pratiques de biologie... 17 Annexe : Liste des sujets de la session 2013... 26 Travaux d initiative personnelle encadrés
Epreuve de biologie... 2 Annexe : Liste des sujets de la session 2013... 9 Travaux pratiques de biologie... 17 Annexe : Liste des sujets de la session 2013... 26 Travaux d initiative personnelle encadrés
Critères pour les méthodes de quantification des résidus potentiellement allergéniques de protéines de collage dans le vin (OIV-Oeno 427-2010)
") Méthode OIV- -MA-AS315-23 Type de méthode : critères Critères pour les méthodes de quantification des résidus potentiellement allergéniques de protéines de collage (OIV-Oeno 427-2010) 1 Définitions des
Méthode OIV- -MA-AS315-23 Type de méthode : critères Critères pour les méthodes de quantification des résidus potentiellement allergéniques de protéines de collage (OIV-Oeno 427-2010) 1 Définitions des
Les OGM. 5 décembre 2008. Nicole Mounier
 Les OGM 5 décembre 2008 Nicole Mounier Université Claude Bernard Lyon 1 CGMC, bâtiment Gregor Mendel 43, boulevard du 11 Novembre 1918 69622 Villeurbanne Cedex OGM Organismes Génétiquement Modifiés Transfert
Les OGM 5 décembre 2008 Nicole Mounier Université Claude Bernard Lyon 1 CGMC, bâtiment Gregor Mendel 43, boulevard du 11 Novembre 1918 69622 Villeurbanne Cedex OGM Organismes Génétiquement Modifiés Transfert
University of Tokyo Graduate School of Agricultural and Life Sciences et. Kanagawa Academy of Science and Technology
 COMMUNIQUÉ DE PRESSE le 25 mars 2014 Une étude conjointe menée par University of Tokyo Graduate School of Agricultural and Life Sciences et Kanagawa Academy of Science and Technology suggère que le sirop
COMMUNIQUÉ DE PRESSE le 25 mars 2014 Une étude conjointe menée par University of Tokyo Graduate School of Agricultural and Life Sciences et Kanagawa Academy of Science and Technology suggère que le sirop
Rôle des acides biliaires dans la régulation de l homéostasie du glucose : implication de FXR dans la cellule bêta-pancréatique
 Rôle des acides biliaires dans la régulation de l homéostasie du glucose : implication de FXR dans la cellule bêta-pancréatique Tuteur : Anne Muhr-Tailleux cardiovasculaires et diabète (Equipe 1) Institut
Rôle des acides biliaires dans la régulation de l homéostasie du glucose : implication de FXR dans la cellule bêta-pancréatique Tuteur : Anne Muhr-Tailleux cardiovasculaires et diabète (Equipe 1) Institut
Biologie Appliquée. Dosages Immunologiques TD9 Mai 2015. Stéphanie Sigaut INSERM U1141 [email protected]
 Biologie Appliquée Dosages Immunologiques TD9 Mai 2015 Stéphanie Sigaut INSERM U1141 [email protected] 1 ELISA 2 3 4 [Ac] 5 6 7 8 9 Correction : Faire la moyenne D0-1 et D0-2 pour toute les valeurs
Biologie Appliquée Dosages Immunologiques TD9 Mai 2015 Stéphanie Sigaut INSERM U1141 [email protected] 1 ELISA 2 3 4 [Ac] 5 6 7 8 9 Correction : Faire la moyenne D0-1 et D0-2 pour toute les valeurs
POLITIQUE DE BIOSÉCURITÉ
 Date d entrée en vigueur: Mai 2006 Remplace/amende: VRS-52/s/o Origine: Vice-rectorat aux services Numéro de référence: VPS-52 DÉFINITION Une substance biologique dangereuse se définit comme un organisme
Date d entrée en vigueur: Mai 2006 Remplace/amende: VRS-52/s/o Origine: Vice-rectorat aux services Numéro de référence: VPS-52 DÉFINITION Une substance biologique dangereuse se définit comme un organisme
La filtration glomérulaire et sa régulation
 UE3-2 - Physiologie rénale Chapitre 4 : La filtration glomérulaire et sa régulation Professeur Diane GODIN-RIBUOT Année universitaire 2010/2011 Université Joseph Fourier de Grenoble - Tous droits réservés.
UE3-2 - Physiologie rénale Chapitre 4 : La filtration glomérulaire et sa régulation Professeur Diane GODIN-RIBUOT Année universitaire 2010/2011 Université Joseph Fourier de Grenoble - Tous droits réservés.
Tableau récapitulatif : composition nutritionnelle de la spiruline
 Tableau récapitulatif : composition nutritionnelle de la spiruline (Valeur énergétique : 38 kcal/10 g) Composition nutritionnelle Composition pour 10 g Rôle Protéines (végétales) 55 à 70 % Construction
Tableau récapitulatif : composition nutritionnelle de la spiruline (Valeur énergétique : 38 kcal/10 g) Composition nutritionnelle Composition pour 10 g Rôle Protéines (végétales) 55 à 70 % Construction
Banque Agro-Veto Session 2014 Rapport sur les concours A filière BCPST
 1 Épreuve de biologie Épreuve non prise en compte au concours PC BIO Concours Nb cand. Moyenne Ecart type Note la plus basse Note la plus haute A BIO 2158 11,94 4,10 1 20 A ENV 883 13,54 3,69 1,5 20 Ce
1 Épreuve de biologie Épreuve non prise en compte au concours PC BIO Concours Nb cand. Moyenne Ecart type Note la plus basse Note la plus haute A BIO 2158 11,94 4,10 1 20 A ENV 883 13,54 3,69 1,5 20 Ce
ACIDES BASES. Chap.5 SPIESS
 ACIDES BASES «Je ne crois pas que l on me conteste que l acide n ait des pointes Il ne faut que le goûter pour tomber dans ce sentiment car il fait des picotements sur la langue.» Notion d activité et
ACIDES BASES «Je ne crois pas que l on me conteste que l acide n ait des pointes Il ne faut que le goûter pour tomber dans ce sentiment car il fait des picotements sur la langue.» Notion d activité et
FUSION PAR CONFINEMENT MAGNÉTIQUE
 FUSION PAR CONFINEMENT MAGNÉTIQUE Séminaire de Xavier GARBET pour le FIP 06/01/2009 Anthony Perret Michel Woné «La production d'énergie par fusion thermonucléaire contrôlée est un des grands défis scientifiques
FUSION PAR CONFINEMENT MAGNÉTIQUE Séminaire de Xavier GARBET pour le FIP 06/01/2009 Anthony Perret Michel Woné «La production d'énergie par fusion thermonucléaire contrôlée est un des grands défis scientifiques
Partie Observer : Ondes et matière CHAP 04-ACT/DOC Analyse spectrale : Spectroscopies IR et RMN
 Partie Observer : Ondes et matière CHAP 04-ACT/DOC Analyse spectrale : Spectroscopies IR et RMN Objectifs : Exploiter un spectre infrarouge pour déterminer des groupes caractéristiques Relier un spectre
Partie Observer : Ondes et matière CHAP 04-ACT/DOC Analyse spectrale : Spectroscopies IR et RMN Objectifs : Exploiter un spectre infrarouge pour déterminer des groupes caractéristiques Relier un spectre
Synthèse et propriétés des savons.
 Synthèse et propriétés des savons. Objectifs: Réaliser la synthèse d'un savon mise en évidence de quelques propriétés des savons. I Introduction: 1. Présentation des savons: a) Composition des savons.
Synthèse et propriétés des savons. Objectifs: Réaliser la synthèse d'un savon mise en évidence de quelques propriétés des savons. I Introduction: 1. Présentation des savons: a) Composition des savons.
Univers Vivant Révision. Notions STE
 Univers Vivant Révision Notions STE Chap. 13) L Écologie 1) a) Qu est-ce que l empreinte écologique? L empreinte écologique correspond à la surface terrestre et aquatique totale nécessaire à un individu,
Univers Vivant Révision Notions STE Chap. 13) L Écologie 1) a) Qu est-ce que l empreinte écologique? L empreinte écologique correspond à la surface terrestre et aquatique totale nécessaire à un individu,
Microscopie Confocale. Principes de base & Applications en Biologie Cellulaire
 Université Paris Descartes L3 - Licence Professionnelle «Industries chimiques et Pharmaceutiques Option Biotechnologie» Microscopie Confocale Principes de base & Applications en Biologie Cellulaire Bruno
Université Paris Descartes L3 - Licence Professionnelle «Industries chimiques et Pharmaceutiques Option Biotechnologie» Microscopie Confocale Principes de base & Applications en Biologie Cellulaire Bruno
La séparation membranaire : comment maintenir la performance des membranes?
 La séparation membranaire : comment maintenir la performance des membranes? Alfa Arzate, ing., Ph.D. Journées Acéricoles Hiver 2010 OBJECTIF DE LA PRÉSENTATION L objectif premier de cette présentation
La séparation membranaire : comment maintenir la performance des membranes? Alfa Arzate, ing., Ph.D. Journées Acéricoles Hiver 2010 OBJECTIF DE LA PRÉSENTATION L objectif premier de cette présentation
Liquides oraux : et suspensions. Préparations liquides pour usage oral. Solutions
 Préparations pharmaceutique Cours de en 2ème petites Année quantités de Master en Pharmacie Liquides oraux : solutions, Préparation sirops pharmaceutique et suspensions en petites quantités Section des
Préparations pharmaceutique Cours de en 2ème petites Année quantités de Master en Pharmacie Liquides oraux : solutions, Préparation sirops pharmaceutique et suspensions en petites quantités Section des
101 Adoptée : 12 mai 1981
 LIGNE DIRECTRICE DE L OCDE POUR LES ESSAIS DE PRODUITS CHIMIQUES 101 Adoptée : 12 mai 1981 «Spectres d'absorption UV-VIS» (Méthode spectrophotométrique) 1. I N T R O D U C T I O N I n f o r m a t i o n
LIGNE DIRECTRICE DE L OCDE POUR LES ESSAIS DE PRODUITS CHIMIQUES 101 Adoptée : 12 mai 1981 «Spectres d'absorption UV-VIS» (Méthode spectrophotométrique) 1. I N T R O D U C T I O N I n f o r m a t i o n
ANTICORPS POLYCLONAUX ANTI IMMUNOGLOBULINES
 L OUTIL IDEAL POUR TOUTES LES DETECTIONS IMMUNOCHIMIQUES pour toutes les techniques immunodosages (EIA/ELISA) dot/ westernblot immunohistochimie immunocytochimie cytométrie en flux quel que soit le système
L OUTIL IDEAL POUR TOUTES LES DETECTIONS IMMUNOCHIMIQUES pour toutes les techniques immunodosages (EIA/ELISA) dot/ westernblot immunohistochimie immunocytochimie cytométrie en flux quel que soit le système
Ebauche Rapport finale
 Ebauche Rapport finale Sommaire : 1 - Introduction au C.D.N. 2 - Définition de la problématique 3 - Etat de l'art : Présentatio de 3 Topologies streaming p2p 1) INTRODUCTION au C.D.N. La croissance rapide
Ebauche Rapport finale Sommaire : 1 - Introduction au C.D.N. 2 - Définition de la problématique 3 - Etat de l'art : Présentatio de 3 Topologies streaming p2p 1) INTRODUCTION au C.D.N. La croissance rapide
Nous vous présentons la technologie du traitement de surfaces du 21 ème siècle
 Nous vous présentons la technologie du traitement de surfaces du 21 ème siècle 2009 NADICO Ltd. Germany www.nadico.de Le revêtement Titan Effect TE1022 améliorer la rentabilité des installations solaires
Nous vous présentons la technologie du traitement de surfaces du 21 ème siècle 2009 NADICO Ltd. Germany www.nadico.de Le revêtement Titan Effect TE1022 améliorer la rentabilité des installations solaires
Chapitre 02. La lumière des étoiles. Exercices :
 Chapitre 02 La lumière des étoiles. I- Lumière monochromatique et lumière polychromatique. )- Expérience de Newton (642 727). 2)- Expérience avec la lumière émise par un Laser. 3)- Radiation et longueur
Chapitre 02 La lumière des étoiles. I- Lumière monochromatique et lumière polychromatique. )- Expérience de Newton (642 727). 2)- Expérience avec la lumière émise par un Laser. 3)- Radiation et longueur
Les composites thermoplastiques
 Les composites thermoplastiques Définition Par définition, un thermoplastique (anglais :thermoplast) est un matériau à base de polymère (composé de macromolécules) qui peut être mis en forme, à l état
Les composites thermoplastiques Définition Par définition, un thermoplastique (anglais :thermoplast) est un matériau à base de polymère (composé de macromolécules) qui peut être mis en forme, à l état
Section «Maturité fédérale» EXAMENS D'ADMISSION Session de février 2014 RÉCAPITULATIFS DES MATIÈRES EXAMINÉES. Formation visée
 EXAMENS D'ADMISSION Admission RÉCAPITULATIFS DES MATIÈRES EXAMINÉES MATIÈRES Préparation en 3 ou 4 semestres Formation visée Préparation complète en 1 an 2 ème partiel (semestriel) Niveau Durée de l examen
EXAMENS D'ADMISSION Admission RÉCAPITULATIFS DES MATIÈRES EXAMINÉES MATIÈRES Préparation en 3 ou 4 semestres Formation visée Préparation complète en 1 an 2 ème partiel (semestriel) Niveau Durée de l examen
(72) Inventeur: Baijot, Bruno Faculté des Se. Agronom. de l'etat Dép. de Technol. agro-alimentaire et forestière groupe Ceteder B5800 Gembloux(BE)
 Inventeur: Baijot, Bruno Faculté des Se. Agronom. de l'etat Dép. de Technol. agro-alimentaire et forestière groupe Ceteder B5800 Gembloux(BE)") Patentamt à JEuropâisches European Patent Office Numéro de publication: 0 039 1 04 Office européen des brevets A1 DEMANDE DE BREVET EUROPEEN Numéro de dépôt: 81200405.9 Int. Cl.3: C 12 N 11/00 Date de
Patentamt à JEuropâisches European Patent Office Numéro de publication: 0 039 1 04 Office européen des brevets A1 DEMANDE DE BREVET EUROPEEN Numéro de dépôt: 81200405.9 Int. Cl.3: C 12 N 11/00 Date de
Stockage de chaleur solaire par sorption : Analyse et contrôle du système à partir de sa simulation dynamique
 Stockage de chaleur solaire par sorption : Analyse et contrôle du système à partir de sa simulation dynamique Kokouvi Edem N TSOUKPOE 1, Nolwenn LE PIERRÈS 1*, Lingai LUO 1 1 LOCIE, CNRS FRE3220-Université
Stockage de chaleur solaire par sorption : Analyse et contrôle du système à partir de sa simulation dynamique Kokouvi Edem N TSOUKPOE 1, Nolwenn LE PIERRÈS 1*, Lingai LUO 1 1 LOCIE, CNRS FRE3220-Université
Molécules et Liaison chimique
 Molécules et liaison chimique Molécules et Liaison chimique La liaison dans La liaison dans Le point de vue classique: l approche l de deux atomes d hydrogd hydrogènes R -0,9-1 0 0,5 1 1,5,5 3 3,5 4 R
Molécules et liaison chimique Molécules et Liaison chimique La liaison dans La liaison dans Le point de vue classique: l approche l de deux atomes d hydrogd hydrogènes R -0,9-1 0 0,5 1 1,5,5 3 3,5 4 R
Chapitre : METHODES D'ETUDE DE LA CELLULE
 Chapitre : METHODES D'ETUDE DE LA CELLULE 1ère PARTIE : METHODES D'ETUDE DE LA MORPHOLOGIE DES CELLULES La plupart des méthodes sont basées sur l'emploi des microscopes, la taille des cellules et de leurs
Chapitre : METHODES D'ETUDE DE LA CELLULE 1ère PARTIE : METHODES D'ETUDE DE LA MORPHOLOGIE DES CELLULES La plupart des méthodes sont basées sur l'emploi des microscopes, la taille des cellules et de leurs
Chapitre V : La gestion de la mémoire. Hiérarchie de mémoires Objectifs Méthodes d'allocation Simulation de mémoire virtuelle Le mapping
 Chapitre V : La gestion de la mémoire Hiérarchie de mémoires Objectifs Méthodes d'allocation Simulation de mémoire virtuelle Le mapping Introduction Plusieurs dizaines de processus doivent se partager
Chapitre V : La gestion de la mémoire Hiérarchie de mémoires Objectifs Méthodes d'allocation Simulation de mémoire virtuelle Le mapping Introduction Plusieurs dizaines de processus doivent se partager
1 les caractères des êtres humains.
 Quelques rappels des classes précédentes ACTIVITÉ livre pages 8 et 9 : apprendre le bilan de la page 9 Les êtres vivants sont répartis en espèces. Chaque être vivant est formé de cellules. schéma d une
Quelques rappels des classes précédentes ACTIVITÉ livre pages 8 et 9 : apprendre le bilan de la page 9 Les êtres vivants sont répartis en espèces. Chaque être vivant est formé de cellules. schéma d une
Conférence technique internationale de la FAO
 Décembre 2009 ABDC-10/7.2 F Conférence technique internationale de la FAO Biotechnologies agricoles dans les pays en développement: choix et perspectives pour les cultures, les forêts, l élevage, les pêches
Décembre 2009 ABDC-10/7.2 F Conférence technique internationale de la FAO Biotechnologies agricoles dans les pays en développement: choix et perspectives pour les cultures, les forêts, l élevage, les pêches
Partie 1. Addition nucléophile suivie d élimination (A N + E) 1.1. Réactivité électrophile des acides carboxyliques et groupes dérivés
 1.1. Réactivité électrophile des acides carboxyliques et groupes dérivés") Molécules et matériaux organiques Partie 1. Addition nucléophile suivie d élimination (A N + E) 1.1. Réactivité électrophile des acides carboxyliques et groupes dérivés bjectifs du chapitre Notions à connaître
Molécules et matériaux organiques Partie 1. Addition nucléophile suivie d élimination (A N + E) 1.1. Réactivité électrophile des acides carboxyliques et groupes dérivés bjectifs du chapitre Notions à connaître
TD DOSAGE DE PROTEINES ET ELECTROPHORESE : PARTIE THÉORIQUE BST1 SVT
 TD DOSAGE DE PROTEINES ET ELECTROPHORESE : PARTIE THÉORIQUE BST1 SVT Daniela LENER IBMC Texte conseillé pour consultation : Biochimie, Voet & Voet, ed. De Boeck. Dosage des protéines Pendant une purification
TD DOSAGE DE PROTEINES ET ELECTROPHORESE : PARTIE THÉORIQUE BST1 SVT Daniela LENER IBMC Texte conseillé pour consultation : Biochimie, Voet & Voet, ed. De Boeck. Dosage des protéines Pendant une purification
Production d une protéine recombinante
 99 Production d une protéine recombinante Lic. B. PIRSON Lic. J-M. SERONT ISICHt - Mons Production de la protéine recombinante GFP (Green Fluorescent Protein d Aequoria victoria) par une bactérie ( E.
99 Production d une protéine recombinante Lic. B. PIRSON Lic. J-M. SERONT ISICHt - Mons Production de la protéine recombinante GFP (Green Fluorescent Protein d Aequoria victoria) par une bactérie ( E.
Parcours de visite, lycée Exposition: LA RADIOACTIVITÉ De Homer à oppenheimer
 Complétez le schéma de gestion des déchets nucléaires en vous aidant du panneau, les surfaces des cercles sont proportionnelles à leur importance Parcours de visite, lycée Exposition: LA RADIOACTIVITÉ
Complétez le schéma de gestion des déchets nucléaires en vous aidant du panneau, les surfaces des cercles sont proportionnelles à leur importance Parcours de visite, lycée Exposition: LA RADIOACTIVITÉ
pka D UN INDICATEUR COLORE
 TP SPETROPHOTOMETRIE Lycée F.BUISSON PTSI pka D UN INDIATEUR OLORE ) Principes de la spectrophotométrie La spectrophotométrie est une technique d analyse qualitative et quantitative, de substances absorbant
TP SPETROPHOTOMETRIE Lycée F.BUISSON PTSI pka D UN INDIATEUR OLORE ) Principes de la spectrophotométrie La spectrophotométrie est une technique d analyse qualitative et quantitative, de substances absorbant
Sérodiagnostic de la polyarthrite rhumatoïde
 1 ETSL Sérodiagnostic de la polyarthrite rhumatoïde TP 1 GABIN-GAUTHIER 13/11/2009 I. LA MALADIE... 2 II. TECHNIQUES QUALITATIVES... 2 1. PRINCIPE... 2 2. MODE OPERATOIRE... 3 2.1. WRST ou Waaler Rose
1 ETSL Sérodiagnostic de la polyarthrite rhumatoïde TP 1 GABIN-GAUTHIER 13/11/2009 I. LA MALADIE... 2 II. TECHNIQUES QUALITATIVES... 2 1. PRINCIPE... 2 2. MODE OPERATOIRE... 3 2.1. WRST ou Waaler Rose
Chapitre 1: Facteurs d'échelle
 Chapitre 1: Facteurs d'échelle Des considérations générales sur la taille des objets ou des êtres vivants et leur influence sur différents paramètres, permettent d'établir simplement quelques lois ou tendances,
Chapitre 1: Facteurs d'échelle Des considérations générales sur la taille des objets ou des êtres vivants et leur influence sur différents paramètres, permettent d'établir simplement quelques lois ou tendances,
Compléments - Chapitre 5 Spectroscopie
 ompléments - hapitre 5 Spectroscopie Spectroscopie par résonance magnétique nucléaire (RMN 13 ) Tandis que la spectroscopie RMN 1 H fournit des données sur la disposition des atomes d'hydrogène dans une
ompléments - hapitre 5 Spectroscopie Spectroscopie par résonance magnétique nucléaire (RMN 13 ) Tandis que la spectroscopie RMN 1 H fournit des données sur la disposition des atomes d'hydrogène dans une